Assessing the Impact of Drought Stress and Soil Cultivation in Chardonnay and Xynisteri Grape Cultivars


 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Conditions and Cultivars
2.2. Vegetative Soil Coverage
2.3. Plant Growth, Production and Physiological Parameters
2.4. Total Phenols and Antioxidant Activity in Leaves
2.5. Plant Nutrient Content
2.6. Phytohormones in Leaves
2.7. Berries Qualitative Attributes
2.8. Statistical Analysis
3. Results
3.1. Vineyards Soil Properties and Water Content
3.2. Biomass Production from Cover Crop
3.3. Plant Growth and Physiology
3.4. Polyphenols and Antioxidant Activity
3.5. Mineral Content in Leaf Stems
3.6. Phytohormones in Leaves
3.7. Grape Quality Attributes
3.8. Overall Effect
4. Discussion
4.1. Vineyards Soil Properties and Soil Water Content
4.2. Biomass Production from Cover Crop
4.3. Plant Growth and Physiology
4.4. Polyphenols and Antioxidant Activity
4.5. Mineral Content in Leaf Stems and Phytohormones in Leaves
4.6. Quality Attributes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Min, Z.; Li, R.; Chen, L.; Zhang, Y.; Li, Z.; Liu, M.; Ju, Y.; Fang, Y. Alleviation of drought stress in grapevine by foliar-applied strigolactones. Plant Physiol. Biochem. 2019, 135, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.G.; Sutherland, R.A. Stomatal control of xylem embolism. Plant Cell Environ. 1991, 11, 111–121. [Google Scholar] [CrossRef]
- Chen, L.; Jun, A.I.; Wang, Z.X.; Zhao, Y. Research progress on effect of drought stress on the physiological property and microstructure in grapevine. North. Hortic. 2011, 6, 205–209. [Google Scholar]
- Patakas, A.; Noitsakis, B.; Chouzouri, A. Optimization of irrigation water use in grapevines using the relationship between transpiration and plant water status. Agric. Ecosyst. Environ. 2005, 106, 253–259. [Google Scholar] [CrossRef]
- Kofidis, G.; Bozabalidis, A.; Chartzoulakis, K. Leaf anatomical alterations induced by drought stress in two avocado cultivars. J. Biol. Res. 2004, 1, 115–120. [Google Scholar]
- Ferrandino, A.; Ferrandino, A.; Perrone, I.; Tramontini, S. Physiological and molecular mechanisms of resistance to drought in Vitis vinifera L: Aspects of primary and secondary metabolisms and adaptation of genotypes. Italus Hortus 2009, 16, 23–34. [Google Scholar]
- Salazar-Parra, C.; Aguirreolea, J.; Sánchez-Díaz, M.; Irigoyen, J.J.; Morales, F. Climate change (elevated CO2, elevated temperature and moderate drought) triggers the antioxidant enzymes’ response of grapevine cv. Tempranillo, avoiding oxidative damage. Physiol. Plant. 2012, 144, 99–110. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Chrysargyris, A.; Aziz, A. Adaptive response of a native mediterranean grapevine cultivar upon short-term exposure to drought and heat stress in the context of climate change. Agronomy 2020, 10, 249. [Google Scholar] [CrossRef] [Green Version]
