Paecilomyces variotii as A Plant-Growth Promoter in Horticulture


Abstract
:
1. Introduction
2. Materials and Methods
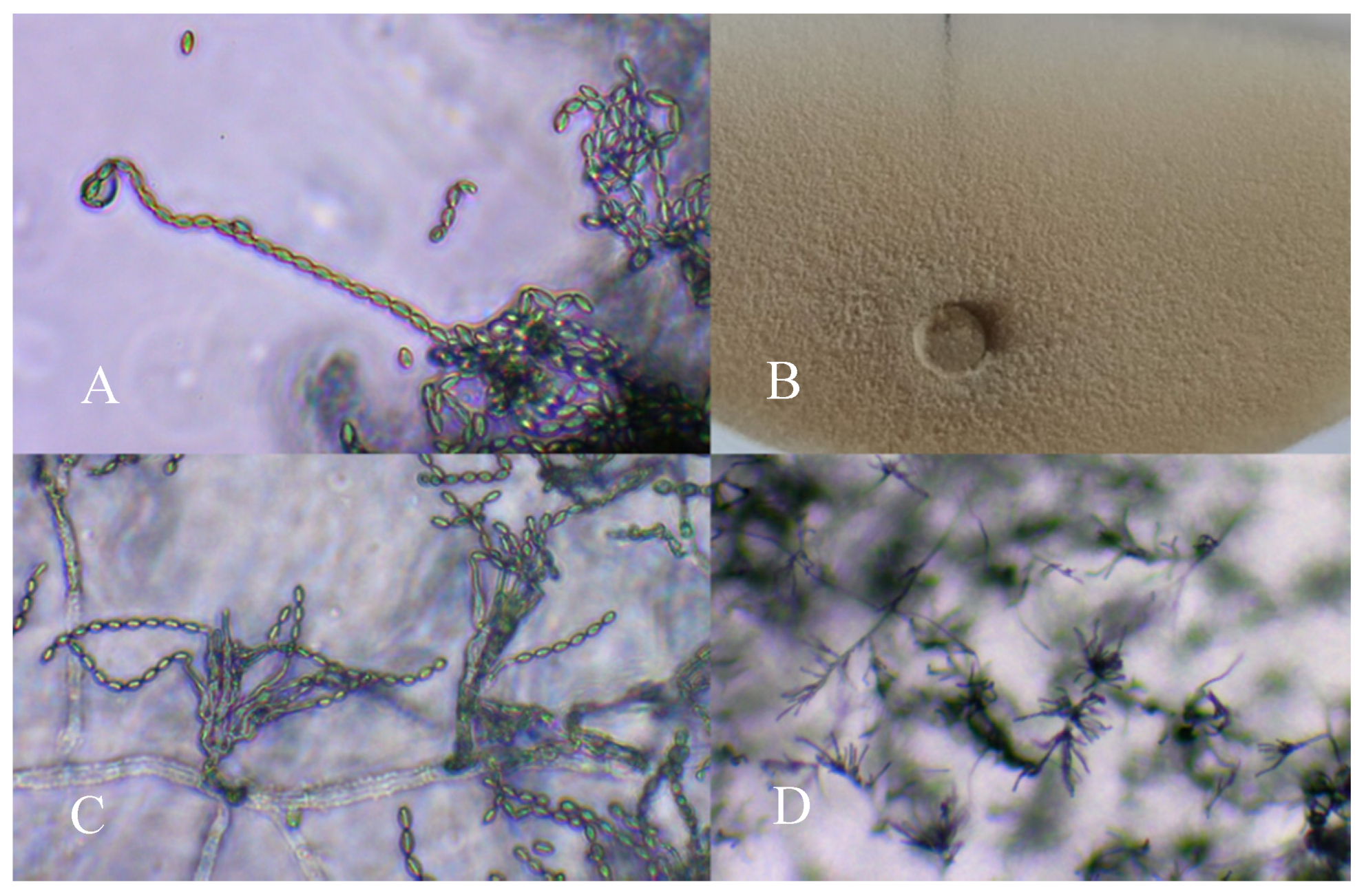
2.1. Isolation of P. variotii from Plant Roots
2.2. Analysis of Plant Growth-Promoting Attributes
2.3. Mass Production of P. variotii on Solid Substrates
2.4. Analysis of Effects of P. variotii on Seed Germination under Laboratory Conditions
2.5. Analysis of Promoter Effects of P. variotii on Pepper and Tomato Seedlings: Experiment 1
2.6. Analysis of Effects of Applying Different Doses of P. variotii to Tomatoes: Experiment 2
2.7. Statistical Analysis
3. Results
3.1. Mass Production of P. variotii on Solid Substrates
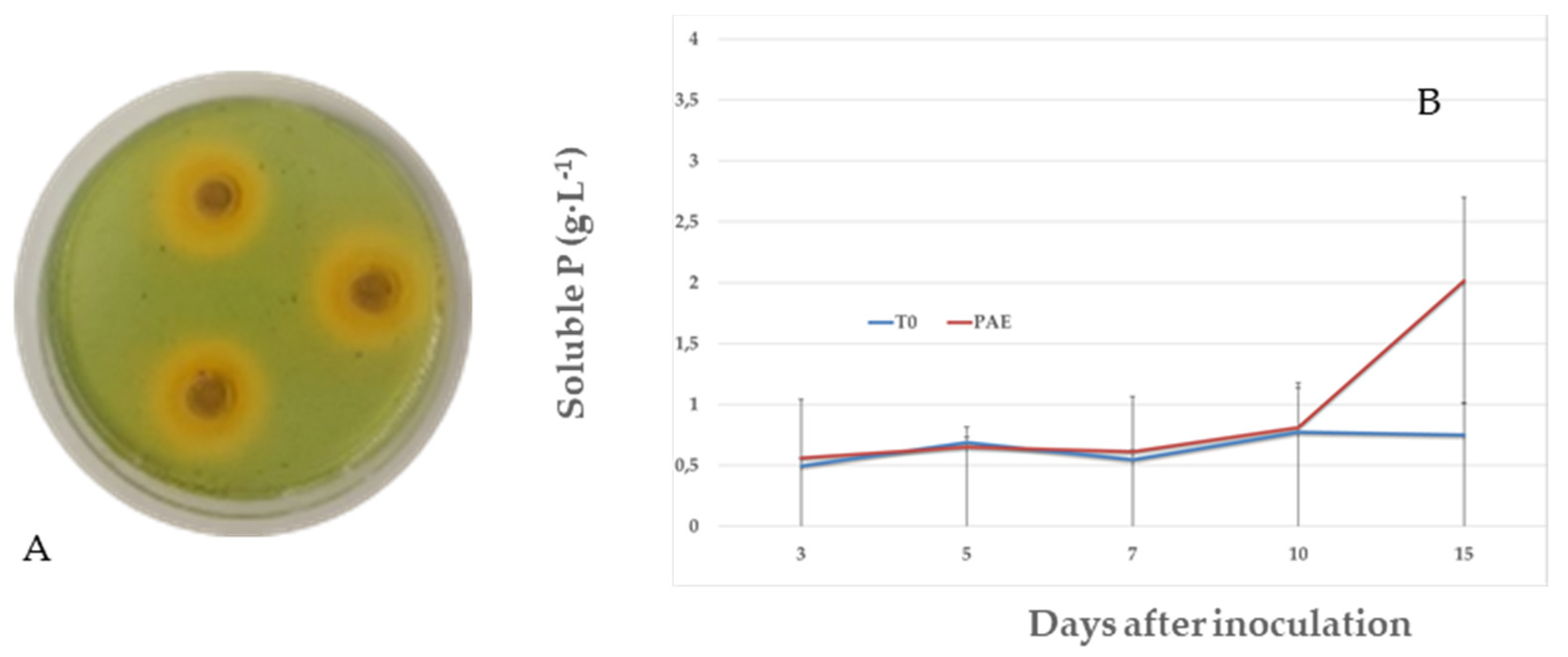
3.2. Plant Growth-Promoting Characteristics of P. variotii: Siderophore Production, IAA and P Solubilization
3.3. P. variotii Inoculation Effects on Tomato and Pepper Seed Germination
3.4. Promoter Effects of P. variotii Isolates on Tomato and Pepper Seedlings: Experiment 1
3.5. Effects of Dose of Application of P. variotii Isolates on Tomato Seedlings and Transplanted Plants: Experiment 2
4. Discussion and Conclusions
5. Patents
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nguyen, T.T.T.; Paul, N.C.; Lee, H.B. Characterization of Paecilomyces variotii and Talaromyces amestolkiae in Korea Based on the Morphological Characteristics and Multigene Phylogenetic Analyses. Mycobiology 2016, 44, 248–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mioso, R.; Marante, F.J.T.; De Laguna, I.H.B. The Chemical Diversity of the Ascomycete Fungus Paecilomyces variotii. Appl. Biochem. Biotechnol. 2015, 177, 781–791. [Google Scholar] [CrossRef] [PubMed]
- De Laguna, I.H.B.; Marante, F.J.T.; Mioso, R. Enzymes and bioproducts produced by the ascomycete fungus Paecilomyces variotii. J. Appl. Microbiol. 2015, 119, 1455–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akinfala, T.O.; Houbraken, J.; Sulyok, M.; Adedeji, A.R.; Odebode, A.C.; Krska, R.; Ezekiel, C.N. Moulds and their secondary metabolites associated with the fermentation and storage of two cocoa bean hybrids in Nigeria. Int. J. Food Microbiol. 2020, 316, 108490. [Google Scholar] [CrossRef] [PubMed]
- Lòpez-Fernàndez, S.; Campisano, A.; Schulz, B.J.; Steinert, M.; Stadler, M.; Surup, F. Viriditins from Byssochlamys spectabilis, their stereochemistry and biosynthesis. Tetrahedron Lett. 2020, 61, 151446. [Google Scholar] [CrossRef]
- Al-Qasim, M.; Abu-Gharbieh, W.; Assas, K. Nematophagal ability of Jordanian isolates of Paecilomyces variotii on the root-knot nematode Meloidogyne javanica. Nematol. Mediterr. 2009, 37, 53–57. [Google Scholar]
