Response of Soil Bacterial Community and Pepper Plant Growth to Application of Bacillus thuringiensis KNU-07

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Bioinformatics Approach for Designing a Strain-Specific Primer for KNU-07
2.2.1. Designing a Primer to Target a Unique Sequence of KNU-07
2.2.2. In Vitro Validation of the Strain-Specific Primer Pair
2.3. In Vitro Plant Growth-Promoting (PGP) Traits Assay
2.4. Greenhouse Experiment
2.4.1. Plant Material and Bacterial Strain Preparation
2.4.2. In Vivo Assay
2.4.3. DNA Extraction from Pure Cultures and Soil Samples
2.4.4. DNA Library Preparation and Amplicon Sequencing
2.4.5. Continuous Tracking of KNU-07 Using qPCR
2.5. Nucleotide Accession Numbers
2.6. Statistical Analysis
3. Results
3.1. In Silico and in Vitro PCR Verification of KNU-07-Specific Primer Pairs
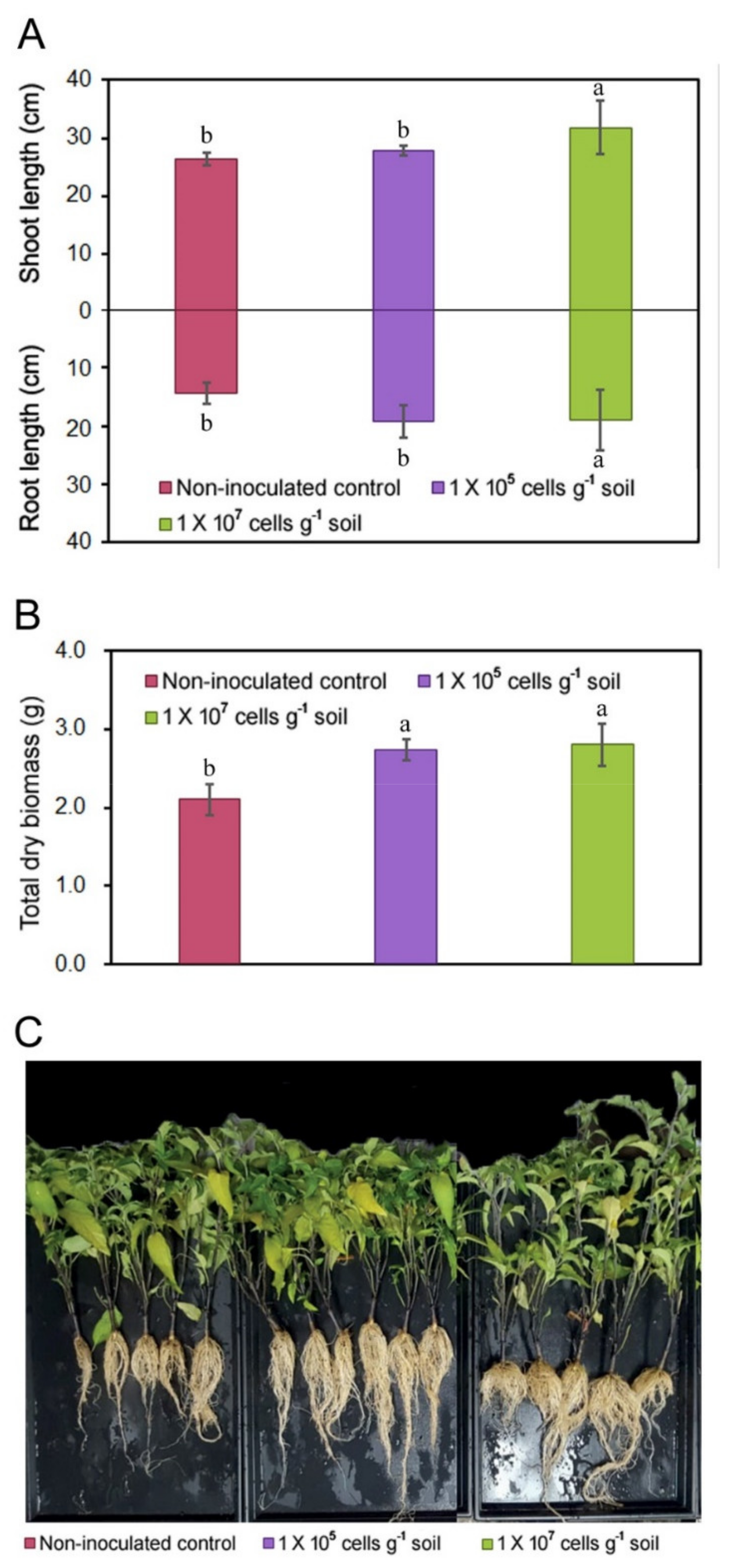
3.2. PGP Activity of KNU-07
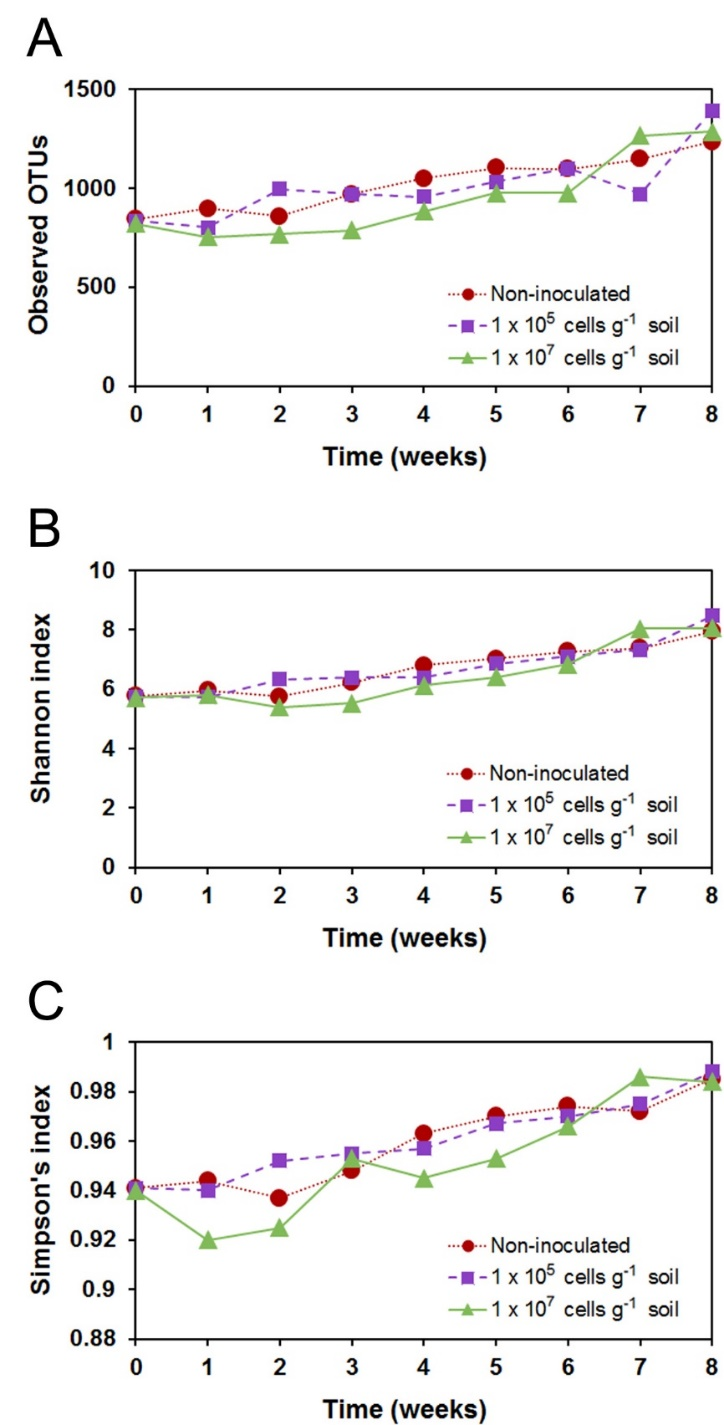
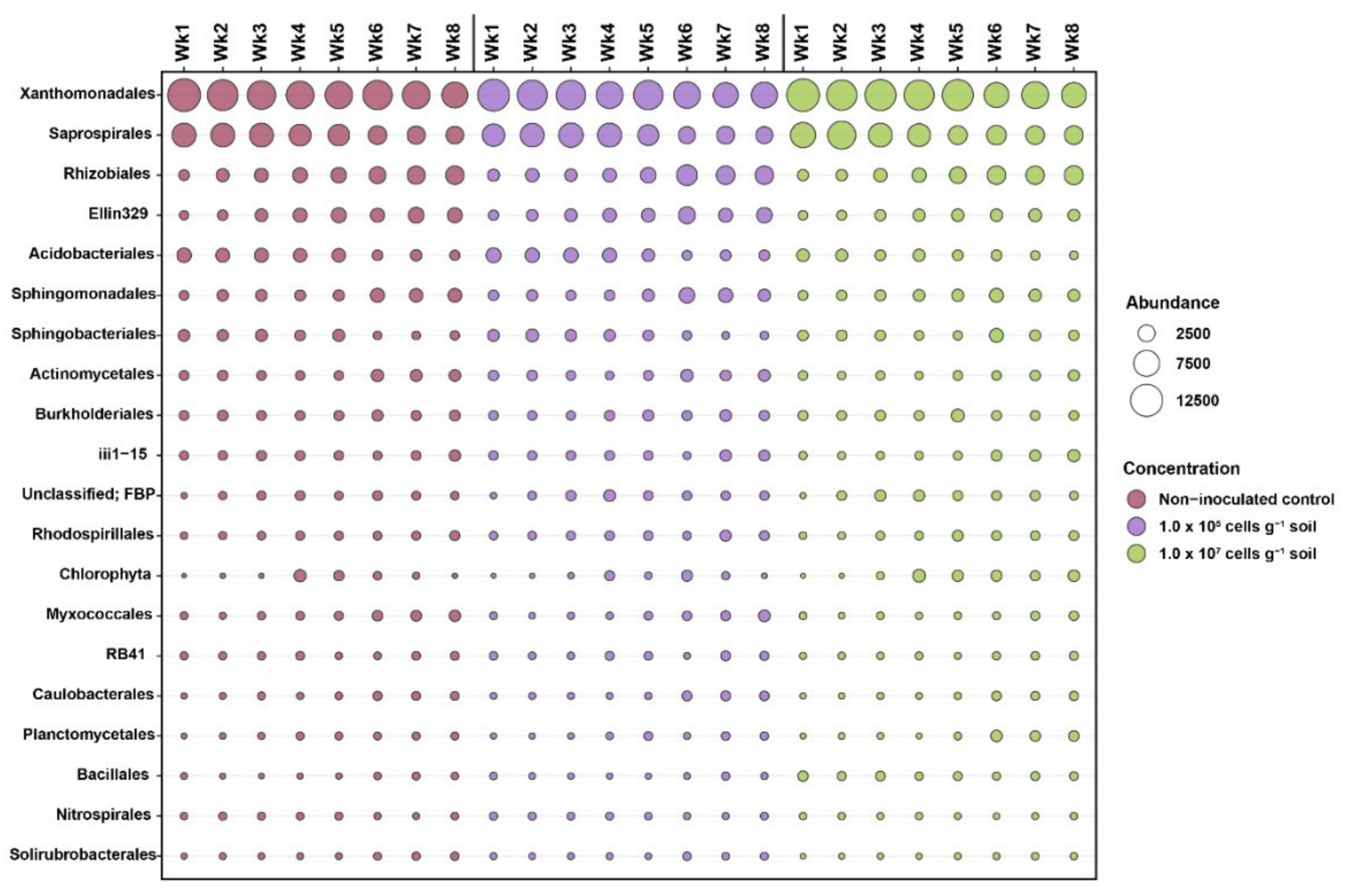
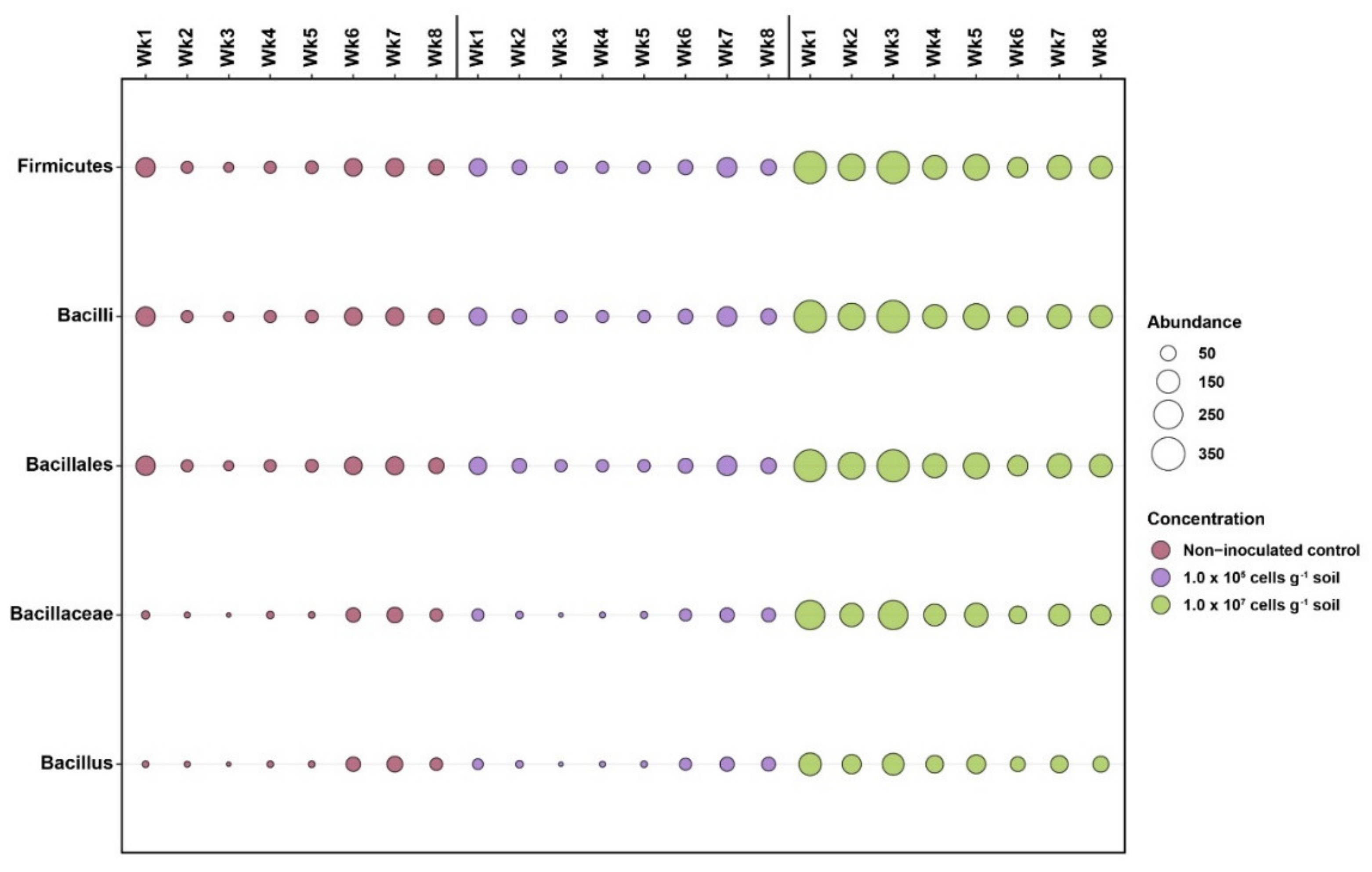
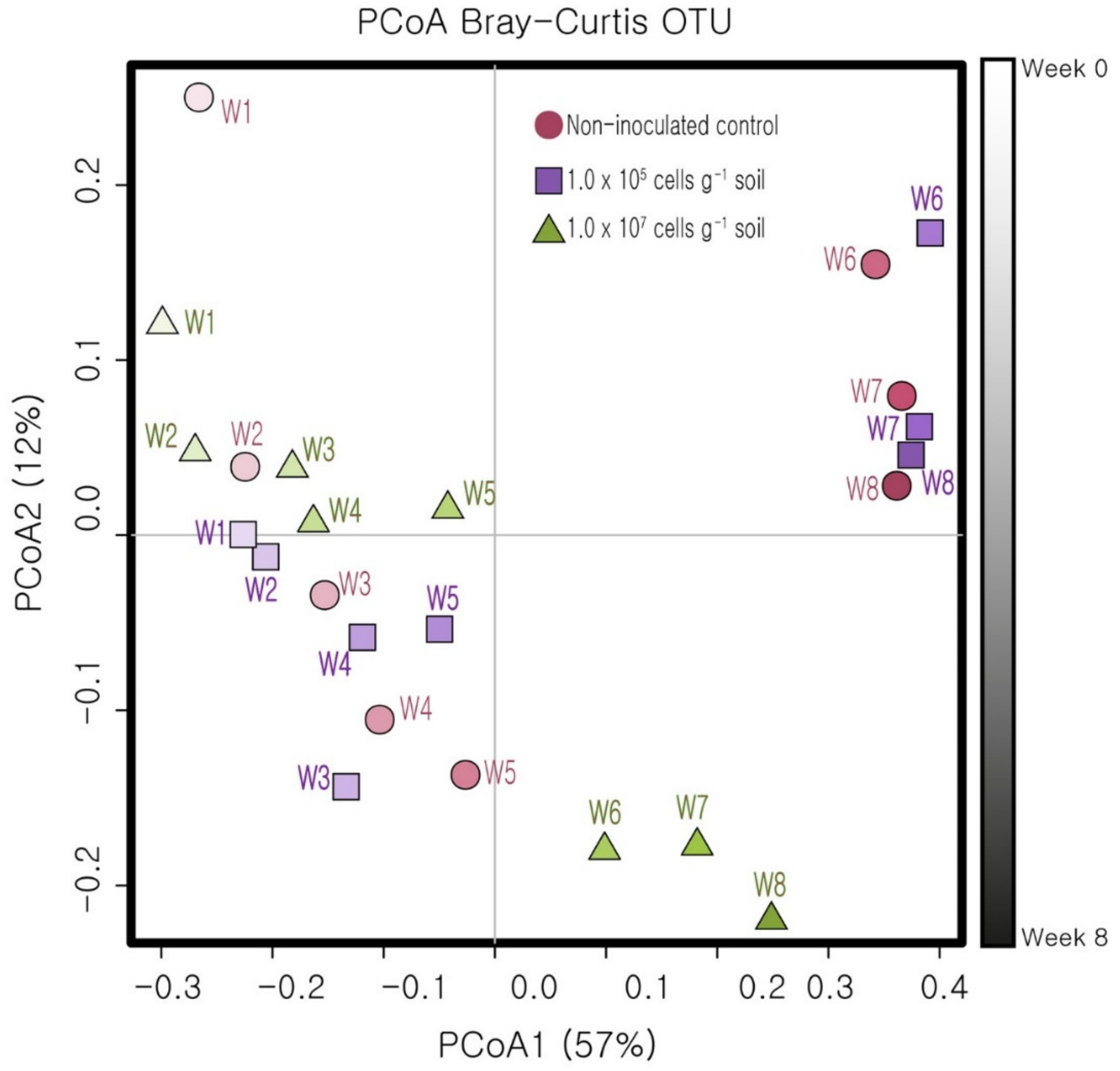
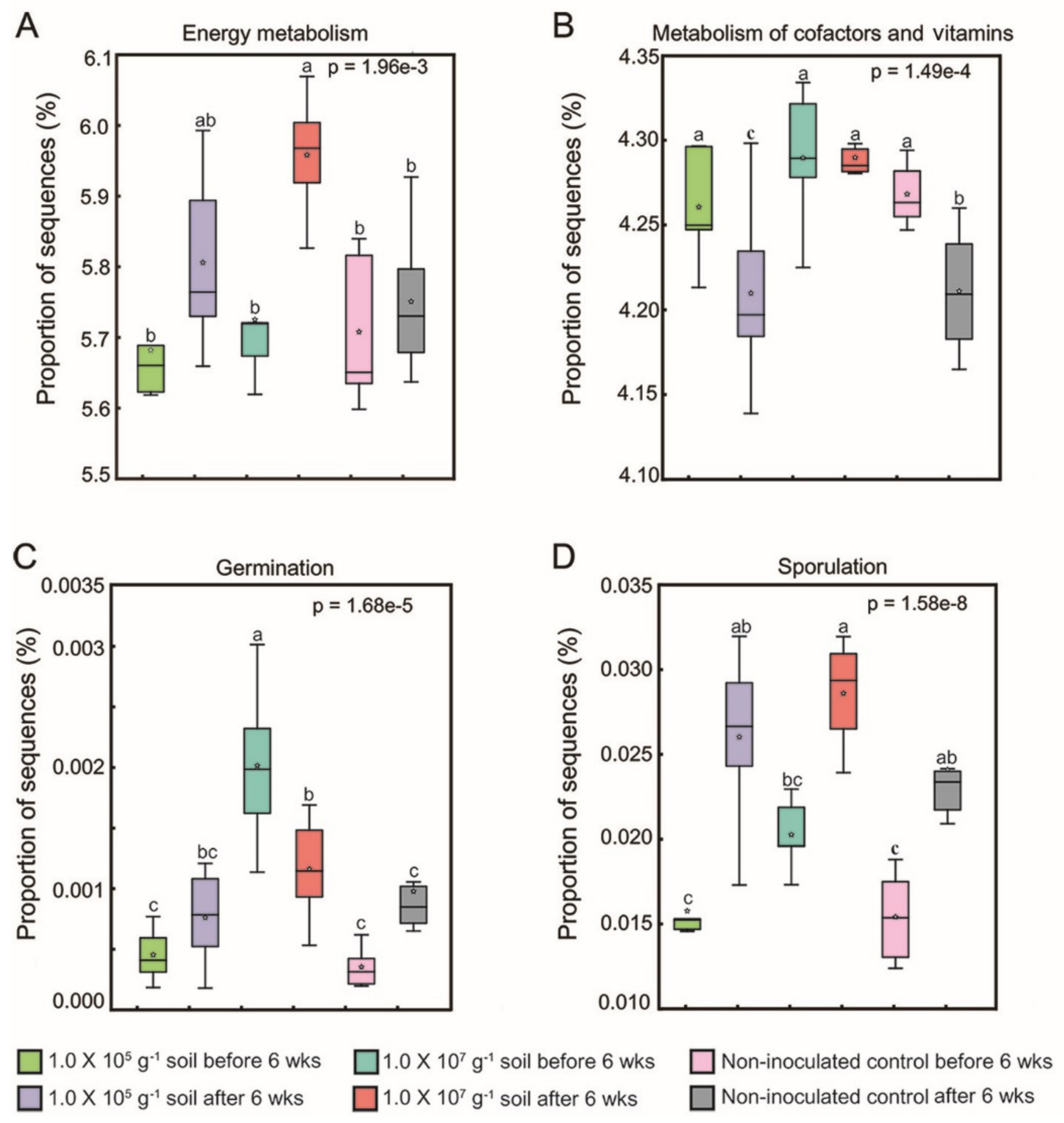
3.3. Response of the Soil Bacterial Community to KNU-07
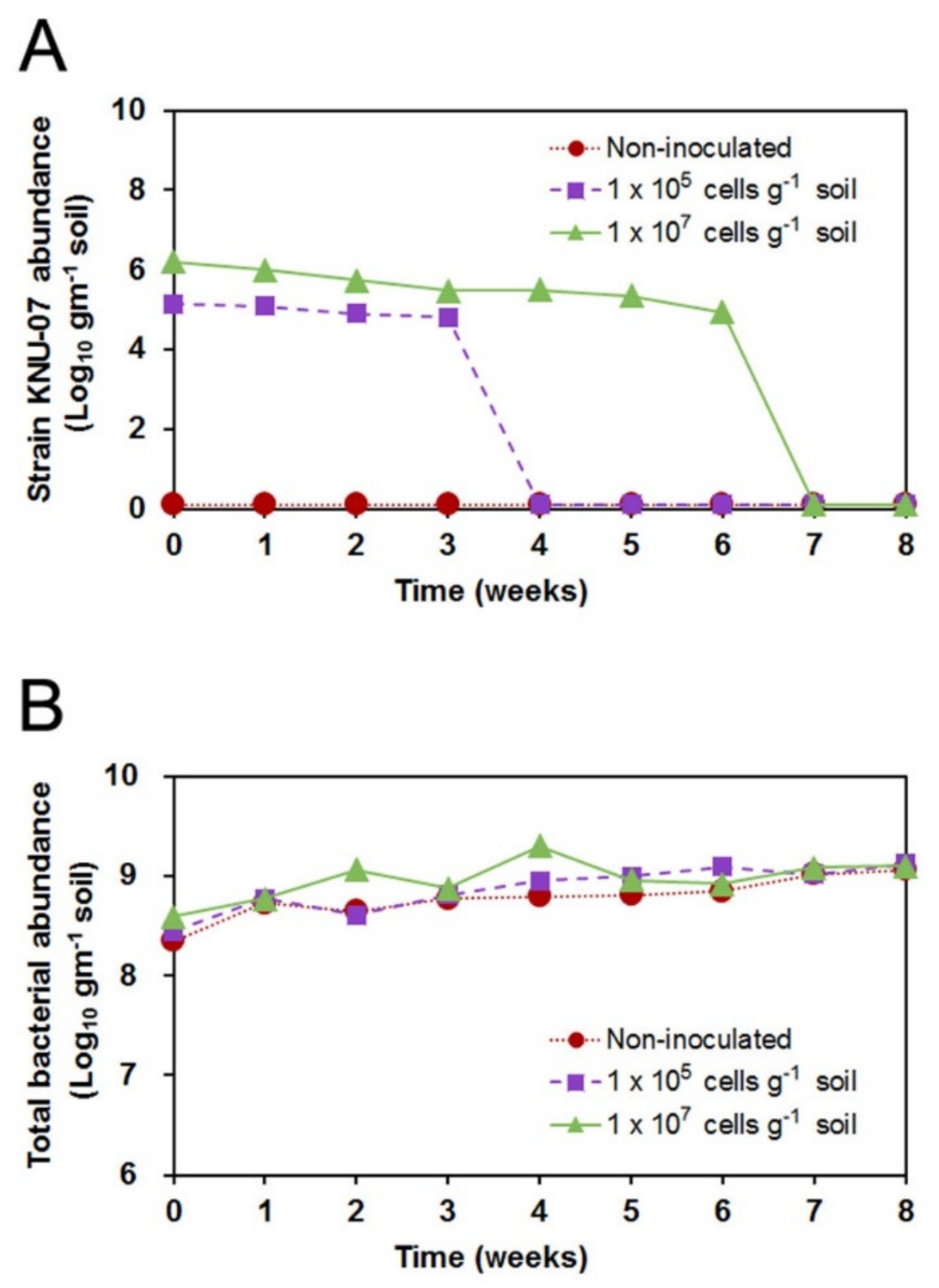
3.4. Tracking of KNU-07 Population Using qPCR
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Anusha, B.G.; Gopalakrishnan, S.; Naik, M.K.; Sharma, M. Evaluation of Streptomyces spp. and Bacillus spp. for biocontrol of Fusarium wilt in chickpea (Cicer arietinum L.). Arch. Phytopathol. Plant Prot. 2019, 52, 417–442. [Google Scholar] [CrossRef]
- Gamez, R.; Cardinale, M.; Montes, M.; Ramirez, S.; Schnell, S.; Rodriguez, F. Screening, plant growth promotion and root colonization pattern of two rhizobacteria (Pseudomonas fluorescens Ps006 and Bacillus amyloliquefaciens Bs006) on banana cv. Williams (Musa acuminata Colla). Microbiol. Res. 2019, 220, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Mhatre, P.H.; Karthik, C.; Kadirvelu, K.; Divya, K.L.; Venkatasalam, E.P.; Srinivasan, S.; Ramkumar, G.; Saranya, C.; Shanmuganathan, R. Plant growth promoting rhizobacteria (PGPR): A potential alternative tool for nematodes bio-control. Biocatal. Agric. Biotechnol. 2019, 17, 119–128. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.W.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [Green Version]
