Genotype by Location Effects on Yield and Seed Nutrient Composition of Common Bean


 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Locations and Site Descriptions
2.2. Experimental Design and Site Management
2.3. Data Collection and Analysis
3. Results
3.1. Impact of Location on Yield and Seed Weight
3.2. Impact of Location on Common Bean Seed Elemental Concentrations
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rawal, V.; Navarro, D.K. The Global Economy of Pulses; FAO: Rome, Italy, 2019; Available online: http://www.fao.org/3/i7108en/i7108en.pdf (accessed on 22 January 2020).
- Broughton, W.J.; Hernandez, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—Model food legumes. Plant Soil 2003, 252, 55–128. [Google Scholar] [CrossRef] [Green Version]
- Beebe, S.; Gonzalez, A.V.; Rengifo, J. Research on trace minerals in the common bean. Food Nutr. Bull. 2000, 21, 387–391. [Google Scholar] [CrossRef]
- Mensack, M.M.; Fitzgerald, V.K.; Ryan, E.P.; Lewis, M.R.; Thompson, H.J.; Brick, M.A. Evaluation of diversity among common beans (Phaseolus vulgaris L.) from two centers of domestication using ‘omics’ technologies. BMC Genom. 2010, 11, 686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gül, K.; Egesel, C.Ö.; Turhan, H. The effects of planting time on fatty acids and tocopherols in chickpea. Eur. Food Res. Technol. 2008, 226, 517–522. [Google Scholar] [CrossRef]
- Wood, J.A.; Knights, E.J.; Harden, S. Milling performance in desi-type chickpea (Cicer arietinum L.): Effects of genotype, environment and seed size. J. Sci. Food Agric. 2008, 88, 108–115. [Google Scholar] [CrossRef]
- Graham, R.D.; Welch, R.M.; Bouis, H.E. Addressing micronutrient malnutrition through enhancing the nutritional quality of staple foods: Principles, perspectives and knowledge gaps. Adv. Agron. 2001, 70, 77–142. [Google Scholar] [CrossRef]
- Graham, R.D. Breeding for nutritional characteristics in cereals. Adv. Plant. Nutr. 1984, 1, 57–102. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Marschner, H.; Römheld, V. Strategies of plants for acquisition of iron. Plant Soil 1994, 165, 261–274. [Google Scholar] [CrossRef]
- Obrador, A.; Novillo, J.; Alvarez, J.M. Mobility and availability to plants of two zinc sources applied to a calcareous soil. Soil Sci. Soc. Am. J. 2003, 67, 564–572. [Google Scholar] [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification. Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Sarkar, A.N.; Wyn Jones, R.G. Effect of rhizosphere pH on the availability and uptake of Fe, Mn and Zn. Plant Soil 2008, 66, 361–372. [Google Scholar] [CrossRef]
- Vanlauwe, B.; Bationo, A.; Chianu, J.; Giller, K.E.; Merckx, R.; Mokwunye, U.; Ohiokpehai, O.; Pypers, P.; Tabo, R.; Shepherd, K.D.; et al. Integrated soil fertility management—Operational definition and consequences for implementation and dissemination. Outlook Agric. 2008, 39, 17–24. [Google Scholar] [CrossRef] [Green Version]
- US Department of Agriculture. Agricultural Research Service. Nutrient Data Laboratory. USDA National Nutrient Database for Standard Reference, Release 28 (Slightly revised). Version Current: May 2016. Available online: http://www.ars.usda.gov/nea/bhnrc/mafcl (accessed on 2 July 2019).
- Hofstrand, J. Economics of Tile Drainage. Iowa State Univ., Ext. and Outreach. 2010. Available online: https://www.extension.iastate.edu/agdm/articles/ hof/HofJuly10.html (accessed on 20 May 2019).
- US Climate DATA. Available online: https://www.usclimatedata.com/climate/boone/iowa/united-states/usia0091 (accessed on 5 September 2019).
- USDA. National Resources Conservation Service. Official Soil Series Descriptions. Available online: https://soilseries.sc.egov.usda.gov/osdname.aspx (accessed on 24 September 2019).
