Hereafter, results coming from the BIOFOSF field case-studies are reported and discussed with respect to the relevant hypotheses.
3.1. Case-Study on Potato Crop Cultivation
In
Table 3, the PHY and ET-PHY contents in all FERT and PPP used in the potato case-study are reported.
As expected, FOS contained both the PHY (1800 mg kg−1) and ET-PHY (441 mg kg−1), while K-PHY only PHY (555,000 g kg−1). An amount of 1.8 mg kg−1 of PHY was found in pelleted poultry manure, in absence of contemporary presence of ET-PHY.
The analysis of all collected soil samples gave an ET-PHY content always lower than 0.01 mg kg
−1 (<LOQ
ET-PHY) and PHY content always lower than 0.1 mg kg
−1 (<LOQ
PHY), without any significant difference among treatments, both at the beginning of the trial and at harvesting (data not reported). In
Table 4, PHY and ET-PHY content in potato leaves, as affected by FERT and PPP, are reported.
The PHY content in potato leaves was significantly affected by FERT only at 105 DAP (
p = 0.0361), being the highest under PM fertilization, while at harvest no FERT effect was observed on PHY and ET-PHY content. On the contrary, as expected, the FERT did not affect the foliar ET-PHY, either at 105 or 150 DAP. The PPP strongly influenced the foliar PHY at both 105 and 150 DAP, being the highest under K-PHY treatment (
p = 0.0008). The ET-PHY was influenced only at 105 DAP (
p = 0.0425), being the highest under FOS treatment, as expected. A slight interaction was observed between FERT and PPP on foliar PHY after 105 DAP (
p = 0.0177), due to the PM fertilization: anyway, the found PHY increase in potato leaves (15.37 mg kg
−1) three months after the animal manure application was not observed at harvest. In
Table 5, PHY and ET-PHY contents in potato tubers, as affected by FERT and PPP, are reported.
In tubers, the fertilization influenced only the PHY content after 105 DAP (P = 0.0290), being again the highest one under PM treatment. The crop protection management strongly affected PHY content in tubers at 105 (p = 0.0006) and 150 DAP (p = 0.0009), but not the ET-PHY one. Again, a significant interaction between FERT and PPP on PHY content in tubers was observed at 105 DAP (p = 0.0442), being the highest under PM treatment.
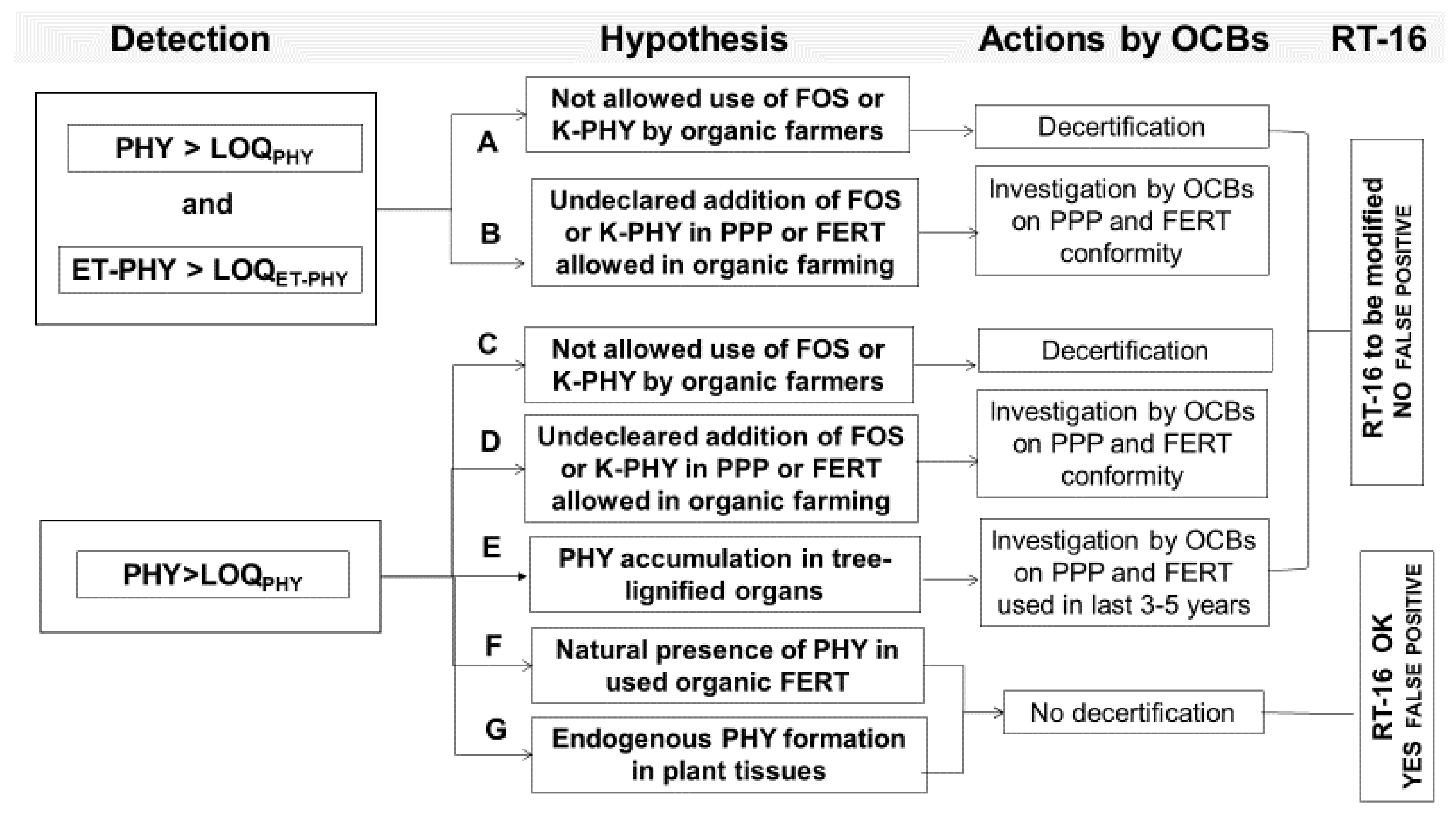