- Perveen, Z.; Shahzad, S. A comparative study of the efficacy of Paecilomyces species against root-knot nematode Meloidogyne incognita. Pak. J. Nematol. 2013, 31, 125–135. [Google Scholar]
- Ahmad, R.Z.; Sidi, B.B.; Endrawati, D.; Ekawasti, F. Paecilomyces lilacinus and P. variotii as a predator of nematode and trematode eggs. IOP Conf. Ser. Earth Environ. Sci. 2019, 299, 012056. [Google Scholar] [CrossRef]
- Rodrigo, S.; Santamaria, O.; Halecker, S.; Lledó, S.; Stadler, M. Antagonism betweenByssochlamys spectabilis(anamorphPaecilomyces variotii) and plant pathogens: Involvement of the bioactive compounds produced by the endophyte. Ann. Appl. Boil. 2017, 171, 464–476. [Google Scholar] [CrossRef]
- Mohammadi, S.; Soltani, J.; Piri, K. Soilborne and invertebrate pathogenic Paecilomyces species show activity against pathogenic fungi and bacteria. J. Crop. Prot. 2016, 5, 377–387. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.M.; Wang, J.N.; Wang, B.G. Oxepine-containing diketopiperazine alkaloids from the algal-derived endophytic fungus Paecilomyces variotii EN-291. Helv. Chim. Acta 2015, 98, 800–804. [Google Scholar] [CrossRef]
- Hajano, J.U.D.; Lodhi, M.; Pathan, M.A.; Khanzada, A.; Shah, G.S. In-vitro evaluation of fungicides, plant extracts and bio-control agents against rice blast pathogen Magnaporthe oryzae couch. Pak. J. Bot. 2012, 44, 1775–1778. [Google Scholar]
- Nakajima, M.; Itoi, K.; Takamatsu, Y.; Sato, S.; Furukawa, Y.; Furuya, K.; Honma, T.; Kadotani, J.; Kozasa, M.; Haneishi, T. Cornexistin: A new fungal metabolite with herbicidal activity. J. Antibiot. 1991, 44, 1065–1072. [Google Scholar] [CrossRef]
- Song, X.; Zhang, L.H.; Peng, A.T.; Cheng, B.P.; Ling, J.F. First Report of Paecilomyces variotii Isolated From Citrus Psyllid (Diaphorina citri), the Vector of Huanglongbing of Citrus, in China. Plant Dis. 2016, 100, 2526. [Google Scholar] [CrossRef]
- Hyung, J.J.; Kang, H.; Jong, J.J.; Soo, K.Y. Paecilomyces Variotii Extracts for Preventing and Treating Infections Caused by Fish Pathogenic Microorganisms. KR Patent 2013051523, 2013. [Google Scholar]
- García-Peña, E.; Hernández, S.; Auria, R.; Revah, S. Correlation of Biological Activity and Reactor Performance in Biofiltration of Toluene with the Fungus Paecilomyces variotii CBS115145. Appl. Environ. Microbiol. 2005, 71, 4280–4285. [Google Scholar] [CrossRef] [Green Version]