- Franzen, D.W. Fertilizing Pinto, Navy and Other Dry Edible Bean. SF720. North Dakota State Univ., Fargo. 2017. Available online: https://www.ag.ndsu.edu/publications/crops/fertilizing-pinto-navy-and-other-dry-edible-bean/sf720.pdf (accessed on 24 September 2019).
- TAXOUSDA. Predicted USDA Soil Taxonomy Class (Twelfth Edn). 2014. Available online: https://soilgrids.org/#!/?lon=31.683333&lat=-0.3&zoom=14&layer=ORCDRC_M_sl2_250m&vector=1&showInfo=1 (accessed on 16 April 2019).
- Mugume, I.; Mesquita, M.D.S.; Basalirwa, C.; Bamutaze, Y.; Reuder, J.; Nimusiima, A.; Waiswa, D.; Mujuni, G.; Tao, S.; Ngailo, T.J. Patterns of Dekadal rainfall variation over a selected region in Lake Victoria basin, Uganda. Atmosphere 2016, 7, 150. [Google Scholar] [CrossRef] [Green Version]
- Lunze, L.; Abang, M.M.; Buruchara, R.; Ugen, M.A.; Nabahungu, N.L.; Rachier, G.O.; Ngongo, M.; Rao, I. Integrated soil fertility management in bean-based cropping systems of Eastern, Central and Southern Africa. In Soil Fertility Improvement and Integrated Nutrient Management—A Global Perspective; Whalen, J.K., Ed.; InTech Open Ltd.: London, UK, 2012; pp. 239–272. [Google Scholar]
- Mallarino, A.; Sawyer, J.E.; Barnhart, S.K. A General Guide for Crop Nutrient and Limestone Recommendations in Iowa. PM1688; Iowa State Univ., Ext. and Outreach: Ames, IA, USA, 2013. [Google Scholar]
- Copeland, L.O.; Hesterman, O.B.; Pierce, F.J.; Tesar, M.B. AG FACTS—Seeding Practices for Michigan Crops; Michigan State Univ. Coop. Ext. Serv: East Lansing, MI, USA, 2016; Available online: http://fieldcrop.msu.edu/uploads/documents/E2107.pdf (accessed on 22 July 2019).
- Sunday, G.; Ocen, D. Fertilizer Consumption and Fertilizer Use by Crop in Uganda; Ministry of Agriculture and, Animal Industry and Fisheries, Uganda National Bureau of Statistics: Kampala, Uganda, 2015.
- De Ron, A.M.; Rodiño, A.P.; Santalla, M.; González, A.M.; Lema, M.J.; Martín, I.; Kigel, J. Seedling emergence and phenotypic response of common bean germplasm to different temperatures under controlled conditions and in open field. Front. Plant Sci. 2016, 7, 1087. [Google Scholar] [CrossRef] [Green Version]
- Kandel, H.; Osorno, J.; VanderWal, J.; Posch, J.; Cooper, K.; Eslinger, H.; Nelson, S.; Ostlie, M.; Schatz, B.; Endres, G.; et al. North Dakota dry bean. Variety Trial Results for 2018 and Selection Guide; A654-18; North Dakota State University Extension: Fargo, ND, USA, 2018. [Google Scholar]
- Balasubramanian, P.M.; Chatterton, S.; Conner, R.L. AAC Y012 and AAC Y015 yellow dry bean. Cultivar description. Can. J. Plant Sci. 2017, 97, 340–343. [Google Scholar]
- Delate, K. Alternative Crops for Iowa. 2013. Available online: https://www.extension.iastate.edu/alternativeag/cropproduction.html (accessed on 10 September 2019).
