1. Introduction
Soil contains a large and variable quantity of potentially pathogenic microorganisms, such as fungi, bacteria and viruses that interact with seeds and seedlings. Maize (
Zea mays L.) seeds and seedlings are susceptible to infection from a number of fungal pathogens. This leads to the emergence of a small number of plants and, thus, heavy potential yield losses, for crops characterized by precision sowing and when there is not the possibility of self-regulating the plant population by means of tillering [
1]. Moreover, the plant population may be reduced to such an extent that replanting is necessary. Numerous soil fungi are associated with maize seedling diseases, such as
Fusarium, Pythium, Rhizoctonia and
Phytophthora [
2]. Among these, the
Fusarium genus is the most widespread fungus that affects maize in temperate areas. Several
Fusarium species can infect the seeds, seedlings and plants (root, stem) of maize and this can result in pre- or post-emergence damping-off [
3,
4]. The most common
Fusarium species isolated from maize crops in temperate maize growing areas are
F. verticillioides (Sacc.) Nirenberg, and
F. graminearum Schwabe. These fungal pathogens can survive in the soil or on crop debris [
5] and they affect the seeds and seedlings after planting germination (soil-borne infection). Moreover, seed inoculation with
Fusarium spp. (seed-borne infection) may contribute to the stand losses caused by damping-off, particularly if seeds are produced in growing areas prone to
Fusarium ear rot during ripening [
6]. Previous studies that were conducted to evaluate the contribution of
F. verticillioides seed-borne inoculum to maize seedling blight epidemics reported conflicting results [
6,
7,
8]. However, there is a general agreement on the negative effect of seed-borne
Fusarium spp. on germination and on a reduced seedling growth [
3,
9], while no field studies have highlighted the subsequent influence on plant growth or development considering the delay of anthesis and kernel maturity.
In North Italy, maize, with 800,000 ha located in the Po plain, is the most important crop, playing a key role in supporting agri-food supply chains. Prevention in the field is generally ineffective in reducing seed-borne infection, although any crop practice that favors a rapid germination and seedling growth can help to minimize its contribution to damping-off. However, only a few agronomic solutions are effective in reducing
Fusarium soil-borne infection. Of these solutions, crop rotation or burying debris may be able to reduce the potential of soil inoculum [
10]. Furthermore, in North Italy, as in several other temperate growing areas, the agronomic tendency is to anticipate the sowing time to such times when the soil temperature is above 10 °C in order to reduce water stress and injuries from insects and disease during ripening [
11], which leads to a positive effect on grain yields. This practice, in addition to the application of strip tillage or other minimum tillage practices on cereal farms, has led to more critical conditions for seedling mortality and plant development due to the longer germination. A high plant density is required to fully benefit from the yield potential of modern hybrids [
12], and the possible reduction in plant population after sowing led to clear yield gap. Thus, direct control solutions are necessary to minimize the risk associated with seedling mortality and the influence of fungal infection on plant growth and development. The use of chemicals is considered the best option for this purpose. As a result of the low cost and the specific action of fungicide seed treatments, they are considered an excellent solution to reduce early seed attacks from soil pathogens and to ensure emergence, even under critical environmental conditions [
13]. Moreover, fungicide seed dressings may contribute to minimizing the damping-off and growth delay related to seed-borne infection [
14]. Over the last two decades, fungicides from the phthalimide chemical family (e.g., captan) and dithiocarbamate (e.g., thiram) have progressively been substituted by phenylpyrroles (e.g., fludioxonil) phenylamides (e.g., metalaxyl, metalaxyl-M) and benzimidazoles (e.g., thiabendazole, carbendazim) [
15]. A two-way fungicide combination (two active ingredients for a specific pathogen target) that is widely applied for maize seed dressing is fludioxonil and metalaxyl-M, the former of which shows good activity against
Fusarium and
Rhizoctonia spp. [
16], while the latter is more effective on
Pythium and
Phytophthora [
17,
18]. Both of the previously reported compounds are non-systemic fungicides. Moreover, some of the fungicide formulations that have recently been proposed on the market are often mixtures of several active ingredients, which have different modes of action, in order to increase the control of a wide spectrum of pathogens. Strobilurins (e.g., azoxystrobin, pyraclostrobin and trifloxystrobin), triazoles (e.g., difeconazole, tebuconazole and prothioconazole) and pyrazole carboxamide (e.g., sedaxane) families, which are all characterized by a systemic activity, are some of the recent active ingredients that have been used for maize seed dressings.
