Impacts of Smooth Pigweed (Amaranthus hybridus) on Cover Crops in Southern Ontario


Abstract
:1. Introduction
2. Materials and Methods
2.1. Germination Experiment
2.2. Plant Growth Experiments
2.2.1. Tea
2.2.2. Dried ground
2.2.3. Fresh Trimmed
2.3. Statistical Analysis
3. Results
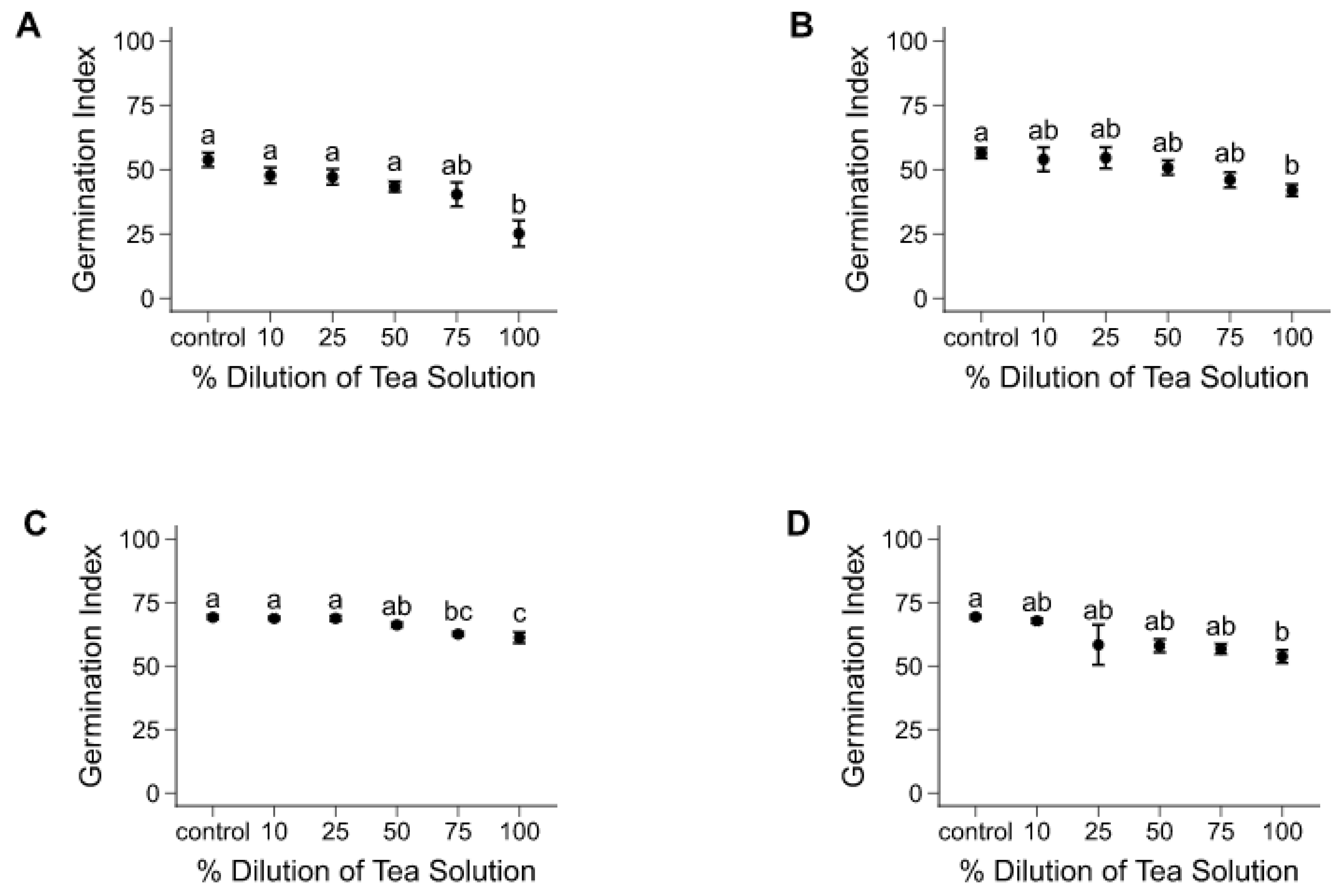
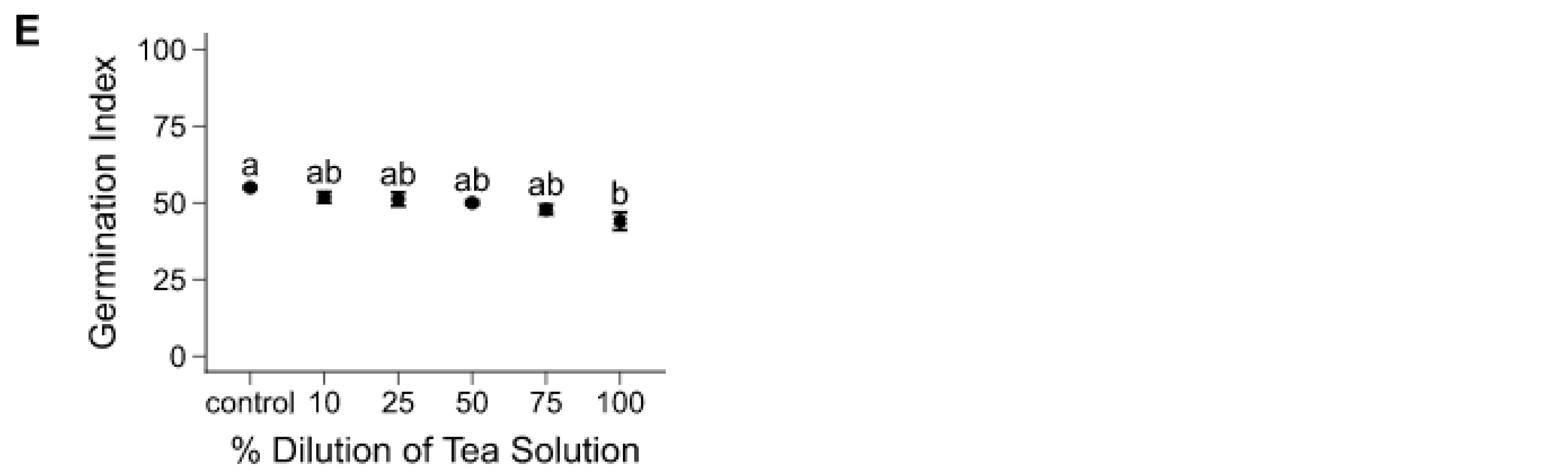
3.1. Seed Germination Experiment
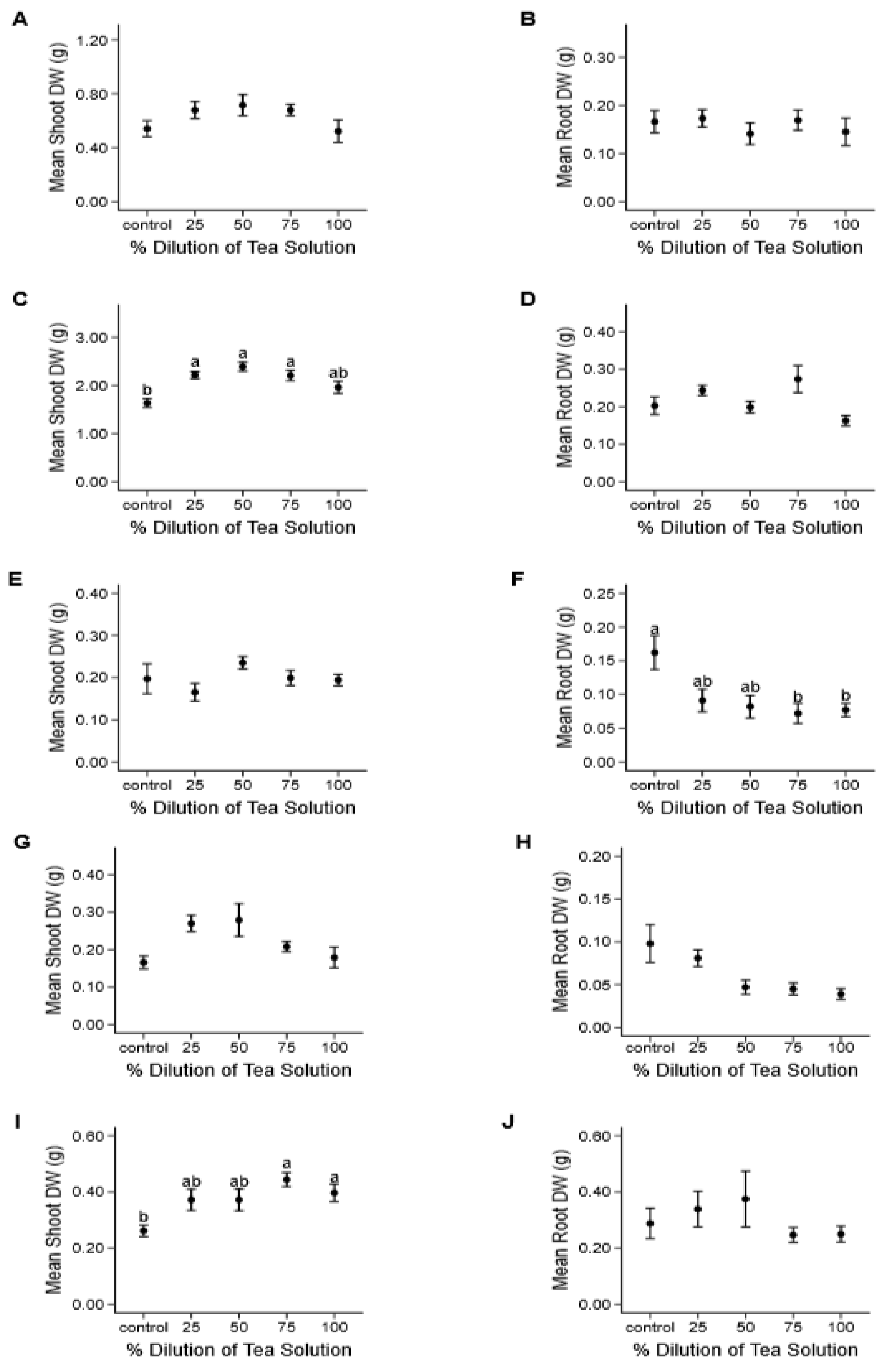
3.2. Growth Experiments
3.2.1. Tea
3.2.2. Dried Ground Material
3.2.3. Fresh Material
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tilman, D. Plant Strategies and the Dynamics and Structure of Plant Communities; Princeton University Press: Princeton, NJ, USA, 1988; pp. 52–97. [Google Scholar]
- Bedoussac, L.; Journet, E.; Hauggaard-Nielsen, H.; Naudin, C.; Corre-Hellou, G.; Jensen, E.S.; Prieur, L.; Justes, E. Ecological principles underlying the increase of productivity achieved by cereal-grain legume intercrops in organic farming. A review. Agron. Sustain. Dev. 2015, 35, 911–935. [Google Scholar] [CrossRef]
- Gitari, H.I.; Karanja, N.N.; Gachene, C.K.K.; Kamau, S.; Sharma, K.; Schulte-Geldermann, E. Nitrogen and phosphorous uptake by potato (Solanum tuberosum L.) and their use efficiency under potato-legume intercropping systems. Field Crops Res. 2018, 222, 78–84. [Google Scholar] [CrossRef]
- Meng, L.; Zhang, A.; Wang, F.; Han, X.; Wang, D.; Li, S. Arbuscular mycorrhizal fungi and Rhizobium facilitate nitrogen uptake and transfer in soybean/maize intercropping system. Front. Plant Sci. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razze, J.M.; Liburd, O.E.; Webb, S.E. Intercropping buckwheat with squash to reduce insect pests and disease incidence and increase yield. Agroecol. Sustain. Food 2016, 40, 863–891. [Google Scholar] [CrossRef]
- Adler, M.J.; Chase, C.A. Comparison of the allelopathic potential of leguminous summer cover crops: Cowpea, sunn hemp, and velvet bean. J. Hortic. Sci. Biotech. 2007, 42, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Gfeller, A.; Herrera, J.M.; Tschuy, F.; Wirth, J. Explanations for Amaranthus retroflexus growth suppression by cover crops. J. Crop Prot. 2018, 104, 11–20. [Google Scholar] [CrossRef]