- Shafi, J.; Tian, H.; Ji, M. Bacillus species as versatile weapons for plant pathogens: A review. Biotechnol. Biotechnol. Equip. 2017, 31, 446–459. [Google Scholar] [CrossRef] [Green Version]
- Babu, A.G.; Kim, J.D.; Oh, B.T. Enhancement of heavy metal phytoremediation by Alnus firma with endophytic Bacillus thuringiensis GDB-1. J. Hazard. Mater. 2013, 250, 477–483. [Google Scholar] [CrossRef]
- Azizoglu, U. Bacillus thuringiensis as a Biofertilizer and Biostimulator: A Mini-Review of the Little-Known Plant Growth-Promoting Properties of Bt. Curr. Microbiol. 2019, 76, 1379–1385. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Sidhu, G.K.; Datta, S.; Dhanjal, D.S.; Koul, B.; Janeja, H.S.; Singh, J. Plant growth promoting rhizobacteria from heavy metal contaminated soil promote growth attributes of Pisum sativum L. Biocatal. Agric. Biotechnol. 2019, 17, 665–671. [Google Scholar] [CrossRef]
- Wang, M.; Chen, S.; Han, Y.; Chen, L.; Wang, D. Responses of soil aggregates and bacterial communities to soil-Pb immobilization induced by biofertilizer. Chemosphere 2019, 220, 828–836. [Google Scholar] [CrossRef]
- You, C.; Zhang, C.; Kong, F.; Feng, C.; Wang, J. Comparison of the effects of biocontrol agent Bacillus subtilis and fungicide metalaxyl-mancozeb on bacterial communities in tobacco rhizospheric soil. Ecol. Eng. 2016, 91, 119–125. [Google Scholar] [CrossRef]
- Wang, T.; Liu, M.Q.; Li, H.X. Inoculation of phosphate-solubilizing bacteria Bacillus thuringiensis B1 increases available phosphorus and growth of peanut in acidic soil. Acta Agric. Scand. Sect. B Soil Plant Sci. 2014, 64, 252–259. [Google Scholar]