- Kyraleou, M.; Koundouras, S.; Kallithraka, S.; Theodorou, N.; Proxenia, N.; Kotseridis, Y. Effect of irrigation regime on anthocyanin content and antioxidant activity of Vitis vinifera L. cv. Syrah grapes under semiarid conditions. J. Sci. Food Agric. 2016, 96, 988–996. [Google Scholar] [CrossRef]
- Acevedo-Opazo, C.; Ortega-Farias, S.; Fuentes, S. Effects of grapevine (Vitis vinifera L.) water status on water consumption, vegetative growth and grape quality: An irrigation scheduling application to achieve regulated deficit irrigation. Agric. Water Manag. 2010, 97, 956–964. [Google Scholar] [CrossRef]
- Romero, P.; Fernández-Fernández, J.I.; Martinez-Cutillas, A. Physiological thresholds for efficient Regulated Deficit Irrigation management in winegrapes under semiarid conditions: Soil-plant-water relationships and berry composition. Acta Hortic. 2012, 931, 171–178. [Google Scholar] [CrossRef]
- Intrigliolo, D.S.; Castel, J.R. Interactive effects of deficit irrigation and shoot and cluster thinning on grapevine cv. Tempranillo. Water relations, vine performance and berry and wine composition. Irrig. Sci. 2011, 29, 443–454. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2016, 6, 166–171. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.V.; White, M.A.; Cooper, O.R.; Storchmann, K. Climate change and global wine quality. Clim. Chang. 2005, 73, 319–343. [Google Scholar] [CrossRef]
- Beis, A.; Patakas, A. Differential physiological and biochemical responses to drought in grapevines subjected to partial root drying and deficit irrigation. Eur. J. Agron. 2015, 62, 90–97. [Google Scholar] [CrossRef]
- Dagianta, E.; Goumas, D.; Manios, T.; Tzortzakis, N. The use of treated wastewater and fertigation in greenhouse pepper crop as affecting growth and fruit quality. J. Water Reuse Desalin. 2014, 4, 92–99. [Google Scholar] [CrossRef]
- Steenwerth, K.L.; Calderón-Orellana, A.; Hanifin, R.C.; Storm, C.; McElrone, A.J. Effects of various vineyard floor management techniques on weed community shifts and grapevine water relations. Am. J. Enol. Vitic. 2016, 67, 153–162. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Xylia, P.; Litskas, V.; Mandoulaki, A.; Antoniou, D.; Boyias, T.; Stavrinides, M.; Tzortzakis, N. Drought stress and soil management practices in grapevines in Cyprus under the threat of climate change. J. Water Clim. Chang. 2018, 9, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Chrysargyris, A.; Xylia, P.; Antoniou, O.; Tzortzakis, N. Climate change due to heat and drought stress can alter the physiology of maratheftiko local cyprian grapevine variety. J. Water Clim. Chang. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Pellegrino, A.; Lebon, E.; Simonneau, T.; Wery, J. Towards a simple indicator of water stress in grapevine (Vitis vinifera L.) based on the differential sensitivities of vegetative growth components. Aust. J. Grape Wine Res. 2005, 11, 306–315. [Google Scholar] [CrossRef]
- Bahar, E.; Yaşain, A.S. The yield and berry quality under different soil tillage and clusters thinning treatments in grape (Vitis vinifera L.) cv. Cabernet-Sauvignon. Afr. J. Agric. Res. 2010, 5, 2986–2993. [Google Scholar]