- Fageria, N.K.; Baligar, V.C.; Moreira, A.; Portes, T.A. Dry bean genotypes evaluation for growth, yield components and phosphorus use efficiency. J. Plant Nutr. 2010, 33, 2167–2181. [Google Scholar] [CrossRef]
- Mourice, S.K.; Tryphone, G.M. Evaluation of common bean (Phaseolus vulgaris L.) genotypes for adaptation to low phosphorus. ISRN Agron. 2012, 2012, 309614. [Google Scholar] [CrossRef] [Green Version]
- Turuko, M.; Mohammed, A. Effect of different phosphorus fertilizer rates on growth, dry matter yield and yield components of common bean (Phaseolus vulgaris L.). World J. Agric. Res. 2014, 2, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Chekanai, V.; Chikowo, R.; Vanlauwe, B. Response of common beans (Phaseolus vulgaris L.) to nitrogen, phosphorus and rhizobia inoculation across variable soils in Zimbabwe. Agric. Ecosyst. Environ. 2018, 266, 167–173. [Google Scholar] [CrossRef]
- Ribeiro, N.D.; da Rosa, S.S.; Jost, E.; Rosa, D.P.; Poersch, N.L.; Maziero, S.M. Genetics of phosphorus content in common bean seeds. Crop Breed Appl. Biotechnol. 2011, 11, 250–256. [Google Scholar] [CrossRef]
- Lima, E.R.; Santiago, A.S.; Araujo, A.P.; Teixeira, M.G. Effects of the size of sown seed on growth and yield of common bean cultivars of different seed sizes. Braz. J. Plant Physiol. 2005, 17, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Leleji, O.I.; Dickson, M.H.; Crowder, L.V.; Bourke, J.B. Inheritance of crude protein percentage and its correlation with seed yield in beans, Phaseolus vulgaris L. Crop Sci. 1972, 12, 168–171. [Google Scholar] [CrossRef]
- Strauta, L.; Muizniece-Brasava, S.; Alsina, I. Crude protein and ash content in different colored Phaseolus coccineus L. World Acad. Sci. Eng. Technol. Internat. J. Nutr. Food Eng. 2013, 7, 696–701. [Google Scholar]
- Bänziger, M.; Long, J. The potential for increasing the iron and zinc density of Maize through plant-breeding. Food Nutr. Bull. 2000, 21, 397–400. [Google Scholar] [CrossRef]
- Hirschi, K.D. Nutrient biofortification of food crops. Annu. Rev. Nutr. 2009, 29, 401–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petry, N.; Boy, E.; Wirth, J.P.; Hurrell, R.F. Review: The potential of the common bean (Phaseolus vulgaris) as a vehicle for iron biofortification. Nutrients 2015, 7, 1144–1173. [Google Scholar] [CrossRef] [Green Version]
- Schachtman, D.P.; Barker, S.J. Molecular approaches for increasing the micronutrient density in edible portions of food crops. Field Crops Res. 1999, 60, 81–92. [Google Scholar] [CrossRef]
- Uauy, C.; Distelfeld, A.; Fahima, T.; Blechl, A.; Dubcovsky, J. A NAC gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science 2006, 314, 1298–1301. [Google Scholar] [CrossRef] [Green Version]
- Rengel, Z. Physiological mechanisms underlying differential nutrient efficiency of crop genotypes. In Mineral Nutrition of Crops—Fundamental Mechanisms and Implications; Rengel, Z., Ed.; Food Products Press: Binghamton, NY, USA, 1999; Volume 1, pp. 227–265. [Google Scholar]
- Aciksoz, S.B.; Yazici, A.; Ozturk, L.; Cakmak, I. Biofortification of wheat with iron through soil and foliar application of nitrogen and iron fertilizers. Plant Soil 2011, 349, 215–225. [Google Scholar] [CrossRef]
- Ramolemana, G.M. Influence of soil chemical characteristics on the nutritional value of morama (Tylosema esculentum) bean seed a potential crop in Botswana. J. Agric. Sci. 2013, 5, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Zou, C.Q.; Zhang, Y.Q.; Rashid, A.; Ram, H.; Sohu, V.; Hassan, M.; Kaya, Y.; Onder, O.; Lungu, O.; Yagu, M.M.; et al. Biofortification of wheat with zinc through zinc fertilization in seven countries. Plant Soil 2012, 361, 119–130. [Google Scholar] [CrossRef]