Results obtained on potato confirmed that, after application of FOS or K-PHY, both the ET-PHY and/or PHY had been always detected in potato leaves and/or tubers, but to a different extent from application. When FOS was added by foliar application, ET-PHY residuals were found in young potato leaves only, being fully converted into PHY during the following 45 days. The absence of ET-PHY residual in young tubers after 105 DAP testifies that the ET-PHY acropetal translocation from leaves to tubers is not relevant in potato plant, while it was observed for PHY in our experiment. Due to the very fast degradation process which occurs on leaves after the foliar spraying of the fosetyl, the detection of PHY residues, in absence of contemporary presence of ET-PHY ones, cannot be considered as a “false positive”: thus, it is not possible to exclude the fraudulent application of not allowed PPP and, indeed, the organic product should be decertified (hypothesis A).
When K-PHY was given to the soil, no PHY was detected in soil at 105 and 150 DAP, this confirmed that the PHY conversion into phosphate promptly occurred in considered soil, probably due to the presence of favorable soil conditions, temperature, and presence of phosphite-metabolizing microbes which promoted PHY biological oxidation process [
12]. In fact, thanks to soil microbial activity, PHY oxidation to phosphate in soils lasts approximately 3–4 months, a period which corresponded to the potato growing cycle [
15].
Under soil K-PHY treatment, the PHY content significantly increased in young and old potato leaves, evidencing that the PHY was able to basipetally translocate from root system to leaves The interaction observed between PPP and FERT in young leaves and tubers, as a slight increase of PHY in young potato leaves and tubers under PM fertilization and K-PHY supply, not anymore recorded at harvest, suggests that the basipetal translocation of PHY from soil to the different plant organs took place mainly during the first three months of potato grow in field, when the root system is able to uptake more efficiently. Again, when PHY residues are found into tubers, due to the application of not allowed PPP in organic farming (hypothesis C), the organic product should be decertified.