- García-Peña, E.; Ortiz, I.; Hernandez, S.; Revah, S. Biofiltration of BTEX by the fungus Paecilomyces variotii. Int. Biodeterior. Biodegrad. 2008, 62, 442–447. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, G.; Cai, H.; Shi, P.; Chang, W.; Zhang, S.; Zheng, A.; Xie, Q.; Ma, J. Paecilomyces variotii: A Fungus Capable of Removing Ammonia Nitrogen and Inhibiting Ammonia Emission from Manure. PLoS ONE 2016, 11, e0158089. [Google Scholar] [CrossRef]
- Steiner, B.; Aquino, V.R.; Paz, A.A.; Silla, L.M.D.R.; Zavascki, A.; Goldani, L. Paecilomyces variotii as an Emergent Pathogenic Agent of Pneumonia. Case Rep. Infect. Dis. 2013, 2013, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Santamaria, O.; Lledó, S.; Rodrigo, S.; Poblaciones, M.J. Effect of Fungal Endophytes on Biomass Yield, Nutritive Value and Accumulation of Minerals in Ornithopus compressus. Microb. Ecol. 2017, 74, 841–852. [Google Scholar] [CrossRef]
- Lu, C.; Liu, H.; Jiang, D.; Wang, L.; Jiang, Y.; Tang, S.; Hou, X.; Han, X.; Liu, Z.; Zhang, M.; et al. Paecilomyces variotii extracts (ZNC) enhance plant immunity and promote plant growth. Plant Soil 2019, 441, 383–397. [Google Scholar] [CrossRef]
- Sánchez-Montesinos, B.; Diánez, F.; Moreno-Gavíra, A.; Gea, F.J.; Santos, M. Plant Growth Promotion and Biocontrol of Pythium ultimum by Saline Tolerant Trichoderma Isolates under Salinity Stress. Int. J. Environ. Res. Public Health 2019, 16, 2053. [Google Scholar] [CrossRef] [Green Version]
- Raja, N. Biopesticides and Biofertilizers: Ecofriendly Sources for Sustainable Agriculture. J. Fertil. Pestic. 2013, 4, e112. [Google Scholar] [CrossRef] [Green Version]
- Adesemoye, A.; Kloepper, J.W. Plant–microbes interactions in enhanced fertilizer-use efficiency. Appl. Microbiol. Biotechnol. 2009, 85, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenberghe, L.; Garcia, L.M.B.; Rodrigues, C.; Camara, M.C.; Pereira, G.; De Oliveira, J.; Soccol, C.R. Potential applications of plant probiotic microorganisms in agriculture and forestry. AIMS Microbiol. 2017, 3, 629–648. [Google Scholar] [CrossRef]
- Martínez, F.D.; Santos, M.; Carretero, F.; Marin, F. Trichoderma saturnisporum,a new biological control agent. J. Sci. Food Agric. 2015, 96, 1934–1944. [Google Scholar] [CrossRef]
- Fernando, D.; Santos, M.; Francisco, C.; Francisco, M. Biostimulant Activity of Trichoderma saturnisporum in Melon (Cucumis melo). HortScience 2018, 53, 810–815. [Google Scholar] [CrossRef]