- Hollensteiner, J.; Wemheuer, F.; Harting, R.; Kolarzyk, A.M.; Diaz Valerio, S.M.; Poehlein, A.; Brzuszkiewicz, E.B.; Nesemann, K.; Braus-Stromeyer, S.A.; Braus, G.H.; et al. Bacillus thuringiensis and Bacillus weihenstephanensis inhibit the growth of phytopathogenic Verticillium species. Front. Microbiol. 2017, 7, 2171. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Bano, A.; Rahman, M.A.; Guo, J.; Kang, Z.; Babar, M.A. Comparative physiological and metabolic analysis reveals a complex mechanism involved in drought tolerance in chickpea (Cicer arietinum L.) induced by PGPR and PGRs. Sci. Rep. 2019, 9, 2097. [Google Scholar] [CrossRef]
- Gange, A.C.; Gadhave, K.R. Plant growth-promoting rhizobacteria promote plant size inequality. Sci. Rep. 2018, 8, 13828. [Google Scholar] [CrossRef]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Cao, Y.; Pi, H.; Chandrangsu, P.; Li, Y.; Wang, Y.; Zhou, H.; Xiong, H.; Helmann, J.D.; Cai, Y. Antagonism of Two Plant-Growth Promoting Bacillus velezensis Isolates Against Ralstonia solanacearum and Fusarium oxysporum. Sci. Rep. 2018, 8, 4360. [Google Scholar] [CrossRef]
- Mosimann, C.; Oberhänsli, T.; Ziegler, D.; Nassal, D.; Kandeler, E.; Boller, T.; Mäder, P.; Thonar, C. Tracing of two Pseudomonas strains in the root and rhizoplane of maize, as related to their plant growth-promoting effect in contrasting soils. Front. Microbiol. 2017, 7, 2150. [Google Scholar] [CrossRef] [Green Version]
- Aufrecht, J.A.; Timm, C.M.; Bible, A.; Morrell-Falvey, J.L.; Pelletier, D.A.; Doktycz, M.J.; Retterer, S.T. Microfluidics: Quantifying the Spatiotemporal Dynamics of Plant Root Colonization by Beneficial Bacteria in a Microfluidic Habitat (Adv. Biosys. 6/2018). Adv. Biosyst. 2018, 2, 1800048. [Google Scholar] [CrossRef]
- Rilling, J.I.; Acuña, J.J.; Nannipieri, P.; Cassan, F.; Maruyama, F.; Jorquera, M.A. Current opinion and perspectives on the methods for tracking and monitoring plant growth‒promoting bacteria. Soil Biol. Biochem. 2019, 130, 205–219. [Google Scholar] [CrossRef]