Since no FOS or PHY contamination was found in potato leaves or tubers under Cu
2O treatments, independently from the applied FERT, we can affirm that no drift effect of FOS/PHY contamination was observed in field. In potato fertilized with PM and using Cu formulate as PPP (hypothesis F), the absence of PHY residues in leaves and tubers either after 105 or after 150 DAP demonstrates that the amount of 1.8 mg/kg of PHY found in pelleted poultry manure did not cause the tuber contamination and thus the organic products should not be decertified. Actually, some organic fertilizers from animal origin characterized by high amount of phosphate, such as the poultry manure, can contain minimum amounts of PHY: in fact, even if phosphonate derivates (such as aminoethyl phosphonates and phosphonolipids) are not usually supplemented in animal diets, PHY is sometimes detected in feces and in manures, suggesting a microbial origin during the stabilization phase [
16]. Anyway, the PHY applied with poultry manure did not cause any residual effect on the crop, likely for the prompt oxidation of PHY into phosphate in the soil [
2].
All the other FERT treatments under Cu2O protection did not give any effect in terms of PHY contamination of leaves and in tubers during the potato grows, confirming that the plant is not able to synthesize PHY by itself in any plant organs (hypothesis G not confirmed). However, the field trial on tomato, not discussed here, again evidenced that, when FERT and PPP used by the farmer were not contaminated by PHY, the same contaminant was not detected in soil, leaves, and tomato barriers (data not reported).
3.2. Case-Study of Fosetyl-Al Degradation in Rocket Lettuce Cultivation
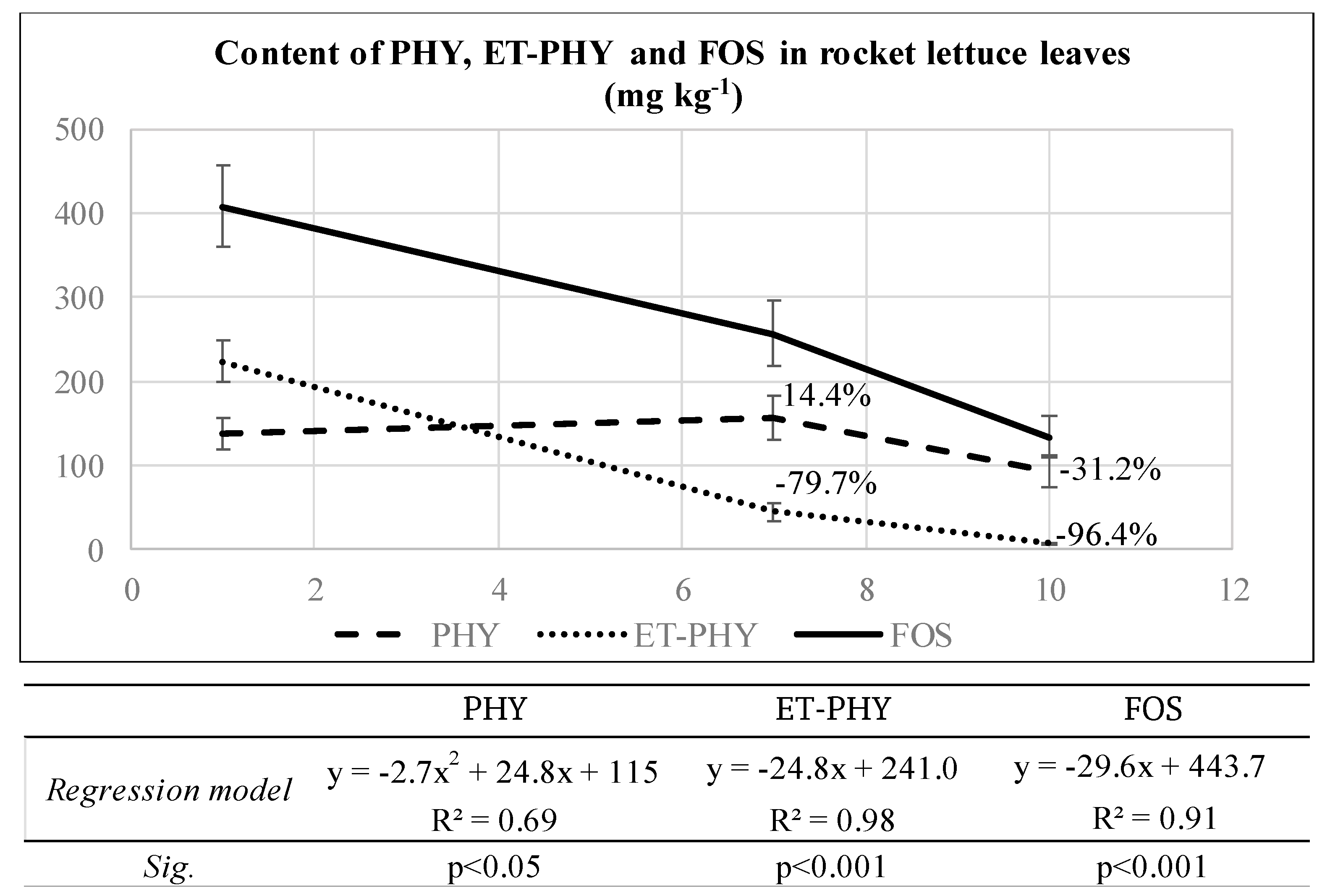
Foliar contents of PHY, ET-PHY, and FOS, calculated as the sum of foliar ET-PHY and PHY in rocket lettuce, percentages of ET-PHY and PHY degradation/ accumulation and curves obtained by regression models for fitting PHY and ET-PHY data are reported in