- Smith, S.E.; Manjarrez, M.; Stonor, R.; McNeill, A.; Smith, F.A. Indigenous arbuscular mycorrhizal (AM) fungi contribute to wheat phosphate uptake in a semi-arid field environment, shown by tracking with radioactive phosphorus. Appl. Soil Ecol. 2015, 96, 68–74. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Ansari, M.; Sahoo, R.K.; Tuteja, N. Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Factories 2014, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.R.; Mohiddin, F.A. Trichoderma: Its Multifarious Utility in Crop Improvement. In Crop Improvement through Microbial Biotechnology; Elsevier BV: Amsterdam, The Netherlands, 2018; pp. 263–291. [Google Scholar]
- Murali, M.; Amruthesh, K.N. Plant Growth-promoting Fungus Penicillium oxalicum Enhances Plant Growth and Induces Resistance in Pearl Millet Against Downy Mildew Disease. J. Phytopathol. 2015, 163, 743–754. [Google Scholar] [CrossRef]
- Shimizu, K.; Hossain, M.M.; Kato, K.; Kubota, M.; Hyakumachi, M. Induction of Defense Responses in Cucumber Plants by Using the Cell-free Filtrate of the Plant Growth-Promoting Fungus Penicillium simplicissimum GP17-2. J. Oleo Sci. 2013, 62, 613–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitas, V.; McCartney, N.; Li, N.; Demers, J.; Kim, J.-E.; Kim, H.-S.; Brown, K.M.; Kang, S. Fusarium Oxysporum Volatiles Enhance Plant Growth Via Affecting Auxin Transport and Signaling. Front. Microbiol. 2015, 6, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, H.; Hossain, M.; Kubota, M.; Hyakumachi, M. Involvement of the salicylic acid signaling pathway in the systemic resistance induced in Arabidopsis by plant growth-promoting fungus Fusarium equiseti GF19-1. J. Oleo Sci. 2013, 62, 415–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.S.; Tang, K.; Guo, S.-X. The Plant Growth-Promoting Fungus (PGPF) Alternaria sp. A13 Markedly Enhances Salvia miltiorrhiza Root Growth and Active Ingredient Accumulation under Greenhouse and Field Conditions. Int. J. Mol. Sci. 2018, 19, 270. [Google Scholar] [CrossRef] [Green Version]
- Islam, S.; Akanda, A.M.; Sultana, F.; Hossain, M. Chilli rhizosphere fungus Aspergillus spp. PPA1 promotes vegetative growth of cucumber (Cucumis sativus) plants upon root colonisation. Arch. Phytopathol. Plant Prot. 2013, 47, 1231–1238. [Google Scholar] [CrossRef]
- Naznin, H.A.; Kimura, M.; Miyazawa, M.; Hyakumachi, M. Analysis of Volatile Organic Compounds Emitted by Plant Growth-Promoting Fungus Phoma sp. GS8-3 for Growth Promotion Effects on Tobacco. Microbes Environ. 2012, 28, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of Blue Agar CAS Assay for Siderophore Detection. J. Microbiol. Boil. Educ. 2011, 12, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Lima-Rivera, D.L.; Lopez-Lima, D.; Desgarennes, D.; Velázquez-Rodríguez, A.S.; Carrión, G. Phosphate solubilization by fungi with nematicidal potential. J. Soil Sci. Plant Nutr. 2016, 16, 507–524. [Google Scholar] [CrossRef] [Green Version]