- Montgomery, D.R. Soil erosion and agricultural sustainability. Proc. Natl. Acad. Sci. USA 2007, 104, 13268–13272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, J.B.; Southard, R.J.; Mitchell, J.P. Agricultural dust production in standard and conservation tillage systems in the san joaquin valley. J. Environ. Qual. 2005, 34, 1260–1269. [Google Scholar] [CrossRef] [Green Version]
- Wolff, M.W.; Alsina, M.M.; Stockert, C.M.; Khalsa, S.D.S.; Smart, D.R. Minimum tillage of a cover crop lowers net GWP and sequesters soil carbon in a California vineyard. Soil Tillage Res. 2018, 175, 244–254. [Google Scholar] [CrossRef]
- Daane, K.M.; Hogg, B.N.; Wilson, H.; Yokota, G.Y. Native grass ground covers provide multiple ecosystem services in Californian vineyards. J. Appl. Ecol. 2018, 55, 2473–2483. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Bermúdez, P.; Olmo, M.; Gil, J.; García-Férriz, L.; Olmo, C.; Boluda, R.; Gavidia, I. Cover crops and pruning in Bobal and Tempranillo vineyards have little influence on grapevine nutrition. Sci. Agric. 2016, 73, 260–265. [Google Scholar] [CrossRef] [Green Version]
- Keller, M. From the ASEV soil environment and vine mineral nutrition symposium: Deficit irrigation and vine mineral nutrition. Am. J. Enol. Vitic. 2005, 56, 267–283. [Google Scholar]
- Tombesi, S.; Nardini, A.; Frioni, T.; Soccolini, M.; Zadra, C.; Farinelli, D.; Poni, S.; Palliotti, A. Stomatal closure is induced by hydraulic signals and maintained by ABA in drought-stressed grapevine. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef]
- Davies, W.J.; Kudoyarova, G.; Hartung, W. Long-distance ABA signaling and its relation to other signaling pathways in the detection of soil drying and the mediation of the plant’s response to drought. J. Plant Growth Regul. 2005, 24, 285–295. [Google Scholar] [CrossRef] [Green Version]
- McAdam, S.A.M.; Brodribb, T.J. Hormonal dynamics contributes to divergence in seasonal stomatal behaviour in a monsoonal plant community. Plant Cell Environ. 2015, 38, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N.; Yadav, G.; Singh, J.; Mishra, R.K.; Kumar, V.; Verma, R.; Upadhyay, R.G.; Pandey, M.; et al. Abscisic acid signaling and abiotic stress tolerance in plants: A review on current knowledge and future prospects. Front. Plant Sci. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.F.; Xu, T.F.; Wang, Z.Z.; Fang, Y.L.; Xi, Z.M.; Zhang, Z.W. The ameliorative effects of exogenous melatonin on grape cuttings under water-deficient stress: Antioxidant metabolites, leaf anatomy, and chloroplast morphology. J. Pineal Res. 2014, 57, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zheng, P.; Meng, J.; Xi, Z. Effect of exogenous 24-epibrassinolide on chlorophyll fluorescence, leaf surface morphology and cellular ultrastructure of grape seedlings (Vitis vinifera L.) under water stress. Acta Physiol. Plant. 2015, 37, 1729. [Google Scholar] [CrossRef]