Figure 2.
A fast degradation of FOS was recorded in rocket lettuce, being 79.7% the foliar degradation of ET-PHY after 7 and 96.4% after 10 days from FOS treatment. On the opposite, at 7 days from FOS treatment, PHY increased to 14.4% in respect to the starting value, while after 10 days it decreased by 31.2%. The best regression model to fit data of foliar ET-PHY was a typical linear regression, where the Y-function was the ET-PHY content, and the X-variable was the time (as days after treatment), giving a negative slope (−24.8) and an intercept with the X-axis at 10 days after treatment. This curve showed a good data fit, being R2 very high (0.98, at p < 0.001). Otherwise, the best regression model able to describe the foliar PHY content was a negative quadratic one, where the Y-function was the PHY content and the X-variable was the time, with a first quadratic term negative (−2.7) and the second one positive (24.8). This second order, polynomial curve did not fit at the best the experimental data, even if the R2 was acceptable (0.69). Regression model for foliar FOS was not considered, since FOS content in rocket lettuce leaves was derived by an applied formula, and thus not an independent variable.
After a foliar treatment with fosetyl-Al of this broadleaf species, the ET-PHY degradation was extremely rapid within the first 10 days after phytosanitary treatment, while again the PHY showed a high persistence on foliar apparatus. The degradation kinetics of ET-PHY and PHY showed opposite trends being, as expected, typically linear the first one, and more responsive to a negative quadratic pattern the second one. This suggested that the PHY kinetic curve is the resultant of two opposite, contemporary processes: from one side, the increase of PHY deriving from ET-PHY degradation and, in parallel, the simultaneous decrease of PHY, probably due to its partial oxidation into phosphate during time.