- Fiske, C.H.; Subbarow, Y. The colorimetry determination of phosphorous. J. Biol. Chem. 1925, 66, 375–400. [Google Scholar]
- Murali, M.; Amruthesh, K.N.; Sudisha, J.S.; Niranjana, R.; Shetty, H.S. Screening for plant growth promoting fungi and their ability for growth promotion and induction of resistance in pearl millet against downy mildew disease. J. Phytol. 2012, 4, 30–36. [Google Scholar]
- Dickson, A.; Leaf, A.L.; Hosner, J.F. Quality Appraisal of White Spruce and White Pine Seedling Stock in Nurseries. For. Chron. 1960, 36, 10–13. [Google Scholar] [CrossRef]
- Peterson, R.L.; Wagg, C.; Pautler, M. Associations between microfungal endophytes and roots: Do structural features indicate function? Botany 2008, 86, 445–456. [Google Scholar] [CrossRef]
- Della Mónica, I.F.; Saparrat, M.C.; Godeas, A.M.; Scervino, J.M. The co-existence between DSE and AMF symbionts affects plant P pools through P mineralization and solubilization processes. Fungal Ecol. 2015, 17, 10–17. [Google Scholar] [CrossRef]
- Huang, W.-K.; Cui, J.-K.; Liu, S.-M.; Kong, L.-A.; Wu, Q.-S.; Peng, H.; He, W.-T.; Sun, J.-H.; Peng, D.-L. Testing various biocontrol agents against the root-knot nematode (Meloidogyne incognita) in cucumber plants identifies a combination of Syncephalastrum racemosum and Paecilomyces lilacinus as being most effective. Boil. Control. 2016, 92, 31–37. [Google Scholar] [CrossRef]
- Goettel, M.S.; Roberts, D.W. Mass production, formulation and field application of entomopathogenic fungi. In Biological Control of Locusts and Grasshopper; Lomer, C.J., Prior, C., Eds.; CAB International: Wallingford, UK, 1992. [Google Scholar]
- Lane, B.S.; Trinci, A.P.; Gillespie, A.T. Influence of cultural conditions on the virulence of conidia and blastospores of Beauveria bassiana to the green leafhopper, Nephotettix virescens. Mycol. Res. 1991, 95, 829–833. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Schuster, A.; Schmoll, M. Biology and biotechnology of Trichoderma. Appl. Microbiol. Biotechnol. 2010, 87, 787–799. [Google Scholar] [CrossRef] [Green Version]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial siderophores and their potential applications: A review. Environ. Sci. Pollut. Res. 2015, 23, 3984–3999. [Google Scholar] [CrossRef] [PubMed]
- Vala, A.K.; Vaidya, S.Y.; Dube, H.C. Siderophore production by facultative marine fungi. Indian J. Mar. Sci. 2000, 29, 339–340. [Google Scholar]