- Goulart, M.C.; Cueva-Yesquén, L.G.; Hidalgo Martinez, K.J.; Attili-Angelis, D.; Fantinatti-Garboggini, F. Comparison of specific endophytic bacterial communities in different developmental stages of Passiflora incarnata using culture-dependent and culture-independent analysis. Microbiologyopen 2019, 8, e896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraková, L.; Šoltys, K.; Otlewska, A.; Pietrzak, K.; Purkrtová, S.; Savická, D.; Puškárová, A.; Bučková, M.; Szemes, T.; Budiš, J.; et al. Comparison of methods for identification of microbial communities in book collections: Culture-dependent (sequencing and MALDI-TOF MS) and culture-independent (Illumina MiSeq). Int. Biodeterior. Biodegrad. 2018, 131, 51–59. [Google Scholar] [CrossRef]
- Stets, M.I.; Alqueres, S.; Souza, E.M.; Pedrosa, F.O.; Schmid, M.; Hartmann, A.; Cruz, L.M. Quantification of Azospirillum brasilense FP2 bacteria in wheat roots by strain-specific quantitative PCR. Appl. Environ. Microbiol. 2015, 81, 6700–6709. [Google Scholar] [CrossRef] [Green Version]
- Mendis, H.C.; Thomas, V.P.; Schwientek, P.; Salamzade, R.; Chien, J.T.; Waidyarathne, P.; Kloepper, J.; De La Fuente, L. Strain-specific quantification of root colonization by plant growth promoting rhizobacteria Bacillus firmus I-1582 and Bacillus amyloliquefaciens QST713 in non-sterile soil and field conditions. PLoS ONE 2018, 13, e0193119. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Coissac, E.; Zundel, S.; Riaz, T.; Shehzad, W.; Bessière, J.; Taberlet, P.; Pompanon, F. An in silico approach for the evaluation of DNA barcodes. BMC Genom. 2010, 11, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T.; Robert, V. Methods for isolation, phenotypic characterization and maintenance of yeasts. In The Yeasts; Elsevier: London, UK; Amsterdam, The Netherlands, 2011; ISBN 9780444521491. [Google Scholar]
- Pérez-Miranda, S.; Cabirol, N.; George-Téllez, R.; Zamudio-Rivera, L.S.; Fernández, F.J. O-CAS, a fast and universal method for siderophore detection. J. Microbiol. Methods 2007, 31, 446–459. [Google Scholar] [CrossRef]