- Phattarakul, N.; Rerkasem, B.; Li, L.J.; Wu, L.H.; Zou, C.Q.; Ram, H.; Sohu, V.S.; Kang, F.S.; Surek, H.; Kalayci, M.; et al. Biofortification of rice grain with zinc through zinc fertilization in different countries. Plant Soil 2012, 361, 131–141. [Google Scholar] [CrossRef]
- Welch, R.M. Effects of nutrient deficiencies on seed production and quality. Adv. Plant Nutr. 1986, 2, 205–247. [Google Scholar]
- Moraghan, J.T.; Grafton, K. Genetic diversity and mineral composition of common bean seed. J. Sci. Food Agric. 2001, 81, 404–408. [Google Scholar] [CrossRef]
- Alley, M.M.; Vanlauwe, B. The Role of Fertilizers in Integrated Plant Nutrient Management, 1st ed.; IFA: Paris, France; TSBF-CIAT: Nairobi, Kenya, 2009. [Google Scholar]
- Tiessen, H. Phosphorus in the Global Environment. In The Ecophysiology of Plant-Phosphorus Interactions; White, P.J., Hammond, J.P., Eds.; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Fageria, N.K.; da Costa, J.G.C. Evaluation of common bean genotypes for phosphorus use efficiency. J. Plant Nutr. 2000, 23, 1145–1152. [Google Scholar] [CrossRef]
- Polania, J.; Rao, I.M.; Cajiao, C.; Grajales, M.; Rivera, M.; Velasquez, F.; Raatz, B.; Beebe, S.E. Shoot and root traits contribute to drought resistance in recombinant inbred lines of MD 23-24 × SEA 5 of common bean. Front. Plant Sci. 2007, 8, 296. [Google Scholar] [CrossRef]
- Lynch, J.P.; Brown, K.M. Topsoil foraging—An architectural adaptation of plants to low phosphorus availability. Plant Soil 2001, 237, 225–237. [Google Scholar] [CrossRef]
- Shen, J.; Yuan, L.; Zhang, J.; Li, H.; Bai, Z.; Chen, X.; Zhang, W.; Zhang, F. Phosphorus dynamics: From soil to plant. Plant Physiol. 2011, 156, 997–1005. [Google Scholar] [CrossRef] [Green Version]
- Dechassa, N.; Schenk, M.K. Exudation of organic anions by roots of cabbage carrot and potato as influenced by environmental factors and plant age. J. Plant Nutr. Soil Sci. 2004, 167, 623–629. [Google Scholar] [CrossRef]
- Hammond, J.P.; Broadley, M.R.; White, P.J. Genetic responses to phosphorus deficiency. Ann. Bot. 2004, 94, 323–332. [Google Scholar] [CrossRef]
- Islam, F.M.A.; Basford, K.E.; Jara, C.; Redden, R.J.; Beebe, S. Seed compositional and disease resistance differences among gene pools in cultivated common bean. Genet. Res. Crop Evol. 2002, 49, 285–293. [Google Scholar] [CrossRef]
- Sangakkara, U.R.; Hartwig, U.A.; Nosberger, J. Soil moisture and potassium affect the performance of symbiotic nitrogen fixation in faba bean and common bean. Plant Soil 1996, 184, 123–130. [Google Scholar] [CrossRef]
- Laboski, C.A.M.; Peters, J.B. Nutrient Application Guidelines for Field, Vegetable, and Fruit Crops in Wisconsin; A2809; University of Wisconsin Cooperative Extension: Madison, WI, USA, 2012. [Google Scholar]
- Parnes, R. Soil Fertility. A Guide to Organic and Inorganic Soil Amendments. 2013. Available online: https://www.nofa.org/soil/html/potassium.php (accessed on 3 July 2019).
- Fageria, N.K.; Barbosa Filho, M.P.; da Costa, J.G.C. Potassium use efficiency in common bean genotypes. J. Plant Nutr. 2001, 24, 1937–1945. [Google Scholar] [CrossRef]
- Manjula, N. Soils, plant nutrition and nutrient management. In Soil Testing and Plant Diagnostic Service Laboratory; University of Missouri Extension: Columbia, MO, USA, 2017; Available online: https://extension2.missouri.edu/mg4 (accessed on 14 September 2019).