- Florence, A.M.; Higley, L.G.; Drijber, R.A.; Francis, C.A.; Lindquist, J.L. Cover crop mixture diversity, biomass productivity, weed suppression, and stability. PLoS ONE 2019, 14, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kaye, J.P.; Quemada, M. Using cover crops to mitigate and adapt to climate change. A review. Agron. Sustain. Dev. 2017, 37, 1–17. [Google Scholar] [CrossRef]
- Duke, S.O. Proving allelopathy in crop-weed interactions. Weed Sci. 2015, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Rice, E. Allelopathy, 3rd ed.; Academic Press: Cambridge, MA, USA, 1984; pp. 1–4. [Google Scholar]
- Skinner, E.M.; Diaz-Perez, J.C.; Schomber, H.H.; Vencill, W. Allelopathic effects of sunnhemp (Crotalaria juncea L.) on germination of vegetables and weeds. Hort. Sci. 2012, 47, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Kalinova, S.; Golubinova, I.; Hristoskov, A.; Ilieva, A. Allelopathic effect of aqueous extract from root systems of Johnson grass on seed germination and initial development of soybean, pea and vetch. Ratar. Povrt. 2012, 49, 250–256. [Google Scholar] [CrossRef]
- Carvalho, M.S.S.; Andrade-Vieira, L.F.; Santos, F.E.; Correa, F.F.; Cardoso, M.G.; Vilela, L.R. Allelopathic potential and phytochemical screening of ethanolic extracts from five species of Amaranthus spp. in the plant model Lactuca sativa. Sci. Hortic. 2019, 245, 90–98. [Google Scholar] [CrossRef]
- Wazir, L.; Sadiq, M.; Baloch, M.; Awan, I.; Khan, E.; Shah, I. Application of bio-herbicide alternatives for chemical weed control in rice. Pak. J. Weed Sci. Res. 2011, 17, 245–252. [Google Scholar]
- Mushtaq, M.; Cheema, Z.; Khaliq, A. Effects of mixture of allelopathic plant aqueous extracts on Trianthema portulacastrum L. weed. Allelopath. J. 2010, 25, 205–212. [Google Scholar]
- Wang, M.; Wu, C.; Cheng, Z.; Meng, H. Growth and physiological changes in continuously cropped eggplant (Solanum melongena L.) upon relay intercropping with garlic (Allium sativum L.). Front. Plant Sci. 2015, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Riaz, T.; Khan, S.N.; Javaid, A. Management of Fusarium corm rot of gladiolus (Gladiolus grandiflorus sect. Blandus cv. Aarti) by using leaves of allelopathic plants. Afr. J. Biotechnol. 2010, 9, 4681–4686. [Google Scholar]