Ruthbaum and Baille [
38] observed that PHY is highly soluble in water and accessible to plants [
13]. Based on this high solubility, PHY supplied by foliar spray is rapidly absorbed and translocated within the plant [
39,
40]. The following conversion of PHY to phosphate may result from slow chemical oxidation or by oxidizing bacteria and fungi that have been found living, for example, on citrus and avocado leaves [
10,
11,
12,
13,
14,
15,
16,
17,
18,
19,
20,
21,
22,
23,
24,
25,
26,
27,
28,
29,
30,
31,
32,
33,
34,
35,
36,
37,
38,
39,
40,
41]: as a matter of fact, there is consistent evidence that PHY is more readily absorbed into plant tissues than phosphate [
12]. Anyway, phosphonate oxidation to phosphate should provide a good electron source for microbial energy metabolism, as the electrons are released at a very low redox potential [
7]. Some strictly anaerobic, phosphite-oxidizing sulphate-reducing bacteria were found in nature, able to oxidize phosphonate into phosphate [
42]. In plant cells, the only way to oxidize the PHY into phosphate to use PHY as a P-source is through the bacterial phosphite dehydrogenase expression, as already found in Arabidopsis and tobacco [
43]: this mechanism could be claimed to explain our finding on PHY decrease on rocket leaves.
Again, data confirmed that the PHY detection, without contemporary presence of ET-PHY (hypothesis C), has not to be interpreted as a “false positive”, since just after three weeks from foliar application of a fosetyl-derivate, the ET-PHY molecule is not detectable anymore (<0.01 mg kg−1 after 20 days).
3.3. Long-Term Monitoring of Fruit Tree Contamination
In
Table 6, the PHY content in fruit, leaves and in 1-year and 3-years old branches collected from organic pear trees of the two considered varieties (‘Abate Fetel’ and ‘William’) in both the Italian farms (FRIULI-VG. and EMILIA-R.) are reported.
Although farmers used only FERT and PPP allowed in organic farming in the last five years, PHY contamination was detected in fruits of both the two pear varieties and in both the farms. The ‘Abate Fetel’ variety showed higher PHY contamination when compared to ‘William’ ones. Based on fruit PHY content from 2015 to 2018, a significant decrease of PHY contamination was observed on time, except for ‘William’ pears from FRIULI-VG. farm, where in 2016 a significant increase of PHY fruit content (1.4 mg kg
−1) was found compared to the previous year (0.5 mg kg
−1). In 2016 and 2017, a PHY contamination was recorded in leaves, while in 2017 also in young and old pear tree branches, evidencing the ability of pear trees of stocking the PHY for several years in old, lignified plant organs. Regression analysis of PHY content in pears recorded from 2015 to 2018 showed that the exponential equations were the best models able to fit the PHY content decrease in fruits, especially in ER orchards, independent from the tree varieties (
Table 7).
The analysis of the used micronutrient fertilizers, seaweed extracts, Cu-oxichloride, and Bordeaux mixture, applied in the three years (2015, 2016, and 2017) of trials on pear trees by the farmers gave unexpected results: even if all were regularly registered as allowed in organic farming (Annex I and II to the Reg. EC n.889/2008) [
21], five of them contained relevant amount of PHY and, sometimes also of ET-PHY (
Table 8).
Based on previous literature on PHY persistency in tree crops [
14,
41,
42,
43,
44], we carried out this study on pear trees to understand the origin of PHY contamination on organic fruits: we found a PHY contamination in fruits, leaves, but unexpectedly also in young and old branches of pear trees, independently from the orchard location and the pear tree variety. This evidenced a contamination apparently not justifiable by the organic management applied in ER and FR farms, both converted into organic ones from more than ten years. The evaluation of the PPP and FERT applied to pear trees in ER and FR orchards from 2015 to 2017 by the farmers confirmed that all the technical means were allowed in organic farming (Annexes I and II to the Reg. EC n.889/2008) [
21], but three FERT contained PHY between 7.0 and 37 mg kg
−1, and two Cu-based PPP contained relevant amount of both PHY and ET-PHY (up to 656 mg kg
−1). Considering that at least three treatments/year with Bordeaux mixture were carried out on pear trees every year, reasonably the PPP and FERT treatments were the cause of the PHY residues detected in organic pears (hypothesis D). Previous studies on PHY contamination due to foliar FOS application on apples showed that residual PHY level in fruits was positively correlated with the amount of PHY present in the applied FOS or K-PHY formulates, the plant size and yield [
45]. The same authors observed that PHY residuals were accumulated in different plant organs, such as the roots and buds of the apple tree. Our study on pear trees showed that the young, and even the old branches, acted as a physiological sink of PHY, from which during the growing season it translocated to leaves and then fruits for reproductive proposals (hypothesis E). The PHY storage in pear tree branches, and the following PHY decreasing observed in pear fruits during the four years of monitoring suggested that, when fruit trees are contaminated, three years of organic conversion period could be not always enough to guarantee the respect of MLR for pesticide residues, as established by the Italian DM n. 309/2011 [
27]. In fact, it can be stored and re-mobilized by the plant, thus prolonging in the following seasons the residual effect of the FOS/PHY treatments and generating a long-term contamination of fruits which could request more than 5 years to repristinate an uncontaminated system [
45].