- Cichy, K.A.; Wiesinger, J.A.; Berry, M.; Nchimbi-Msolla, S.; Fourie, D.; Porch, T.G.; Ambechew, D.; Miklas, P.N. The role of genotype and production environment in determining the cooking time of dry beans (Phaseolus vulgaris L.). Legume Sci. 2019, 1, e13. [Google Scholar] [CrossRef] [Green Version]
- Hirpa, L.; Nigussie-Dechassa, R.; Setegn, G.; Bultosa, G. Chemical quality of common beans as influences by genotype and aluminum rates under two soil liming regimes. Afr. J. Food Agric. Nutr. Dev. 2015, 15, 9872–9891. [Google Scholar]
- Sadras, V.O. Evolutionary aspects of the trade-off between seed size and number in crops. Field Crops Res. 2007, 100, 125–138. [Google Scholar] [CrossRef]
- Smith, J.R.; Rao, I.M.; Merchant, A. Source-sink relationships in crop plants and their influence on yield development and nutritional quality. Front. Plant Sci. 2018, 9, 1889. [Google Scholar] [CrossRef] [Green Version]
- Beebe, S. Common Bean Breeding in the Tropics. In Plant Breeding Reviews; Janick, J., Ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2012; Volume 36, pp. 357–426. [Google Scholar]



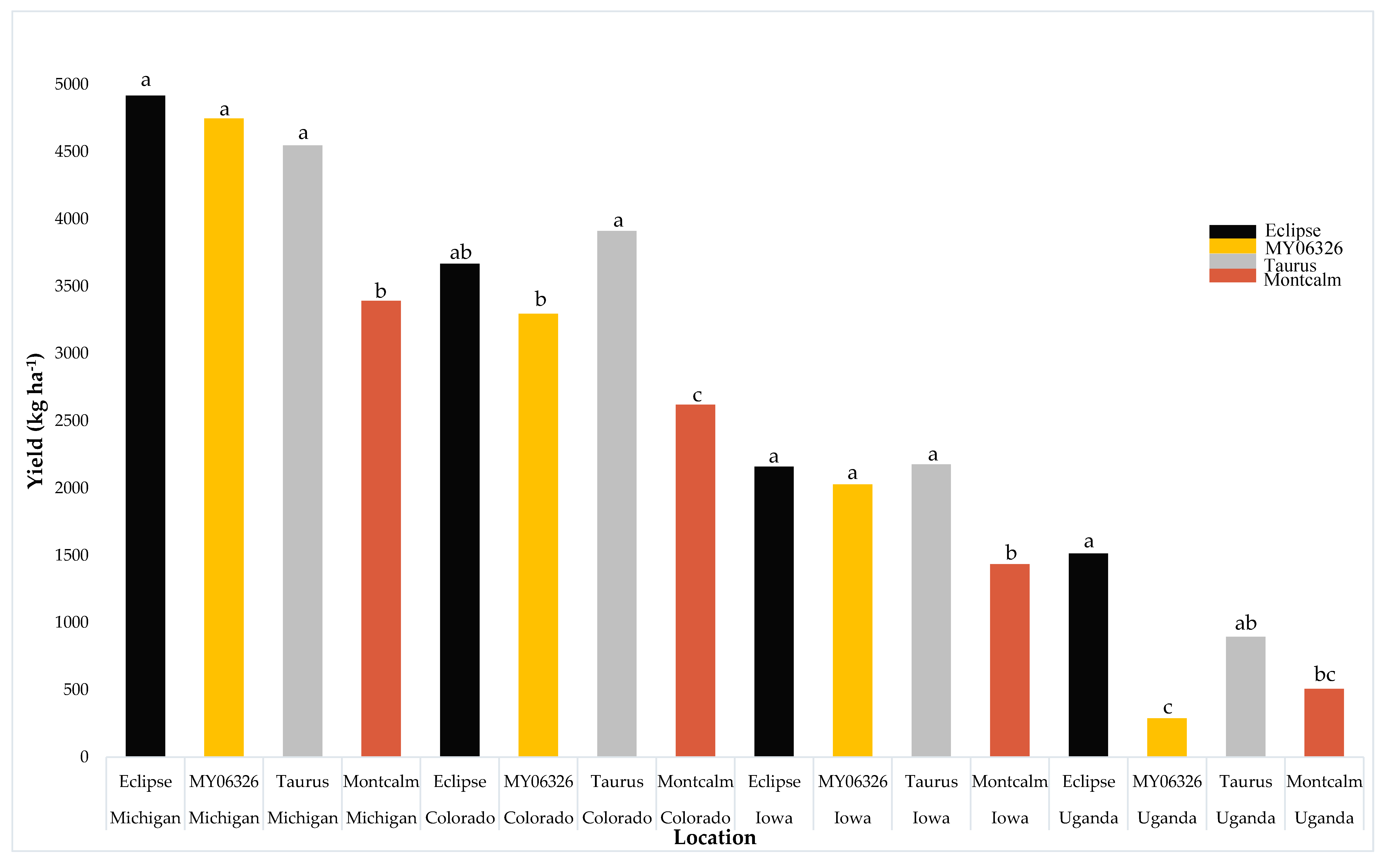{kind=link}
{kind=link}
| Location | Elevation | Annual Precipitation | Previous Crop | Row Spacing | Nutrient Additions | Preplant Tillage | Planting Date | Weed Management | Insect Management |
|---|---|---|---|---|---|---|---|---|---|
| m. m.s.l. | cm | cm | kg ha−1 | ||||||
| Colorado, USA | 1550 | 408 | maize | 76 | 247 46-0-0 6725 manure 80 11-52-0 | vertical tillage, moldboard plow, field cultivator twice | 9 Jun. 2017 30 May 2018 | pendimethalin, eptam, S-metolachlor, cultivation | not required |