- Renshaw, J.C.; Robson, G.; Trinci, A.P.; Wiebe, M.; Livens, F.R.; Collison, D.; Taylor, R. Fungal siderophores: Structures, functions and applications. Mycol. Res. 2002, 106, 1123–1142. [Google Scholar] [CrossRef]
- Ivanova, E.G.; Doronina, N.V.; Trotsenko, Y.A. Aerobic Methylobacteria Are Capable of Synthesizing Auxins. Microbiology 2001, 70, 392–397. [Google Scholar] [CrossRef]
- Khalid, A.; Arshad, M.; Zahir, Z. Screening plant growth-promoting rhizobacteria for improving growth and yield of wheat. J. Appl. Microbiol. 2004, 96, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Kucharova, Z. Selection for root colonizing bacteria stimulating wheat growth in saline soils. Biol. Fertil. Soil 2009, 45, 563–571. [Google Scholar] [CrossRef]
- Ali, S. Endophytic fungi from Caralluma acutangula can secrete plant growth promoting enzymes. Fresenius Environ. Bull. 2019, 28, 2688–2696. [Google Scholar]
- Waqas, M.; Khan, A.L.; Shahzad, R.; Ullah, I.; Khan, A.R.; Lee, I.-J. Mutualistic fungal endophytes produce phytohormones and organic acids that promote japonica rice plant growth under prolonged heat stress*. J. Zhejiang Univ. Sci. B 2015, 16, 1011–1018. [Google Scholar] [CrossRef] [Green Version]
- Waqas, M.; Khan, A.L.; Lee, I.-J. Bioactive chemical constituents produced by endophytes and effects on rice plant growth. J. Plant Interact. 2013, 9, 478–487. [Google Scholar] [CrossRef]
- Kucey, R.M.N. Effect of Penicillium bilaji on the Solubility and Uptake of P and Micronutrients from Soil by Wheat. Can. J. Soil Sci. 1988, 68, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.S.; Zaidi, A.; Ahemad, M.; Oves, M.; Wani, P.A. Plant growth promotion by phosphate solubilizing fungi–current perspective. Arch. Agron. Soil Sci. 2010, 56, 73–98. [Google Scholar] [CrossRef]
- Bilal, S.; Shahzad, R.; Khan, A.L.; Kang, S.-M.; Imran, Q.M.; Al-Harrasi, A.; Yun, B.-W.; Lee, I.-J. Endophytic Microbial Consortia of Phytohormones-Producing Fungus Paecilomyces formosus LHL10 and Bacteria Sphingomonas sp. LK11 to Glycine max L. Regulates Physio-hormonal Changes to Attenuate Aluminum and Zinc Stresses. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Alikhani, H.A.; Hosseini, H.M. Indole-3-acetic acid (IAA) production trait, a useful screening to select endophytic and rhizosphere competent bacteria for rice growth promoting agents. MethodsX 2015, 2, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Mastouri, F.; Björkman, T.; Harman, G.E. Seed Treatment withTrichoderma harzianumAlleviates Biotic, Abiotic, and Physiological Stresses in Germinating Seeds and Seedlings. Phytopathology 2010, 100, 1213–1221. [Google Scholar] [CrossRef] [Green Version]