Starting from trendlines describing the PHY content in pears, we calculated the corresponding theoretical periods (in days or in years) needed to reach a fruit PHY content equal to quantification limit, that means: y = 0.1 mg kg
−1 as LOQ
PHY (QuPPe Method 1.3 ver.8.1); y = 0.05 mg kg
−1 as LOQ
PHY (QuPPe Method 1.3, ver.10,
Table 7) [
29]. In studied orchards, the tree decontamination could need from 2 to 5 years to reach PHY < 0.1 mg kg
−1 and from 3 to 5 years to reach the PHY < 0.05 mg kg
−1 in fruits, depending on the initial value of the PHY detected in the fruits and lignified organs of trees, but also by the different varieties. This is key for defining MRL of PHY after 3 years of conversion period in organic orchards, especially if they were treated with FOS or K-PHY under conventional management, or in case of accidental contamination by external inputs. With few exceptions, at least five years of conversion period seem an adequate period to assure the complete PHY decontamination of a previously contaminated organic orchard. Anyway, it is important to carry out a preliminary monitoring at the beginning of the organic conversion by using young branches as testing materials for early evaluation of PHY residue level in such orchards. This monitoring would allow to take as soon as possible appropriate measures to reduce undesired contamination by the farmers, for example by selecting FERT and PPP which can guarantee the absence of PHY in their formulation.
3.4. Case-Study on Tomato Greenhouse Cultivation
The analysis of the FERT and PPP used in the greenhouse trial on tomato showed that PM, PK, Cu-Zn fertilizers, and NO did not contain either FOS (<0.01 mg kg−1) or PHY (<0.1 mg kg−1). Unexpectedly, 1.1 mg kg−1 of PHY was found in SM, while FOS was absent (<0.1 mg kg−1).
Soils sampled in the two greenhouses at the beginning and at the end of the trial gave no contamination by FOS (<0.1 mg kg−1) and PHY (<1.0 mg kg−1), independent from the soil fertilization. Similarly, all tomato leaves and berries sampled did not show any FOS (<0.01 mg kg−1) and PHY (<0.1 mg kg−1) residues in both the compared unfertilized/fertilized systems.
As already observed on potato, the absence of PHY also in the CNT tomatoes confirmed again that vegetable plants were not able to produce PHY by ourselves (hypothesis G). It should be remarked the SM, positive to PHY, even though applied several times on tomato by fertigation, did not determine any residue neither in tomato leaves nor in harvested berries. This could be due to low PHY content found in commercial products: in fact, considering 4800 tomato plants in each greenhouse, the five application of SM corresponded to 750 kg ha−1 and to 0.15 g plant−1 of SM. Since SM contained 1.1 mg kg−1 of PHY, each tomato plant received approximately 0.17 mg of PHY during the growing cycle. This PHY supplementation by fertigation was evidently not able to induce contamination in the marketable product: this is in line with the previous results obtained in potato trial, where PM, containing PHY, was applied. These findings evidenced that the application of organic fertilizers cannot be considered the cause of PHY residues detection in organic vegetables, making null the F hypothesis, at least when the recorded PHY content in organic fertilizers is lower than 2 mg kg−1.

 ,
, 



{kind=link}
{kind=link}
{kind=link}