| Iowa, USA | 325 | 974 | maize | 76 | 247 18-46-0 195 0-0-62 | chisel plow, tandem disk, field cultivator | 31 May 2017 16 May 2018 | pendimethalin, cultivation, hand weeding | dimethoate |
| Michigan, USA | 287 | 854 | maize | 50 | 91 19-10-19 45 46-0-0 | chisel plot, tandem disk, field cultivator | 13 Jun. 2017 13 Jun. 2018 | eptam, ethylfluralin, S-metolachlor, bentazon, fomesafen, imazamox | esfenvalerate |
| Masaka, Uganda | 1242 | 658 | maize | 50 | 124 17-17-17 14,826 limestone | moldboard plow, hand hoeing to level and fine | 7 Nov. 2017 3 Mar. 2018 | hand weeding three times | cypermethrin |
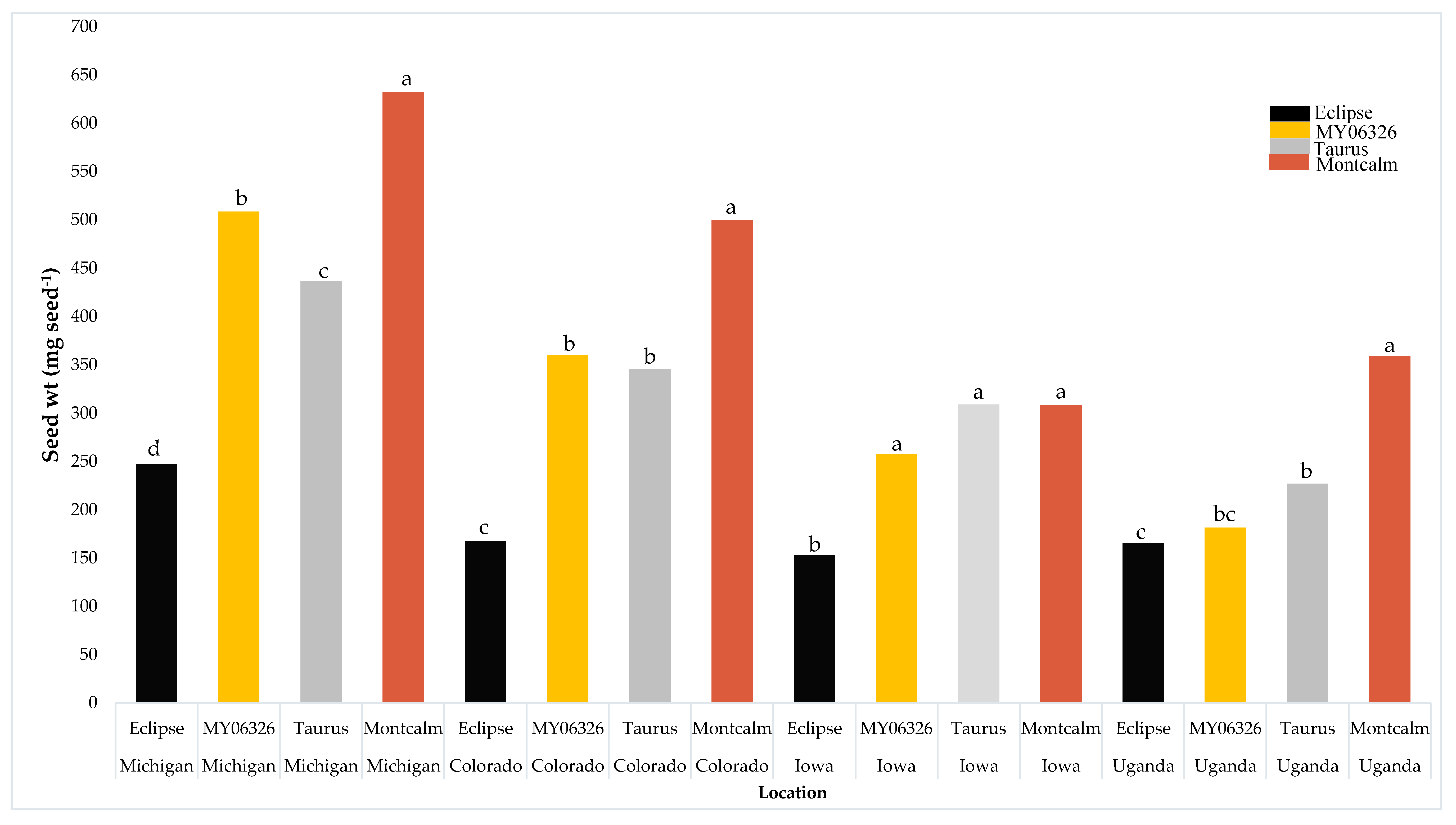
| Parameter | Stand Density R9 (no. m−2) | Yield (kg ha−1) | Seed Weight (mg seed−1) |
|---|---|---|---|
| Location | |||
| Colorado | 23 b | 3374 a | 343 b |
| Iowa | 37 a | 1951 b | 257 c |
| Michigan | 21 b | 4402 a | 456 a |
| Uganda | 17 b | 802 b | 233 c |
| SE | 2.3 | 677 | 33 |
| Variety | |||
| Eclipse | 27 a | 3066 a | 183 c |
| Montcalm | 18 b | 1989 c | 450 a |
| MY06326 | 25 a | 2590 b | 327 b |
| Taurus | 27 a | 2883 ab | 330 b |
| SE | 1.5 | 386 | 18 |
| p > F | |||
| Location (L) | *** | *** | *** |
| Variety (V) | *** | *** | *** |
| L × V | ns | * | *** |
| Location | pH | OM (%) | Fe (mg kg−1) | Zn (mg kg−1) | P (mg kg−1) | K (mg kg−1) | Na (mg kg−1) | Mg (mg kg−1) | Mn (mg kg−1) | N (mg kg−1) | Al (mg kg−1) | SO4−2-S (mg kg−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Colorado | 8.0 a | 3.0 | 34.6 | 6.6 | 54.4 b | 324.5 a | 47.6 a | 602.0 a | 61.8 ab | 1100 b | 47.0 c | 30.5 a |