- Litskas, V.D.; Irakleous, T.; Tzortzakis, N.; Stavrinides, M.C. Determining the carbon footprint of indigenous and introduced grape varieties through Life Cycle Assessment using the island of Cyprus as a case study. J. Clean. Prod. 2017, 156, 418–425. [Google Scholar] [CrossRef]
- Hatmi, S.; Gruau, C.; Trotel-Aziz, P.; Villaume, S.; Rabenoelina, F.; Baillieul, F.; Eullaffroy, P.; Clément, C.; Ferchichi, A.; Aziz, A. Drought stress tolerance in grapevine involves activation of polyamine oxidation contributing to improved immune response and low susceptibility to Botrytis cinerea. J. Exp. Bot. 2015, 66, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Chrysargyris, A.; Drouza, C.; Tzortzakis, N. Optimization of potassium fertilization/nutrition for growth, physiological development, essential oil composition and antioxidant activity of Lavandula angustifolia Mill. J. Soil Sci. Plant Nutr. 2017, 17, 291–306. [Google Scholar] [CrossRef] [Green Version]
- Klados, E.; Tzortzakis, N. Effects of substrate and salinity in hydroponically grown Cichorium spinosum. J. Soil Sci. Plant Nutr. 2014, 14, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Chrysargyris, A.; Panayiotou, C.; Tzortzakis, N. Nitrogen and phosphorus levels affected plant growth, essential oil composition and antioxidant status of lavender plant (Lavandula angustifolia Mill.). Ind. Crops Prod. 2016, 83, 577–586. [Google Scholar] [CrossRef]
- Xylia, P.; Chrysargyris, A.; Botsaris, G.; Tzortzakis, N. Potential application of spearmint and lavender essential oils for assuring endive quality and safety. Crop Prot. 2017, 102, 94–103. [Google Scholar] [CrossRef]
- Marinou, E.; Chrysargyris, A.; Tzortzakis, N. Use of sawdust, coco soil and pumice in hydroponically grown strawberry. Plant Soil Environ. 2013, 59, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Haeck, A.; Van Langenhove, H.; Harinck, L.; Kyndt, T.; Gheysen, G.; Höfte, M.; Demeestere, K. Trace analysis of multi-class phytohormones in Oryza sativa using different scan modes in high-resolution Orbitrap mass spectrometry: Method validation, concentration levels, and screening in multiple accessions. Anal. Bioanal. Chem. 2018, 410, 4527–4539. [Google Scholar] [CrossRef] [PubMed]
- OIV. Compendium of International Methods of Analysis of Wines and Musts; International Organisation of Vine and Wine: Paris, France, 2012. [Google Scholar]
- Chrysargyris, A.; Nikou, A.; Tzortzakis, N. Effectiveness of Aloe vera gel coating for maintaining tomato fruit quality. N. Z. J. Crop Hortic. Sci. 2016, 44, 203–217. [Google Scholar] [CrossRef]
- Deng, Y.; Wu, Y.; Li, Y. Effects of high O2 levels on post-harvest quality and shelf life of table grapes during long-term storage. Eur. Food Res. Technol. 2005, 221, 392–397. [Google Scholar] [CrossRef]
- Du, B.; He, B.-J.; Shi, P.-B.; Li, F.-Y.; Li, J.; Zhu, F. Phenolic content and antioxidant activity of wine grapes and table grapes. J. Med. Plants Res. 2012, 6, 3381–3387. [Google Scholar]
- Bate-smith, E.C. Astringent tannins of the leaves of Geranium species. Phytochemistry 1981, 20, 211–216. [Google Scholar] [CrossRef]