- Kyaw, A.; Maung-U, K.; Toe, T. Determination of inorganic phosphate with molybdate and Triton X-100 without reduction. Anal. Biochem. 1985, 145, 230–234. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- CLARKE, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SAS Institute. SAS Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2012. [Google Scholar]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagele, S.B.; Lee, H.G.; Kim, S.W.; Lee, Y.S. Phenazine and 1-undecene producing Pseudomonas chlororaphis subsp. aurantiaca strain KNU17Pc1 for growth promotion and disease suppression in Korean maize cultivars. J. Microbiol. Biotechnol. 2019, 29, 66–78. [Google Scholar]
- Viljoen, J.J.F.; Labuschagne, N.; Fourie, H.; Sikora, R.A. Biological control of the root-knot nematode Meloidogyne incognita on tomatoes and carrots by plant growth-promoting rhizobacteria. Trop. Plant Pathol. 2019, 44, 284–291. [Google Scholar] [CrossRef]
- Vidal-Quist, J.C.; Rogers, H.W.J.; Mahenthiralingam, E.; Berry, C. Bacillus thuringiensis colonises plant roots in a phylogeny-dependent manner. FEMS Microbiol. Ecol. 2013, 86, 474–489. [Google Scholar] [CrossRef] [Green Version]
- Ke, X.; Feng, S.; Wang, J.; Lu, W.; Zhang, W.; Chen, M.; Lin, M. Effect of inoculation with nitrogen-fixing bacterium Pseudomonas stutzeri A1501 on maize plant growth and the microbiome indigenous to the rhizosphere. Syst. Appl. Microbiol. 2019, 42, 248–260. [Google Scholar] [CrossRef]
- Wang, J.; Li, Q.; Xu, S.; Zhao, W.; Lei, Y.; Song, C.; Huang, Z. Traits-based integration of multi-species inoculants facilitates shifts of indigenous soil bacterial community. Front. Microbiol. 2018, 9, 1692. [Google Scholar] [CrossRef]
- Bouskill, N.J.; Lim, H.C.; Borglin, S.; Salve, R.; Wood, T.E.; Silver, W.L.; Brodie, E.L. Pre-exposure to drought increases the resistance of tropical forest soil bacterial communities to extended drought. ISME J. 2013, 7, 384–394. [Google Scholar] [CrossRef]
- Harbison, A.B.; Carson, M.A.; Lamit, L.J.; Basiliko, N.; Bräuer, S.L. A novel isolate and widespread abundance of the candidate alphaproteobacterial order (Ellin 329), in southern Appalachian peatlands. FEMS Microbiol. Lett. 2016, 363, fnw151. [Google Scholar] [CrossRef] [Green Version]