Most of the studies conducted to evaluate the effect of fungicide seed treatments on
Fusarium species have been performed in vitro, in growth chambers or in greenhouses to investigate the plant–fungus pathosystem in the first growth stages [
3,
19], while only a few experiments have taken into account the complete crop cycle under field conditions. An even smaller number have investigated the effects of fungicide seed treatments on plant growth and development until harvest [
20], and in particular on grain yield [
18,
21]. Moreover, no information is available on the different effects of seed infection by
F. graminearum and
F. verticillioides from planting to harvest. The aim of the study has been to evaluate the role of fungicides applied to maize seeds in order to minimize the damping-off caused by seed-borne
Fusarium infection and to enhance the early vigor of plants and grain yield under different production conditions.
4. Discussion
The data collected from a large number of field studies have clearly shown the advantages of fungicide seed treatments on controlling seed-borne
F. verticillioides and
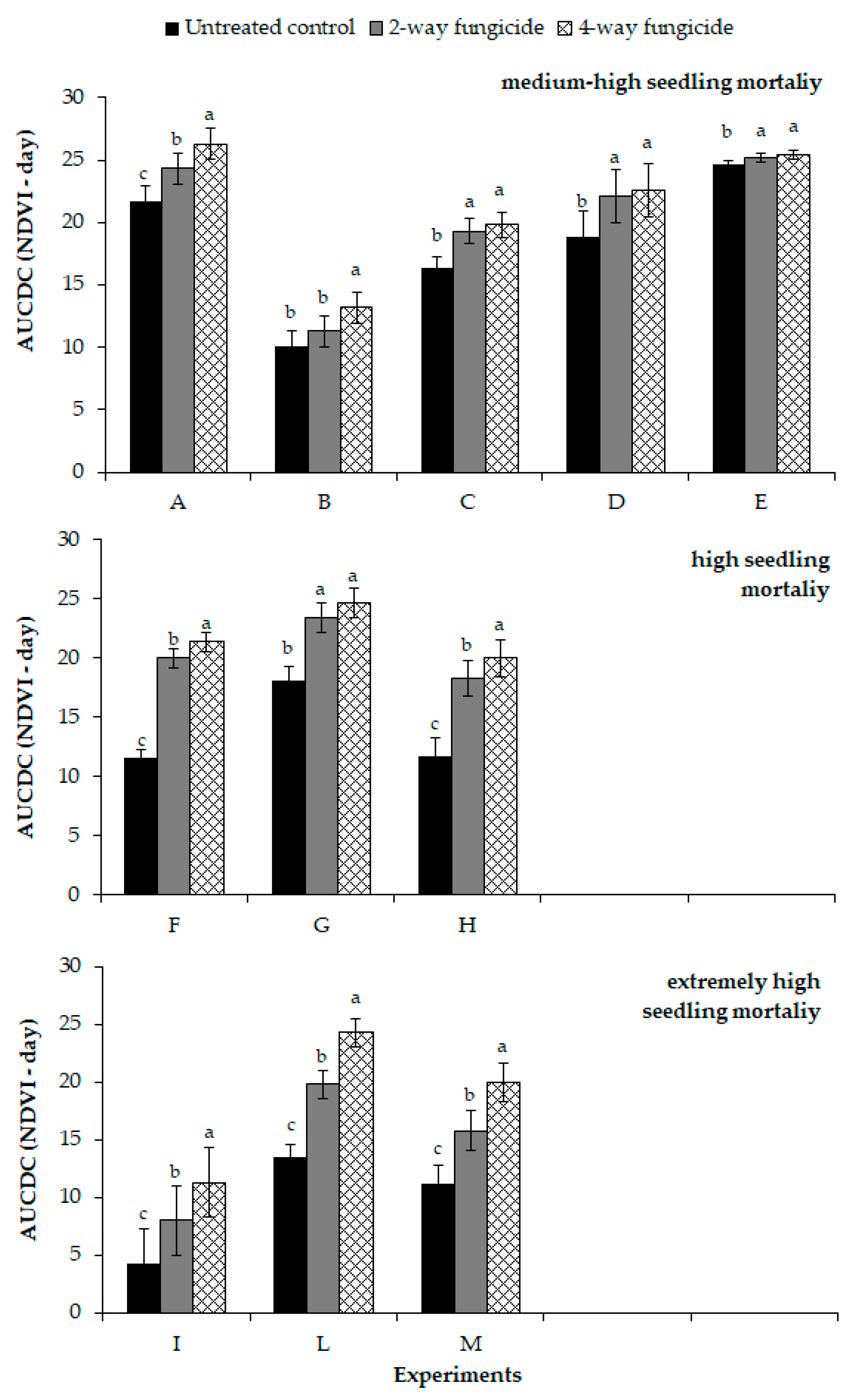
F. graminearum, in terms of both maize emergency and vigor (speed of growth), as well as on reducing and, in some cases, totally eliminating, the productive losses caused by fungi. Eleven experiments were conducted from 2015 to 2019 in different production situations (soil, meteorological trend, agronomic techniques), which have clearly influenced the negative impact of fungal infection on the percentage of emerged plants. The experiments were grouped into three clusters on the basis of the seedling mortality at emergence (medium high, high and extremely high): as expected, the advantage of applying a fungicide as a seed dressing increased moving from a quick and prompt emergence, associated with high air and soil temperatures, to a slow process related to a low-growing degree accumulation [
25]. Early sowings are often associated with low soil temperatures, but are also related to a higher water content in the soil, which in turn leads to slow and uneven emergence that promote seed-borne and soil-borne pathogens such as
Fusarium [
26].