- Cochran, H.C. Effect of seed size on uniformity of pimento transplants (Capsicum annuum L.) at harvest time. J. Am. Sot. Hort. Sci. 1974, 99, 234–235. [Google Scholar]
- Waqas, M.; Khan, A.L.; Hamayun, M.; Kamran, M.; Kang, S.M.; Kim, Y.H.; Lee, I.J. Assessment of endophytic fungi cultural filtrate on soybean seed germination. Afr. J. Biotechnol. 2012, 11, 15135–15143. [Google Scholar]
- Jia, C.; Liu, Z.; Zhang, M.; Jia, J.; Zheng, L.; Wang, Q.; Wang, H. Effects of extracts from Paecilomyces variotii on the yield and quality of cherry radish (Raphanus sativus L. var. radculus pers). J. Agric. Res. Environ. 2019, 36, 176–183. [Google Scholar]
- Anis, M.; Abbasi, W.; Zaki, M.J. Bioefficacy of microbial antagonists against Macrophomina phaseolina on sunflower. Pak. J. Bot. 2010, 42, 2935–2940. [Google Scholar]
- Maitlo, S.A.; Rajput, N.A.; Syed, R.N.; Khanzada, M.A.; Rajput, A.Q.; Lodhi, A.M. Microbial control of Fusarium wilt of chickpea caused by Fusarium oxysporum f. sp. ciceris. Pak. J. Bot. 2019, 51, 2261–2268. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | |||||
|---|---|---|---|---|---|
| % (v/v) water | |||||
| 10 | 20 | 30 | |||
| BH-O% | CFU·g−1 | BH-O% | CFU g−1 | BH-O% | CFU g−1 |
| 90-10 | 1.13 × 108 ± 1.2 × 107 b | 90-10 | 8.5 × 107 ± 7.09 × 106 ef | 90-10 | 9.14 × 107 ± 9.73 × 106 de |
| 80-20 | 7.0 × 107 ± 1.32 × 107 g | 80-20 | 1.05 × 108 ± 1.09 × 107 bc | 80-20 | 1.0 × 108 ± 5.11 × 106 d |
| 70-30 | 8.04 × 107 ± 1.14 × 107 f | 70-30 | 1.24 × 108 ± 1.06 × 107 a | 70-30 | 1.07 × 108 ± 9.54 × 106 bc |
| Treatment | % Germination | Root Length (cm) | Shoot Length (cm) | Seed Vigor Index |
|---|---|---|---|---|
| Pepper | ||||
| p -value | 0.5369 | 0.6162 | 0.0010 | 0.9975 |
| P.variotii | 80 ± 9.79 | 2.37 ± 1.30 | 1.66 ± 0.55 | 323.00 ± 127.07 |
| T0 | 76 ± 7.30 | 2.26 ± 1.24 | 1.98 ± 0.56 | 322.93 ± 119.36 |
| Tomato | ||||
| p-value | 0.0020 | 0.0013 | 0.0036 | 0.0000 |
| P.variotii | 89 ± 3.82 | 4.41 ± 1.33 | 3.30 ± 1.15 | 686.70 ± 210.40 |
| T0 | 75 ± 3.82 | 3.73 ± 1.32 | 2.80 ± 1.02 | 498.70 ± 164.28 |
| Treatment | Length of Stem (cm) | Diameter (mm) | Number of Leaves | Areal Dry Weight (g) | Root Dry Weight (g) | Leaf Area mm2 | DQI |
|---|---|---|---|---|---|---|---|
| Pepper | |||||||
| p-value | 0.0000 | 0.0008 | 0.0000 | 0.0000 | 0.0022 | 0.0020 | 0.0054 |
| P. variotii | 29.63 ± 1.75 | 3.83 ± 0.21 | 7.39 ± 0.54 | 0.42 ± 0.05 | 0.13 ± 0.02 | 83.58 ± 11.27 | 0.051 ± 0.00 |