| Iowa | 6.2 b | 3.0 | 76.0 | 3.0 | 15.7 b | 148.9 bc | 25.6 b | 308.3 b | 23.5 b | 1500 a | 827.9 b | 4.4 c |
| Michigan | 6.0 b | 1.7 | 145.6 | 5.0 | 214 a | 190.9 b | 15.2 b | 125.8 c | 53.0 b | 1000 b | 949.5 a | 12.0 bc |
| Uganda | 5.9 b | 2.3 | 123.6 | 3.2 | 24.8 b | 97.6 c | 20.0 b | 179.0 c | 129.0 a | 1500 a | N/A | 15.8 b |
| SE | 0.3 | 1.0 | 69.7 | 3.5 | 27.2 | 39.8 | 6.2 | 34.5 | 38.6 | 188 | 18.4 | 4.8 |
| p > F | *** | ns | ns | ns | *** | *** | *** | *** | ** | *** | *** | *** |
| Parameter | CP (g kg−1) | Fe (mg kg−1) | Zn (mg kg−1) | P (mg kg−1) | K (mg kg−1) | Mg (mg kg−1) | Mn (mg kg−1) | Al (mg kg−1) | Na (mg kg−1) | S (mg kg−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| Location | ||||||||||
| Colorado | 21 | 150 | 45 | 4253 | 13696 | 1871 | 23 | 147 | 61 | 2405 |
| Iowa | 22 | 111 | 55 | 5395 | 13837 | 2083 | 33 | 25 | 217 | 2221 |
| Michigan | 20 | 61 | 38 | 4901 | 14143 | 1773 | 19 | 174 | 56 | 1921 |
| Uganda | 27 | 65 | 30 | 4395 | 12998 | 1902 | 21 | NA | 12 | 2342 |
| Variety | ||||||||||
| Eclipse | 21 b | 141 | 40 | 4181 b | 12648 b | 2016 ab | 26 | 196 | 86 | 2181 ab |
| Montcalm | 24 a | 83 | 52 | 5110 a | 13432 ab | 1658 c | 22 | 107 | 86 | 2341 a |
| MY06326 | 22 ab | 79 | 38 | 4750 ab | 14473 a | 1846 bc | 24 | 82 | 93 | 2028 b |
| Taurus | 23 ab | 84 | 39 | 4899 ab | 14125 a | 2108 a | 23 | 75 | 80 | 2338 a |
| SE | 1.0 | 36.1 | 10.4 | 482 | 933 | 27828 | 7.2 | 84 | 4771 | 105 |
| p > F | ||||||||||
| Location (L) | 0.08 | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Variety (V) | * | ns | ns | * | ** | *** | ns | ns | ns | ** |