- Kruse, M.; Strandberg, M.; Strandberg, B. Ecological Effects of Allelopathic Plants-a Review; NERI Technical Report No. 315; National Environmental Research Institute: Silkeborg, Denmark, 2000; p. 66. [Google Scholar]
- Yanyan, G.; Wang, Q.; Wang, L.; Liu, W.; Liu, X.; Huang, Y.; Christie, P. Response of soil enzymes and microbial communities to root extracts of the alien Alternanthera philoxeroides. Arch. Agron. Soil Sci. 2018, 64, 708–717. [Google Scholar] [CrossRef]
- Ontario Ministry of Agriculture, Food, and Rural Affairs. Pigweeds (Redroot, Green and Smooth); Factsheet Order#:01-009; Queen’s Printer for Ontario: Toronto, ON, Canada, 2009.
- Costea, M.; Weaver, S.E.; Tardif, F.J. The biology of Canadian weeds. 130. Amaranthus retroflexus L., A. powellii S. Watson and A. hybridus L. Can. J. Plant Sci. 2004, 84, 631–668. [Google Scholar] [CrossRef]
- Bakhshayeshan-Agdam, H.; Salehi-Lisar, S.Y.; Motafakkerazad, R.; Talebpour, A.; Farsad, N. Allelopathic effects of redroot pigweed (Amaranthus retroflexus L.) on germination & growth of cucumber, alfalfa, common bean and bread wheat. Acta Agric. Slov. 2015, 105, 193–202. [Google Scholar] [CrossRef]
- Baličević, R.; Ravlić, M.; Čuk, P.; Šević, N. Allelopathic effect of three weed species on germination and growth of onion cultivars. In Proceedings of the 8th International Scientific/Professional Conference Agriculture in Nature and Environment Protection, Vukovar, Croatia, 1–3 June 2015. [Google Scholar]
- Shahrokhi, S.; Hejazi, S.N.; Khodabandeh, H.; Farboodi, M.; Faramarzi, A. Allelopathic effect of aqueous extracts of pigweed, Amaranthus retroflexus L. organs on germination and growth of five barley cultivars. In Proceedings of the 3rd International Conference on Chemical, Biological and Environmental Engineering, Chengdu, China, 23 September 2011; IACSIT Press: Singapore, 2011; Volume 20, pp. 80–84. [Google Scholar]
- De Souza, M.C.; de Carvalho, L.B.; da Costa Aguiar Alves, P.L.; Fidelis Giancotti, P.R. Allelopathy in pigweed (a review). Can. J. Plant Sci. 2011, 1, 5–12. [Google Scholar]