- Simonsson, M.; Andersson, S.; Andrist-Rangel, Y.; Hillier, S.; Mattsson, L.; Öborn, I. Potassium release and fixation as a function of fertilizer application rate and soil parent material. Geoderma 2007, 140, 188–198. [Google Scholar] [CrossRef]
- Marschner, P.; Rengel, Z. Nutrient Availability in Soils; Elsevier Ltd.: Amsterdam, The Netherlands, 2011; ISBN 9780123849052. [Google Scholar]
- Wichelns, D.; Qadir, M. Achieving sustainable irrigation requires effective management of salts, soil salinity, and shallow groundwater. Agric. Water Manag. 2015, 157, 31–38. [Google Scholar] [CrossRef]
- Gómez, J.A.; Llewellyn, C.; Basch, G.; Sutton, P.B.; Dyson, J.S.; Jones, C.A. The effects of cover crops and conventional tillage on soil and runoff loss in vineyards and olive groves in several Mediterranean countries. Soil Use Manag. 2011, 27, 502–514. [Google Scholar] [CrossRef] [Green Version]
- Soriano, M.A.; Cabezas, J.M.; Ramos, A.; Lora, A.; Gómez, J.A. Characterization of cover crops for use in olive groves and vineyards in certified systems under mediterranean conditions. In Proceedings of the 10th International Symposium on Agriculture and the Environment, Purdue University, West Lafayette, IN, USA, 23–27 May 2016. [Google Scholar]
- Blanco-Canqui, H.; Wienhold, B.J.; Jin, V.L.; Schmer, M.R.; Kibet, L.C. Long-term tillage impact on soil hydraulic properties. Soil Tillage Res. 2017, 170, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Costa, J.M.; Zarrouk, O.; Pinheiro, C.; Lopes, C.M.; Pereira, J.S. Controlling stomatal aperture in semi-arid regions—The dilemma of saving water or being cool? Plant Sci. 2016, 251, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Casas, M.; Pajaro, M.; Lores, M.; Garcia-Jares, C. Polyphenolic composition and antioxidant activity of galician monovarietal wines from native and experimental non-native white grape varieties. Int. J. Food Prop. 2016, 19, 2307–2321. [Google Scholar] [CrossRef]
- Olejar, K.J.; Fedrizzi, B.; Kilmartin, P.A. Enhancement of Chardonnay antioxidant activity and sensory perception through maceration technique. LWT Food Sci. Technol. 2016, 65, 152–157. [Google Scholar] [CrossRef]
- Garrido, T.; Gizdavic-Nikolaidis, M.; Leceta, I.; Urdanpilleta, M.; Guerrero, P.; de la Caba, K.; Kilmartin, P.A. Optimizing the extraction process of natural antioxidants from chardonnay grape marc using microwave-assisted extraction. Waste Manag. 2019, 88, 110–117. [Google Scholar] [CrossRef]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Ali, E.; Fahad, S. Phytohormones enhanced drought tolerance in plants: A coping strategy. Environ. Sci. Pollut. Res. 2018, 25, 33103–33118. [Google Scholar] [CrossRef]
- Price, S.F.; Breen, P.J.; Valladao, M.; Warson, B.T. Cluster sun exposure and quercetin in Pinot noir grapes and wine. Am. J. Enol. Vitic. 1995, 46, 187–194. [Google Scholar]
- Tregoat, O.; Van Leeuwen, C.; Choné, X.; Gaudillère, J.P. Étude du régime hydrique et de la nutrition azotée de la vigne par des indicateurs physiologiques influence sur le comportement de la vigne et la maturation du raisin (vitis vinifera L. cv merlot, 2000, bordeaux). J. Int. Sci. Vigne Vin 2002, 36, 133–142. [Google Scholar]