- Van Overbeek, L.; Van Elsas, J.D. Effects of plant genotype and growth stage on the structure of bacterial communities associated with potato (Solanum tuberosum L.). FEMS Microbiol. Ecol. 2008, 64, 283–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degrune, F.; Theodorakopoulos, N.; Colinet, G.; Hiel, M.P.; Bodson, B.; Taminiau, B.; Daube, G.; Vandenbol, M.; Hartmann, M. Temporal dynamics of soil microbial communities below the seedbed under two contrasting tillage regimes. Front. Microbiol. 2017, 8, 1127. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Wang, Z.; Li, N.; Wang, Y.; Feng, J.; Zhang, X. Bacillus amyloliquefaciens B1408 suppresses Fusarium wilt in cucumber by regulating the rhizosphere microbial community. Appl. Soil Ecol. 2019, 136, 55–66. [Google Scholar] [CrossRef]
- He, Y.; Pantigoso, H.A.; Wu, Z.; Vivanco, J.M. Co-inoculation of Bacillus sp. and Pseudomonas putida at different development stages acts as a biostimulant to promote growth, yield and nutrient uptake of tomato. J. Appl. Microbiol. 2019, 127, 196–207. [Google Scholar] [CrossRef]
- Omony, J.; de Jong, A.; Krawczyk, A.O.; Eijlander, R.T.; Kuipers, O.P. Dynamic sporulation gene co-expression networks for Bacillus subtilis 168 and the food-borne isolate Bacillus amyloliquefaciens: A transcriptomic model. Microb. Genom. 2018, 4, e000157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, L.W.; Kuramae, E.E.; Navarrete, A.A.; Van Veen, J.A.; Tsai, S.M. Taxonomical and functional microbial community selection in soybean rhizosphere. ISME J. 2014, 8, 1577–1587. [Google Scholar] [CrossRef]
- Taute, K.M.; Gude, S.; Tans, S.J.; Shimizu, T.S. High-throughput 3D tracking of bacteria on a standard phase contrast microscope. Nat. Commun. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Banik, A.; Mukhopadhaya, S.K.; Sahana, A.; Das, D.; Dangar, T.K. Fluorescence resonance energy transfer (FRET)-based technique for tracking of endophytic bacteria in rice roots. Biol. Fertil. Soils 2016, 52, 277–282. [Google Scholar] [CrossRef]