| L × V | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| P | K | Mg | Na | Zn | Al | Fe | S | Mn | CP 1 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Seed weight r | 0.136 | 0.133 | −0.394 | −0.027 | 0.098 | −0.166 | −0.112 | −0.131 | −0.058 | −0.223 |
| p > r | 0.19 | 0.20 | 0.0001 | 0.79 | 0.35 | 0.17 | 0.29 | 0.21 | 0.61 | 0.03 |
| Yield r | 0.008 | 0.200 | −0.020 | 0.009 | 0.156 | 0.089 | 0.160 | −0.160 | 0.123 | −0.750 |
| p > r | 0.94 | 0.06 | 0.85 | 0.93 | 0.14 | 0.46 | 0.13 | 0.13 | 0.27 | 0.0001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bulyaba, R.; Winham, D.M.; Lenssen, A.W.; Moore, K.J.; Kelly, J.D.; Brick, M.A.; Wright, E.M.; Ogg, J.B. Genotype by Location Effects on Yield and Seed Nutrient Composition of Common Bean. Agronomy 2020, 10, 347. https://doi.org/10.3390/agronomy10030347
Bulyaba R, Winham DM, Lenssen AW, Moore KJ, Kelly JD, Brick MA, Wright EM, Ogg JB. Genotype by Location Effects on Yield and Seed Nutrient Composition of Common Bean. Agronomy. 2020; 10(3):347. https://doi.org/10.3390/agronomy10030347
Chicago/Turabian StyleBulyaba, Rosemary, Donna M. Winham, Andrew W. Lenssen, Kenneth J. Moore, James D. Kelly, Mark A. Brick, Evan M. Wright, and James B. Ogg. 2020. "Genotype by Location Effects on Yield and Seed Nutrient Composition of Common Bean" Agronomy 10, no. 3: 347. https://doi.org/10.3390/agronomy10030347
APA StyleBulyaba, R., Winham, D. M., Lenssen, A. W., Moore, K. J., Kelly, J. D., Brick, M. A., Wright, E. M., & Ogg, J. B. (2020). Genotype by Location Effects on Yield and Seed Nutrient Composition of Common Bean. Agronomy, 10(3), 347. https://doi.org/10.3390/agronomy10030347