- Ojeda, H.; Andary, C.; Kraeva, E.; Carbonneau, A.; Deloire, A. Influence of pre- and postveraison water deficit on synthesis and concentration of skin phenolic compounds during berry growth of Vitis vinifera cv. Shiraz. Am. J. Enol. Vitic. 2002, 53, 261–267. [Google Scholar]
- Barroso, J.M.; Pombeiro, L.; Rato, A.E. Impacts of crop level, soil and irrigation management in grape berries of cv ‘Trincadeira’ (Vitis vinifera L.). J. Wine Res. 2017, 28, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Deis, L.; Cavagnaro, J.B. Effect of water stress in grape berries cabernet sauvignon (mendoza, argentina) during four years consecutives. J. Life Sci. 2013, 7, 993–1001. [Google Scholar]
- Bucchetti, B.; Matthews, M.A.; Falginella, L.; Peterlunger, E.; Castellarin, S.D. Effect of water deficit on Merlot grape tannins and anthocyanins across four seasons. Sci. Hortic. 2011, 128, 297–305. [Google Scholar] [CrossRef]
- Santesteban, L.G.; Miranda, C.; Royo, J.B. Regulated deficit irrigation effects on growth, yield, grape quality and individual anthocyanin composition in Vitis vinifera L. cv. “Tempranillo.”. Agric. Water Manag. 2011, 98, 1171–1179. [Google Scholar] [CrossRef]
- Koundouras, S.; Marinos, V.; Gkoulioti, A.; Kotseridis, Y.; Van Leeuwen, C. Influence of vineyard location and vine water status on fruit maturation of nonirrigated cv. Agiorgitiko (Vitis vinifera L.). Effects on wine phenolic and aroma components. J. Agric. Food Chem. 2006, 54, 5077–5086. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chardonnay | Xynisteri | |||||||
|---|---|---|---|---|---|---|---|---|
| Plant Parts/Production | Penological Stage | Parameters | Tillage (T) | Irrigation (I) | T × I | Tillage (T) | Irrigation (I) | T × I |
| Leaves | Veraison | Leaf Fluorescence (Fv/Fm) | ns | ns | ns | ns | ns | ns |
| Veraison | Stomatal conductance (mmol m−2 s−1) | * | ns | * | ns | ** | ns | |
| Veraison | Chl a (mg g−1 Fw) | ns | * | *** | ns | ns | ns | |
| Veraison | Chl b (mg g−1 Fw) | ** | ** | *** | ns | ns | ns | |
| Veraison | Total Chl (mg g−1 Fw) | ns | ** | *** | ns | ns | ns | |
| Veraison | Total Phenols (mg GA g−1 Fw) | ns | ns | ns | * | ns | ns | |
| Veraison | DPPH (mg Trolox g−1 Fw) | ns | ns | ns | ns | ** | ns | |
| Veraison | FRAP (mg Trolox g−1 Fw) | ns | ns | ns | ns | ns | ns | |
| Veraison | N (g kg−1 Dw) | ns | ** | * | ns | ns | ns | |
| Veraison | K (g kg−1 Dw) | *** | *** | ns | * | * | ns | |
| Veraison | P (g kg−1 Dw) | *** | *** | *** | *** | ns | *** | |
| Veraison | Mg (g kg−1 Dw) | ns | *** | ns | *** | ** | * | |
| Veraison | Na (g kg−1 Dw) | *** | *** | *** | ns | ** | *** | |
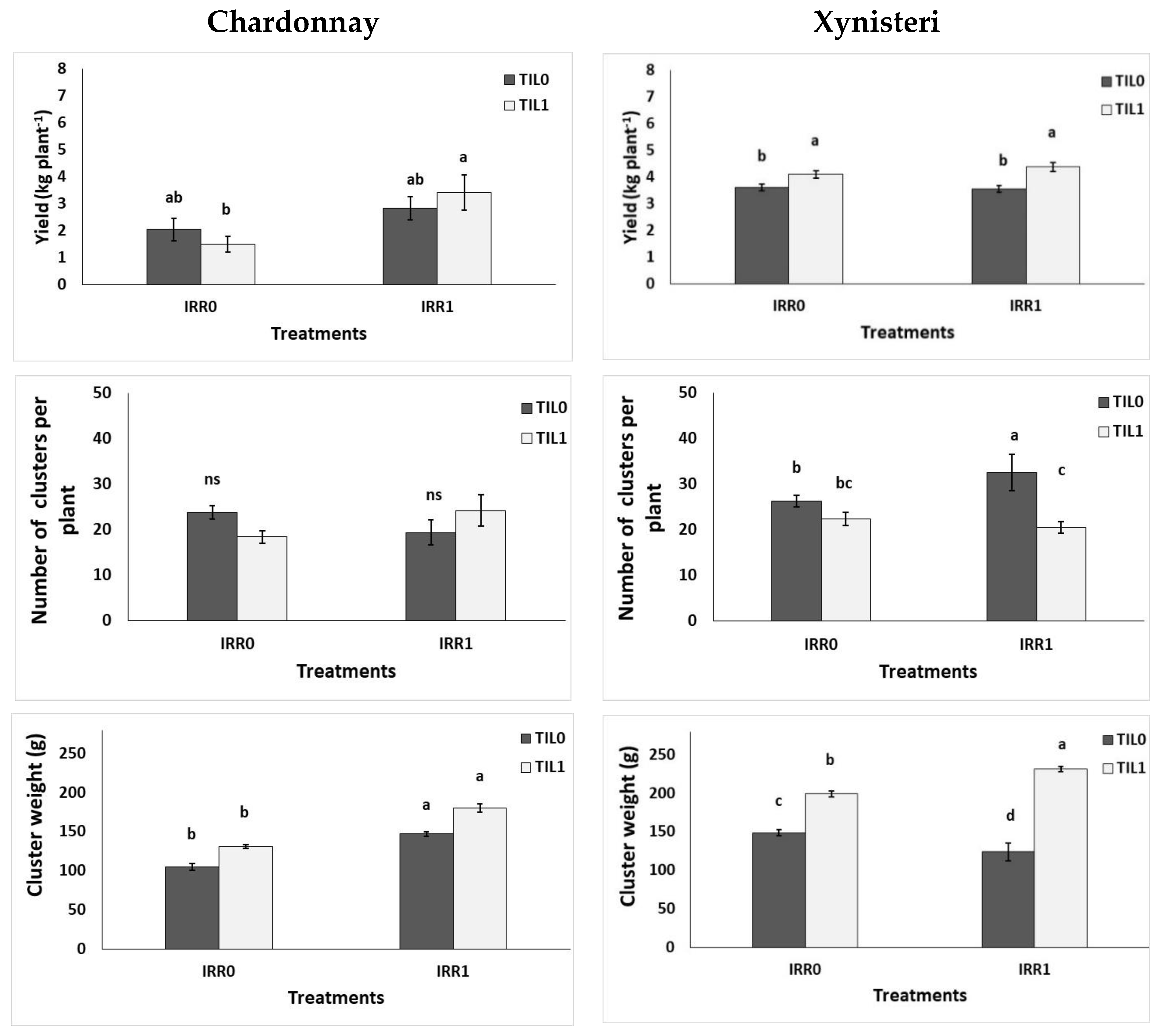
| Production | Harvesting | No of clusters per plant | ns | ns | ns | ** | ns | * |
| Harvesting | Yield (kg plant−1) | ns | ns | ns | ** | ns | ns | |
| Harvesting | Cluster fresh weight (g) | ns | ns | ns | *** | ns | * | |