- Segarra, G.; Puopolo, G.; Porcel-Rodríguez, E.; Giovannini, O.; Pertot, I. Monitoring Lysobacter capsici az78 using strain specific qpcr reveals the importance of the formulation for its survival in vineyards. FEMS Microbiol. Lett. 2016, 363, fnv243. [Google Scholar] [CrossRef] [Green Version]
- Ricketts, C.; Pickler, L.; Maurer, J.; Ayyampalayam, S.; García, M.; Ferguson-Noel, N.M. Identification of strain-specific sequences that distinguish a Mycoplasma gallisepticum vaccine strain from field isolates. J. Clin. Microbiol. 2017, 55, 244–252. [Google Scholar] [CrossRef] [Green Version]
- Coy, R.M.; Held, D.W.; Kloepper, J.W. Rhizobacterial colonization of bermudagrass by Bacillus spp. in a Marvyn loamy sand soil. Appl. Soil Ecol. 2019, 141, 10–17. [Google Scholar] [CrossRef]
- Khabbaz, S.E.; Abbasi, P.A. Isolation, characterization, and formulation of antagonistic bacteria for the management of seedlings damping-off and root rot disease of cucumber. Can. J. Microbiol. 2014, 60, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Andreote, F.D.; De Araújo, W.L.; De Azevedo, J.L.; Van Elsas, J.D.; Da Rocha, U.N.; Van Overbeek, L.S. Endophytic colonization of potato (Solanum tuberosum L.) by a novel competent bacterial endophyte, Pseudomonas putida strain P9, and its effect on associated bacterial communities. Appl. Environ. Microbiol. 2009, 75, 3396–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishizawa, H.; Kuroda, M.; Inoue, K.; Inoue, D.; Morikawa, M.; Ike, M. Colonization and competition dynamics of plant growth-promoting/inhibiting Bacteria in the phytosphere of the Duckweed Lemna minor. Microb. Ecol. 2019, 77, 440–450. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5’→3’) | Reference |
|---|---|---|
| Strain-specific primers | ||
| KNU07F | TGCTCTTTCTGGATTATTCCTTGAG | This study |
| KNU07R | CATCCTTTTGTAGAAGGTATTGCCA | This study |
| Universal primers | ||
| 27F | AGAGTTTGATCMTGGCTCAG | Lane, 1991 |
| 1492R | TACGGYTACCTTGTTACGACTT | Lane, 1991 |
| 515F | GTGCCAGCMGCCGCGG | Lane, 1991 |
| 907R | CCGTCAATTCMTTTRAGTTT | Lane, 1991 |