- Trucco, F.; Hager, A.; Tranel, P.J. Acetolactate synthase mutation conferring imidazolinone-specific herbicide resistance in Amaranthus hybridus. J. Plant Physiol. 2006, 163, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, A.I.; Pueyo, Y.; Giner, M.L.; Foronda, A.; Sanchez-Navarrete, P.; Saiz, H.; Alado, C.L. Evidence for chemical interference effect of an allelopathic plant on neighboring plant species: a field study. PLoS One 2018, 13, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kader, M.A. A comparison of seed germination calculation formulae and the associated interpretation of resulting data. J. Proc. R. Soc. New South Wales 2005, 138, 65–75. [Google Scholar]
- Lee, S.L.; Lee, D.K. What is the proper way to apply the multiple comparison test? Korean J. Anesthesiol. 2018, 71, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Mlakar, S.G.; Jakop, M.; Bavec, M.; Bavec, F. Allelopathic effects of Amaranthus retroflexus and Amaranthus cruentus extracts on germination of garden cress. Afr. J. Agric. Res. 2012, 7, 1492–1497. [Google Scholar] [CrossRef]
- Konstantinović, B.; Blagojević, M.; Konstantinović, B.; Samardžić, N. Allelopathic effect of weed species Amaranthus retroflexus L. on maize seed germination. Rom. Agric. Res. 2014, 31, 315–321. [Google Scholar] [CrossRef]
- Sheeba, M.A.; Mahesh, B.; Devi Chinmayee, M.; Swapna, T.S.; Mini, I. The allelopathic potential of the weed Amaranthus viridis L. on germination and pigment profile of a pulse crop Vigna radiata (L.) R. Wilsczek. J. Aqu. Biol. Fish 2014, 2, 3344–3350. [Google Scholar]
- Marinov-Serafimov, P. Determination of allelopathic effect of some invasive weed species on germination and initial development of grain legume crops. Pestic. Phytomed. 2010, 25, 251–259. [Google Scholar] [CrossRef]
- Béres, I.; Kazinezi, G. Allelopathic effects of shoot extracts and residues of weeds on field crops. Allelopath. J. 2000, 7, 93–98. [Google Scholar]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Allelopathy in agroecosystems. J. Crop Prod. 2008, 4, 1–41. [Google Scholar] [CrossRef]