| Grapes | Harvesting | Total soluble solids (oBrix) | * | ns | ns | ns | ns | ns |
| Harvesting | pH | * | * | ns | ns | ns | ns | |
| Harvesting | Titratable acidity (% tartaric acid) | *** | ** | ns | ns | * | ns | |
| Harvesting | Ascorbic acid (mg 100 mL−1 grape juice) | ns | ns | ns | ns | ns | ns | |
| Harvesting | Total phenols (mg GA 100 g−1 Fw) | *** | ** | ns | *** | *** | *** | |
| Harvesting | Anthocyanins (mg cyn-3-glu 100 g−1 Fw) | ns | ns | ns | ns | ns | ns | |
| Harvesting | Tannins (mg 100 mL−1 grape juice) | ns | ns | ns | ns | ns | ns | |
| TSS | TA | pH | AA | Phenols | Anthocyanins | Tannins | ||
|---|---|---|---|---|---|---|---|---|
| No irrigation | No tillage | 19.50 ± 0.34b | 0.97 ± 0.05a | 2.86 ± 0.04b | 0.58 ± 0.04a | 3.69 ± 0.16b | 0.25 ± 0.08a | 54.12 ± 8.86a |
| Tillage | 21.30 ± 0.54a | 0.71 ± 0.02b | 3.24 ± 0.08a | 0.84 ± 0.05a | 4.81 ± 0.18a | 0.28 ± 0.07a | 31.90 ± 4.89b | |
| Irrigation | No Tillage | 19.77 ± 0.52b | 0.81 ± 0.05b | 3.25 ± 0.08a | 0.81 ± 0.18a | 2.66 ± 0.11c | 0.19 ± 0.08a | 41.07 ± 5.87b |
| Tillage | 21.62 ± 0.28a | 0.62 ± 0.02c | 3.35 ± 0.00a | 0.77 ± 0.05a | 4.45 ± 0.12a | 0.42 ± 0.14a | 37.22 ± 7.96b |
| TSS | TA | pH | AA | Phenols | Anthocyanins | Tannins | ||
|---|---|---|---|---|---|---|---|---|
| No irrigation | No tillage | 17.90 ± 0.46a | 0.63 ± 0.02b | 3.40 ± 0.01a | 0.61 ± 0.04b | 2.65 ± 0.10b | 0.43 ± 0.16a | 58.00 ± 8.01a |
| Tillage | 19.82 ± 0.31a | 0.59 ± 0.01b | 3.35 ± 0.04ab | 0.73 ± 0.03a | 2.85 ± 0.09b | 0.42 ± 0.14a | 25.62 ± 6.74b | |
| Irrigation | No Tillage | 16.55 ± 1.71a | 0.66 ± 0.01ab | 3.31 ± 0.01b | 0.70 ± 0.03ab | 5.11 ± 0.13a | 0.40 ± 0.17a | 46.40 ± 4.78ab |
| Tillage | 17.42 ± 0.31a | 0.70 ± 0.03a | 3.31 ± 0.00b | 0.72 ± 0.03ab | 2.80 ± 0.06b | 0.54 ± 0.12a | 48.35 ± 9.56ab |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrysargyris, A.; Xylia, P.; Litskas, V.; Stavrinides, M.; Heyman, L.; Demeestere, K.; Höfte, M.; Tzortzakis, N. Assessing the Impact of Drought Stress and Soil Cultivation in Chardonnay and Xynisteri Grape Cultivars. Agronomy 2020, 10, 670. https://doi.org/10.3390/agronomy10050670
Chrysargyris A, Xylia P, Litskas V, Stavrinides M, Heyman L, Demeestere K, Höfte M, Tzortzakis N. Assessing the Impact of Drought Stress and Soil Cultivation in Chardonnay and Xynisteri Grape Cultivars. Agronomy. 2020; 10(5):670. https://doi.org/10.3390/agronomy10050670
Chicago/Turabian StyleChrysargyris, Antonios, Panayiota Xylia, Vassilis Litskas, Menelaos Stavrinides, Lisa Heyman, Kristof Demeestere, Monica Höfte, and Nikolaos Tzortzakis. 2020. "Assessing the Impact of Drought Stress and Soil Cultivation in Chardonnay and Xynisteri Grape Cultivars" Agronomy 10, no. 5: 670. https://doi.org/10.3390/agronomy10050670
APA StyleChrysargyris, A., Xylia, P., Litskas, V., Stavrinides, M., Heyman, L., Demeestere, K., Höfte, M., & Tzortzakis, N. (2020). Assessing the Impact of Drought Stress and Soil Cultivation in Chardonnay and Xynisteri Grape Cultivars. Agronomy, 10(5), 670. https://doi.org/10.3390/agronomy10050670