As far as the comparison of seed-borne fungal species is concerned (study II),
F. verticillioides led to a higher seedling mortality and grain yield loss than
F. graminearum under the considered conditions. However, in experiments carried out under controlled conditions in Iowa [
3,
14] and in Brazil [
9],
F. graminearum was the most aggressive
Fusarium species that affected maize emergence. The different susceptibility to the two
Fusarium species could depend on the pathogenicity of the strains [
27] used to infect the seeds and on their interaction with different environmental conditions during germination.
As observed in other research, most of the negative impacts of seed-borne fungal infection are due to the loss of plants that occurs during the emergence stages [
21]. In the present study, the main cause of the yield gap, compared to the uninfected control, was clearly due to the decrease in the number of emerged seedlings, while no further loss of plants was observed in the successive growth stages until harvesting in any of the considered seedling mortality groups. Thus, the effect of
Fusarium seed-borne infection on crop damping-off just seems to be concentrated in the germination phases. Furthermore, study II underlines that the loss of plant population, when it is lower than 15%, could be compensated by an increase in production of the single plant, resulting in a similar grain yield.
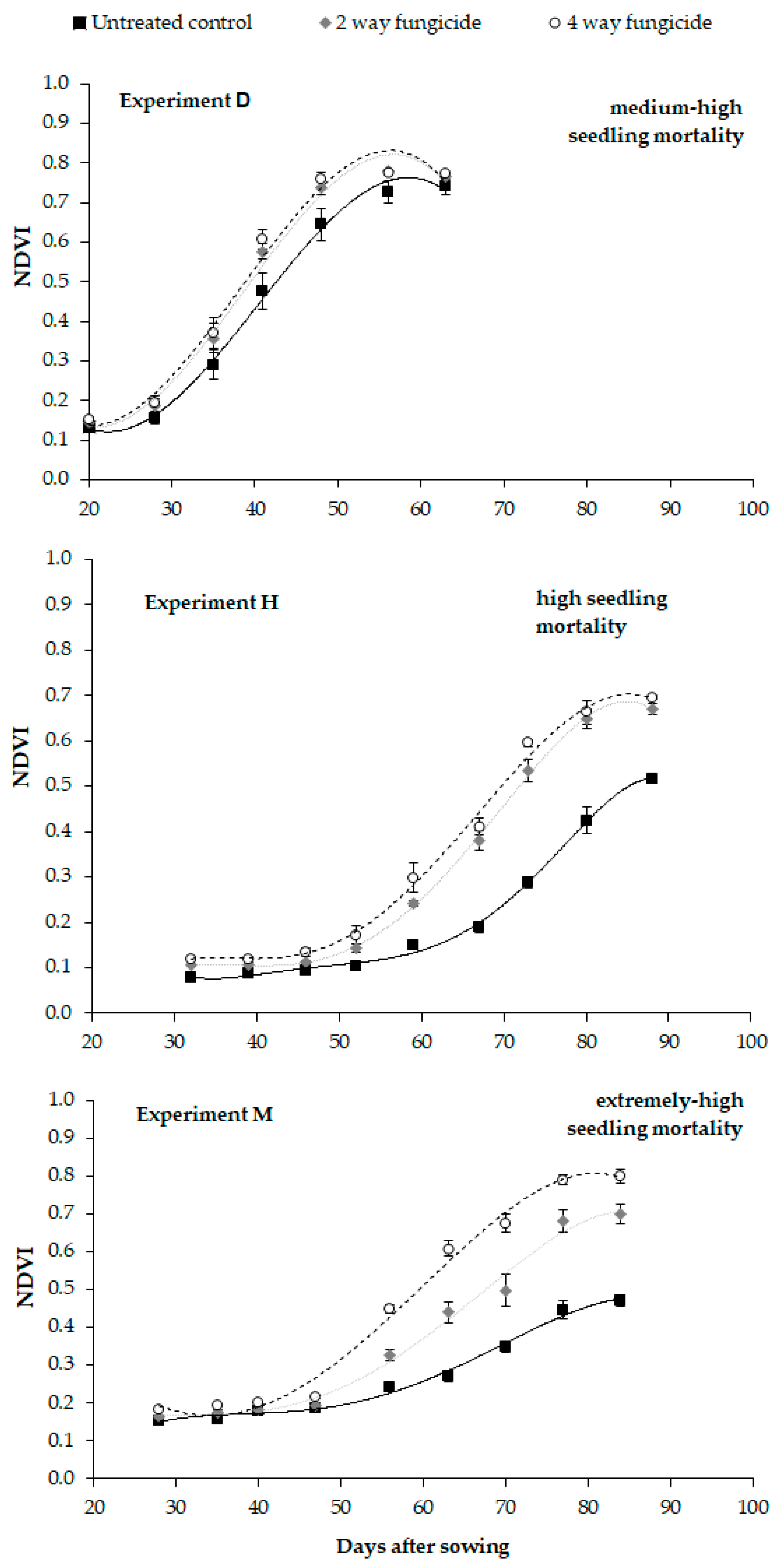
Nevertheless, the fungus activity, apart from influencing plant density, also has an effect on plant vigor and growth, and this was more evident in the experiments where fungal infection was more severe. Plants grown from artificially infected seeds clearly showed a slower growth than the uninfected control (study II); the height, measured at the elevation stage, the NDVI values, collected during the whole growing cycle, and thus AUCDC, were significantly lower. The infection of the
Fusarium-inoculated seeds also slowed down plant development: in study II, the flowering date was postponed by about 2 days (approximately 30 GDDs) compared to the uninfected control. Pinto et al. [
28] reported that systemic
F. verticillioides infection in maize plants affected their photosynthetic performance, mainly as a consequence of a reduction in chlorophyll content, which in turn led to a decrease in the electron transport components and a consequent reduction in carbohydrate synthesis.