- Subtain, M.U.; Hussain, M.; Tabassam, M.A.R.; Ali, M.A.; Ali, M.; Mohsin, M.; Mubushar, M. Role of allelopathy in the growth promotion of plants. J. Agric. Sci. 2014, 2, 141–145. [Google Scholar] [CrossRef]
- Menges, R.M. Allelopathic effects of Palmer Amaranth (Amaranthus palmeri) and other plant residues in soil. Weed Sci. 1987, 35, 339–347. [Google Scholar] [CrossRef]
- Qasem, J.R. Allelopathic effects of Amaranthus retroflexus and Chenopodium murale on vegetable crops. Allelopath. J. 1995, 2, 49–66. [Google Scholar]
- Abbas, T.; Nadeem, M.A.; Tanveer, A.; Chauhan, B.S. Can hormesis of plant-released phytotoxins be used to boost and sustain crop production? Crop Prot. 2017, 93, 69–76. [Google Scholar] [CrossRef]
- Southam, C.M.; Ehrlich, J. Effects of extract of western red-cedar heartwood on certain wood-decaying fungi in culture. Phytopathology 1943, 33, 517–524. [Google Scholar]
- Belz, R.G.; Hurle, K. Dose-response-a challenge for allelopathy? Nonlinearity Biol. Toxicol. Med. 2005, 3, 173–211. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Aro, M.; Hernández-Pérez, R.; Guillén-Sánchez, D.; Torres-Garcia, S. Allelopathic influence of residues from Sphagneticola trilobata on weeds and crops. Planta Daninha 2016, 33, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Prinsloo, G.; Plooy, C.P.D. The allelopathic effects of Amaranthus on seed germination, growth and development of vegetables. Biol. Agric. Hortic. 2018, 34, 268–279. [Google Scholar] [CrossRef]
- Norsworthy, J.K. Allelopathic potential of wild radish. Weed Technol. 2003, 17, 307–313. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Organ | Control (g) | Ground (g) | df | T | P |
|---|---|---|---|---|---|---|
| M | M | |||||
| Chicory | Shoot | 0.54 ± 0.06 | 0.73 ± 0.05 | 18 | 1.92 | 0.072 |
| Root | 0.17 ± 0.02 | 0.17 ± 0.03 | 18 | 0.17 | 0.866 | |
| Oilseed | Shoot | 1.63 ± 0.29 | 1.90 ± 0.38 | 18 | 1.77 | 0.095 |
| Radish | Root | 0.20 ± 0.07 | 0.30 ± 0.12 | 18 | 2.24 | 0.038 |
| Red | Shoot | 0.20 ± 0.04 | 0.10 ± 0.02 | 18 | 2.38 | 0.028 |
| Clover | Root | 0.16 ± 0.03 | 0.03 ± 0.01 | 18 | 4.80 | 0.001 |
| Alfalfa | Shoot | 0.17 ± 0.02 | 0.16 ± 0.04 | 18 | 0.10 | 0.924 |
| Root | 0.10 ± 0.02 | 0.04 ± 0.01 | 18 | 2.60 | 0.024 | |