| Category | Primer | Taxonomic Level | Total Taxa | Number of Mismatches per Primer | ||
|---|---|---|---|---|---|---|
| Perfect Match | 1 Mismatch | 2 Mismatches | ||||
| Bacterial 16S rRNA gene | 27F/1492R | Phylum | 52 | 26 (50%) | 35 (67%) | 41 (79%) |
| Class | 81 | 48(59%) | 63 (78%) | 70 (86%) | ||
| Order | 173 | 114 (66%) | 140 (81%) | 148 (86%) | ||
| Family | 367 | 262 (71%) | 313 (85%) | 323 (88%) | ||
| Genus | 1130 | 830 (73%) | 993 (88%) | 1019 (90%) | ||
| Species | 3747 | 2843 (76%) | 3331 (89%) | 3453 (92%) | ||
| KNU-07 unique region | KNU07F/KNU07R | Phylum | 52 | 1* | 1* | 1* |
| Class | 81 | 1* | 1* | 1* | ||
| Order | 173 | 1* | 1* | 1* | ||
| Family | 367 | 1* | 1* | 1* | ||
| Genus | 1130 | 1* | 1* | 1* | ||
| Species | 3747 | 1* | 1* | 1* | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, H.; Tagele, S.B.; Pham, H.Q.; Kim, M.-C.; Choi, S.-D.; Kim, M.-J.; Park, Y.-J.; Ibal, J.C.; Park, G.-S.; Shin, J.-H. Response of Soil Bacterial Community and Pepper Plant Growth to Application of Bacillus thuringiensis KNU-07. Agronomy 2020, 10, 551. https://doi.org/10.3390/agronomy10040551
Jo H, Tagele SB, Pham HQ, Kim M-C, Choi S-D, Kim M-J, Park Y-J, Ibal JC, Park G-S, Shin J-H. Response of Soil Bacterial Community and Pepper Plant Growth to Application of Bacillus thuringiensis KNU-07. Agronomy. 2020; 10(4):551. https://doi.org/10.3390/agronomy10040551
Chicago/Turabian StyleJo, HyungWoo, Setu Bazie Tagele, Huy Quang Pham, Min-Chul Kim, Seung-Dae Choi, Min-Ji Kim, Yeong-Jun Park, Jerald Conrad Ibal, Gun-Seok Park, and Jae-Ho Shin. 2020. "Response of Soil Bacterial Community and Pepper Plant Growth to Application of Bacillus thuringiensis KNU-07" Agronomy 10, no. 4: 551. https://doi.org/10.3390/agronomy10040551
APA StyleJo, H., Tagele, S. B., Pham, H. Q., Kim, M. -C., Choi, S. -D., Kim, M. -J., Park, Y. -J., Ibal, J. C., Park, G. -S., & Shin, J. -H. (2020). Response of Soil Bacterial Community and Pepper Plant Growth to Application of Bacillus thuringiensis KNU-07. Agronomy, 10(4), 551. https://doi.org/10.3390/agronomy10040551