The fungicide seed treatments reduced the loss, and in some cases removed the gap in the expected plant density, and this led to no difference in the grain yield with the uninfected control (Study II). The seed dressing, apart from being effective in ensuring the desired plant density, also allowed a faster growth of the plants than those of the infected control, as it controlled the systemic infection of both
Fusarium species. Previous studies, which were only carried out under controlled conditions (greenhouse), have reported a significant effect of the application of a fludioxonil and metalaxyl-M mixture on the plant vigor of infected maize [
14,
29] or soybean [
17,
30], as quantified by a higher dry mass of both the shoots and roots. Moreover, Rodriguez-Brljevich [
20] reported that a fungicide seed dressing suppresses the soil-borne infection of
Fusarium spp. in open fields, and results in enhanced photosynthesis and increased plant vigor. To the best of the authors’ knowledge, our study is the first that has quantified the advantage in vigor associated with the control of
Fusarium seed-born infection through a fungicide application in open fields, considering the complete growing cycle until harvest. In our medium-high seedling mortality experiments, the four-way fungicide did not increase the plant population at harvest, compared to the conventional two-way seed dressing, while the broad spectrum treatment increased plant vigor, resulting in a 16% grain yield increase. The seed dressing treatments resulted in a less detrimental vegetative growth, as a consequence of
Fusarium infection, as highlighted by the NDVI measurement, which thwarted any possible delay in the flowering date. This effect could contribute to enhancing the competitiveness of maize, since a delay in flowering and in the consequent ripening is associated with a lower grain yield (e.g., lower solar radiation interception, [
31]), a delay in the harvest date, or a higher grain moisture content at harvesting, and a higher risk of mycotoxin kernel contamination, because of late ripening, as well as a higher incidence of European corn borer injuries on the ears [
11].