| T0 | 27.01 ± 2.07 | 3.65 ± 0.23 | 6.68 ± 0.70 | 0.36 ± 0.04 | 0.11 ± 0.03 | 75.88 ± 11.15 | 0.046 ± 0.00 |
| Tomato | |||||||
| p-value | 0.0002 | 0.0039 | 0.0330 | 0.0042 | 0.1443 | 0.0397 | 0.2059 |
| P. variotii | 27.37 ± 2.06 | 3.81 ± 0.27 | 4.40 ± 0.54 | 0.62 ± 0.07 | 0.14 ± 0.02 | 74.45 ± 11.85 | 0.067 ± 0.01 |
| T0 | 25.61 ± 2.07 | 3.78 ± 0.22 | 4.15 ± 0.48 | 0.56 ± 0.10 | 0.15 ± 0.02 | 69.28 ± 10.17 | 0.063 ± 0.01 |
| Tomato seedling | ||||||
| Treatment | Length of stem (cm) | Diameter (mm) | Nº leaves | Areal dry Weight (g) | Root dry Weight (g) | DQI |
| p-value | 0.0000 | 0.0000 | 0.0002 | 0.0000 | 0.0028 | 0.0481 |
| Pae D1 | 16.12 ± 2.11a | 3.72 ± 0.38b | 3.58 ± 0.53b | 0.27 ± 0.09a | 0.039 ± 0.01a | 0.045 ± 0.01b |
| Pae D2 | 14.46 ± 2.58b | 3.50 ± 0.38c | 3.71 ± 0.84ab | 0.25 ± 0.08a | 0.035 ± 0.01a | 0.046 ± 0.01b |
| Pae D3 | 16.27 ± 3.05a | 3.99 ± 0.28a | 3.93 ± 0.78a | 0.38 ± 0.09a | 0.027 ± 0.01b | 0.054 ± 0.01a |
| T0 | 12.5 ± 2.22b | 3.45 ± 0.36c | 3.29 ± 0.61c | 0.25 ± 0.08b | 0.036 ± 0.01a | 0.045 ± 0.01b |
| Tomato Plants | ||||||
| Treatment | Length of stem (cm) | Diameter (mm) | Internodes | Areal dry Weight (g) | Root dry Weight (g) | DQI |
| p-value | 0.1876 | 0,1244 | 0.9759 | 0.0048 | 0.7796 | 0.0283 |
| Pae D1 | 82.75 ± 12.97ab | 11.57 ± 1.08ab | 14.20 ± 1.93a | 50.67 ± 10.80a | 4.22 ± 1.20a | 24.84 ± 7.68a |
| Pae D2 | 87.20 ± 9.42a | 11.59 ± 1.02ab | 14.30 ± 1.25a | 49.19 ± 10.21a | 4.26 ± 1.64a | 24.23 ± 8.16a |
| Pae D3 | 77.60 ± 7.75b | 11.84 ± 1.08a | 14.10 ± 1.28a | 49.38 ± 10.53a | 4.20 ± 1.37a | 25.78 ± 7.66a |
| T0 | 84.00 ± 7.10ab | 10.58 ± 0.91b | 14.00 ± 1.63a | 39.13 ± 7.99b | 3.73 ± 0.91a | 16.84 ± 3.79b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Gavíra, A.; Diánez, F.; Sánchez-Montesinos, B.; Santos, M. Paecilomyces variotii as A Plant-Growth Promoter in Horticulture. Agronomy 2020, 10, 597. https://doi.org/10.3390/agronomy10040597
Moreno-Gavíra A, Diánez F, Sánchez-Montesinos B, Santos M. Paecilomyces variotii as A Plant-Growth Promoter in Horticulture. Agronomy. 2020; 10(4):597. https://doi.org/10.3390/agronomy10040597
Chicago/Turabian StyleMoreno-Gavíra, Alejandro, Fernando Diánez, Brenda Sánchez-Montesinos, and Mila Santos. 2020. "Paecilomyces variotii as A Plant-Growth Promoter in Horticulture" Agronomy 10, no. 4: 597. https://doi.org/10.3390/agronomy10040597
APA StyleMoreno-Gavíra, A., Diánez, F., Sánchez-Montesinos, B., & Santos, M. (2020). Paecilomyces variotii as A Plant-Growth Promoter in Horticulture. Agronomy, 10(4), 597. https://doi.org/10.3390/agronomy10040597