| Annual | Shoot | 0.26 ± 0.02 | 0.58 ± 0.06 | 18 | 4.95 | <0.001 |
| Rye | Root | 0.29 ± 0.05 | 0.33 ± 0.03 | 18 | 0.68 | 0.506 |
| Species | A. hybridus Treatment | Mortality Rate (%) | Shoot (g) | MS | F | P | Root (g) | MS | F | P |
|---|---|---|---|---|---|---|---|---|---|---|
| Chicory | 0g | 0 | 1.20 ± 0.63 a | 0.56 ± 0.04 a | ||||||
| 10g | 0 | 0.68 ± 0.09 b | 0.97 | 12.40 | < 0.001 | 0.29 ± 0.05 b | 0.19 | 7.87 | 0.003 | |
| 20g | 60 | 0.51 ± 0.21 b | 0.46 ± 0.12 ab | |||||||
| Oilseed | 0g | 0 | 2.99 ± 0.09 a | 0.88 ± 0.08 a | ||||||
| Radish | 10g | 0 | 2.01 ± 0.18 b | 5.27 | 37.05 | < 0.001 | 0.72 ± 0.06 ab | 0.32 | 8.79 | 0.001 |
| 20g | 0 | 1.57 ± 0.57 c | 0.52 ± 0.04 b | |||||||
| Red | 0g | 0 | 0.43 ± 0.05 | 0.29 ± 0.04 | ||||||
| Clover | 10g | 0 | 0.32 ± 0.04 | 0.03 | 2.09 | 0.147 | 0.23 ± 0.02 | 0.03 | 3.26 | 0.057 |
| 20g | 40 | 0.35 ± 0.04 | 0.18 ± 0.03 | |||||||
| Alfalfa | 0g | 0 | 0.59 ± 0.03 a | 0.27 ± 0.04 | ||||||
| 10g | 0 | 0.38 ± 0.06 b | 0.16 | 8.50 | 0.002 | 0.24 ± 0.02 | 0.01 | 1.18 | 0.325 | |
| 20g | 30 | 0.36 ± 0.03 b | 0.20 ± 0.02 | |||||||
| Annual | 0g | 0 | 0.81 ± 0.07 a | 0.50 ± 0.07 | ||||||
| Rye | 10g | 0 | 0.61 ± 0.09 b | 0.16 | 3.36 | 0.050 | 0.51 ± 0.07 | 0.06 | 1.14 | 0.334 |
| 20g | 0 | 0.57 ± 0.04 b | 0.38 ± 0.06 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
VanVolkenburg, H.; Guinel, F.C.; Vasseur, L. Impacts of Smooth Pigweed (Amaranthus hybridus) on Cover Crops in Southern Ontario. Agronomy 2020, 10, 529. https://doi.org/10.3390/agronomy10040529
VanVolkenburg H, Guinel FC, Vasseur L. Impacts of Smooth Pigweed (Amaranthus hybridus) on Cover Crops in Southern Ontario. Agronomy. 2020; 10(4):529. https://doi.org/10.3390/agronomy10040529
Chicago/Turabian StyleVanVolkenburg, Heather, Frédérique C. Guinel, and Liette Vasseur. 2020. "Impacts of Smooth Pigweed (Amaranthus hybridus) on Cover Crops in Southern Ontario" Agronomy 10, no. 4: 529. https://doi.org/10.3390/agronomy10040529
APA StyleVanVolkenburg, H., Guinel, F. C., & Vasseur, L. (2020). Impacts of Smooth Pigweed (Amaranthus hybridus) on Cover Crops in Southern Ontario. Agronomy, 10(4), 529. https://doi.org/10.3390/agronomy10040529