As far as the
F. verticillioides infection is concerned, the broad-spectrum seed treatment (four-way) has proved to be more effective than the two-way fungicide one, and to result in a further significant advantage, even in the production situations with a lower disease pressure. The spectrum of the considered two-way mixture was probably not able to provide an analogous effective control of this pathogen in the considered growing areas, where
F. verticillioides is the predominant and the more harmful species [
5,
32]. As noted in other works, the use of a greater number of active ingredients leads to a broad spectrum of action, which in turn leads to significant improvements in the control of fungal pathogens and, in particular, of
Fusarium [
21,
33]. In addition, the use of active systemic fungicide ingredients with a greater ability to move in seedling tissue could significantly enhance the early season management of this disease. Benzimidazoles (thiabendazole), strobilurins (azoxystrobin), triazoles and pyrazole carboxamide are all able to penetrate the coating of maize seeds and translocate in the xylem to the endosperm, embryo, coleoptiles and radicle [
15]. This could make these ingredients more active in controlling the detrimental effects of such systemic pathogens as
F. verticillioides. Field experience with wheat [
34] highlighted that systemic fungicides have eradicant properties and are able to slow down the progress of existing infections.
Conversely, the four-way fungicide did not induce any further advantage under the
F. graminearum infection conditions. The benzimidazoles and strobilurins probably did not increase the control already provided by fludioxonil, which is highly effective in protecting seedlings from seed-borne
F. graminearum infection [
25]. Furthermore, the application of broad-spectrum fungicides may also determine an indirect advantage for diseases that are already well-controlled by simpler fungicide mixtures, in particular by reducing the risk of resistances [
35]. In fact, although
F. graminearum was included in group E (medium-low risk resistance) by the Fungicide Resistance Action Committee (FRAC), the resistance of its strains to fludioxonil has been reported [
25]. In our conditions, the loss of vigor associated with infection from
F. graminearum was significant, although less evident than that induced by
F. verticillioides. The two-way fungicide seed dressing was able to prevent this negative effect on plant vigor [
19], and an earlier flowering date than for the infected control was observed. As far as vegetative growth is concerned, the four-way fungicide did not lead to any further improvements in crop development or in the anticipation of flowering, compared to the two-way seed treatment.
A direct crop enhancement effect of fungicide seed dressing may be related to the physiological effect that certain fungicide compounds could exert on plants, even in the absence of a fungal infection. Strobilurins have been shown to induce physiological benefits for different crops, such as longer-lasting green leaf tissue and delayed plant senescence (stay green effect), through a reduction in oxidative stress [
36], an increase in photosynthesis efficiency, for higher true photosynthesis, and a reduction in dark respiration [
37]. Enhanced maize performance, even in the absence of disease, has also been reported for foliar applications of azoxystrobin [
38] and pyraclostrobin [
39] from the stem elongation stage to flowering. Conversely, no significant effects have been reported for earlier growing stage applications (five leaf-stage, [
38]), and no data are available concerning the physiological effect of strobilurins applied to maize as seed dressings. The application of pyraclostrobin to soybean seeds under disease-free conditions improved the growth, vigor (plant height, root and shoot dry mass) and chlorophyll index after 14 days of emergency [
40], while strobilurins enhanced rice seedling growth after root cutting injury by inducing reactive oxygen scavenging activity, thus inhibiting reactive oxygen species accumulation [
41]. Under controlled sterilized conditions, pyrazole carboxamide sedaxane facilitates root establishment and intensifies nitrogen and the phenylpropanoid metabolism of maize seedlings [
42].
In conclusion, the reported field experiments have confirmed the impact of the seed-borne infection of the two most common seed pathogens, F. verticillioides and F. graminearum, and quantified the negative effect of infection in different production situations from plant emergence to harvest. In addition, the collected data have highlighted the effectiveness of seed dressings with different fungicide treatments by detecting the advantages, in terms of plant population defense, stimulation of the plant development and final grain yield. The benefits of the broad-spectrum four-way formulation, compared to the conventional two-way fungicide seed dressing, clearly depend on the considered pathogens. As far as the F. verticillioides infection is concerned, the four-way fungicide enhanced both seedling defense and plant vigor, which resulted in a grain yield improvement under different disease infection conditions. In temperate maize-growing areas where the soil and seed occurrence of F. verticillioides inoculum is widespread, the application of four-way fungicide as seed dressing could allow to more successfully anticipate sowing time, even under conservative tillage conditions, often more prone to seedling disease and slow plant development.







{kind=link}
{kind=link}
{kind=link}