Quality Improvement of Netted Melon (Cucumis melo L. var. reticulatus) through Precise Nitrogen and Potassium Management in a Hydroponic System



Abstract
:1. Introduction
2. Materials and Methods
2.1. Hydroponic Systems and Plant Materials
2.2. Experiment I: Reducing N and Modifying K Fertilisers
2.3. Experiment II: Optimisoptimisation of the Plant Growth and Fruit Quality by Finer Adjustments
2.4. Nutrient Solution Monitoring and Water Chemistry Analysis
2.5. Plant Growth and Fruit Quality
2.6. Blind Tasting Tests for Customers’ Preferences
2.7. Statistical Analysis
3. Results
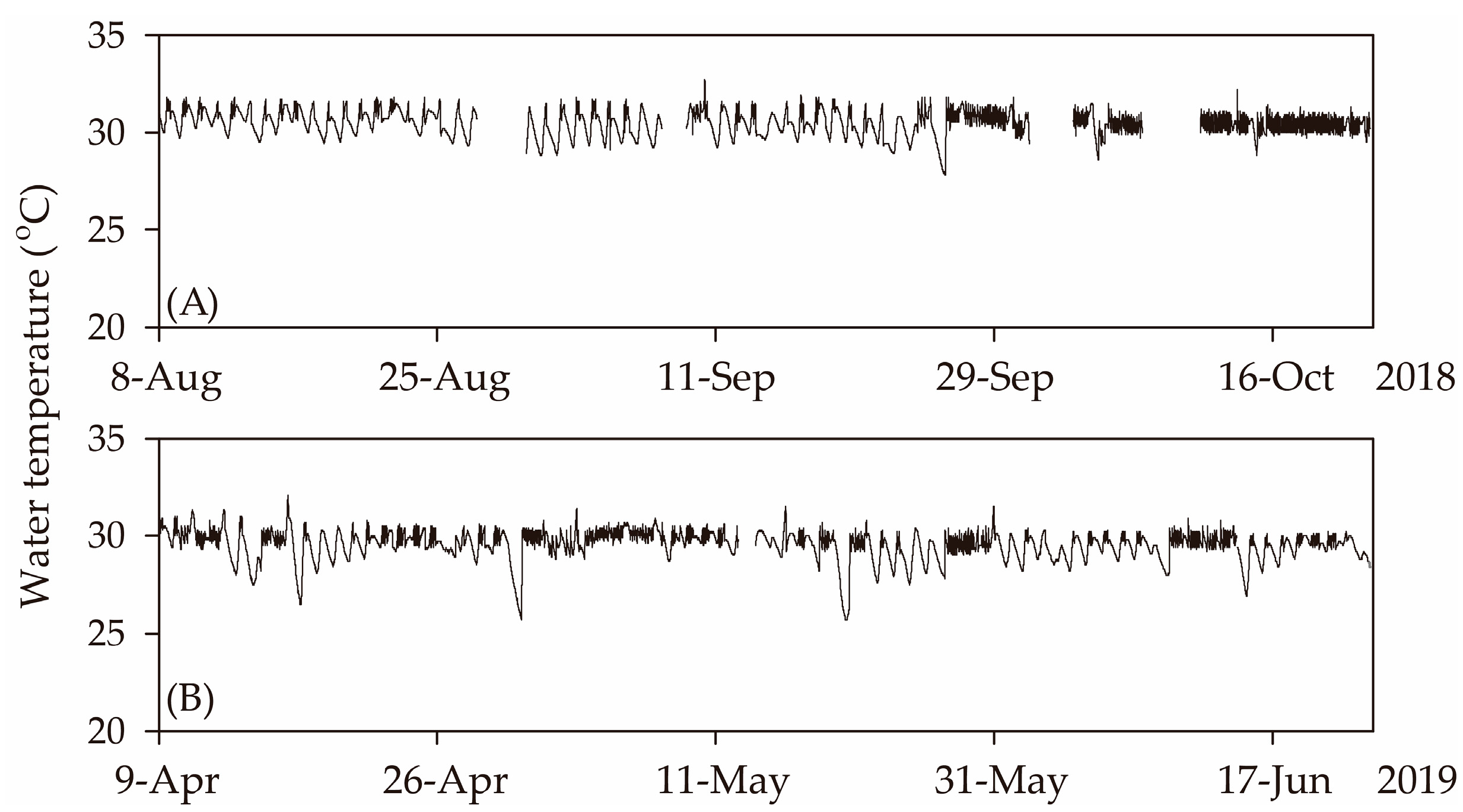
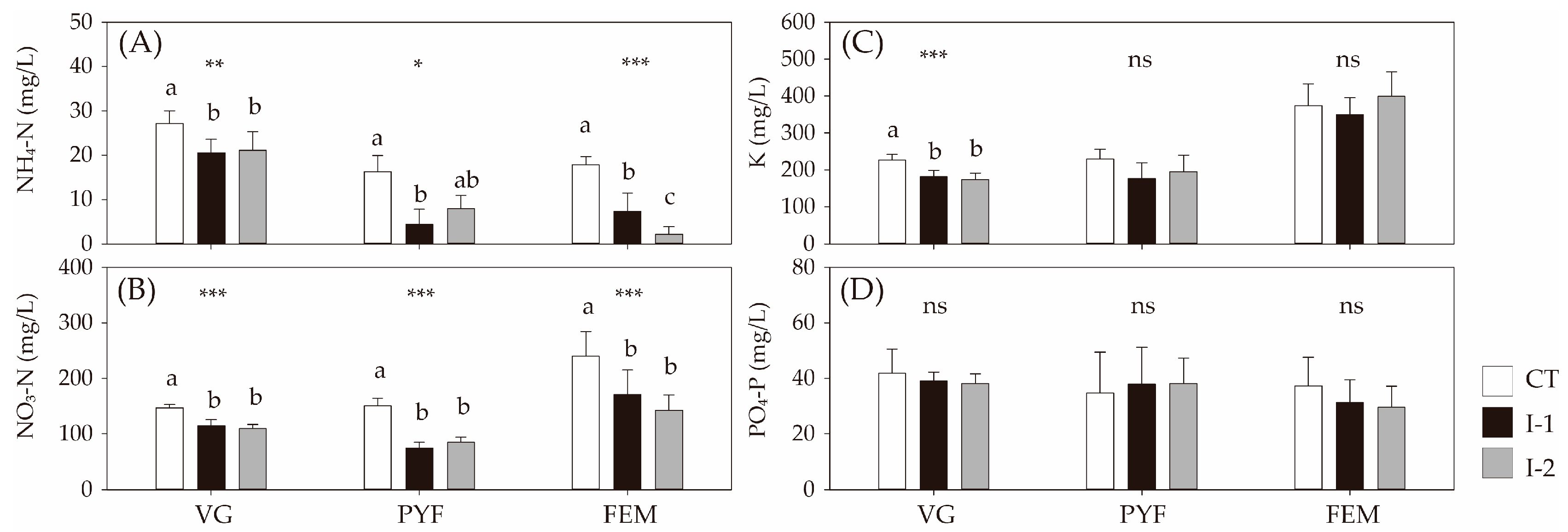
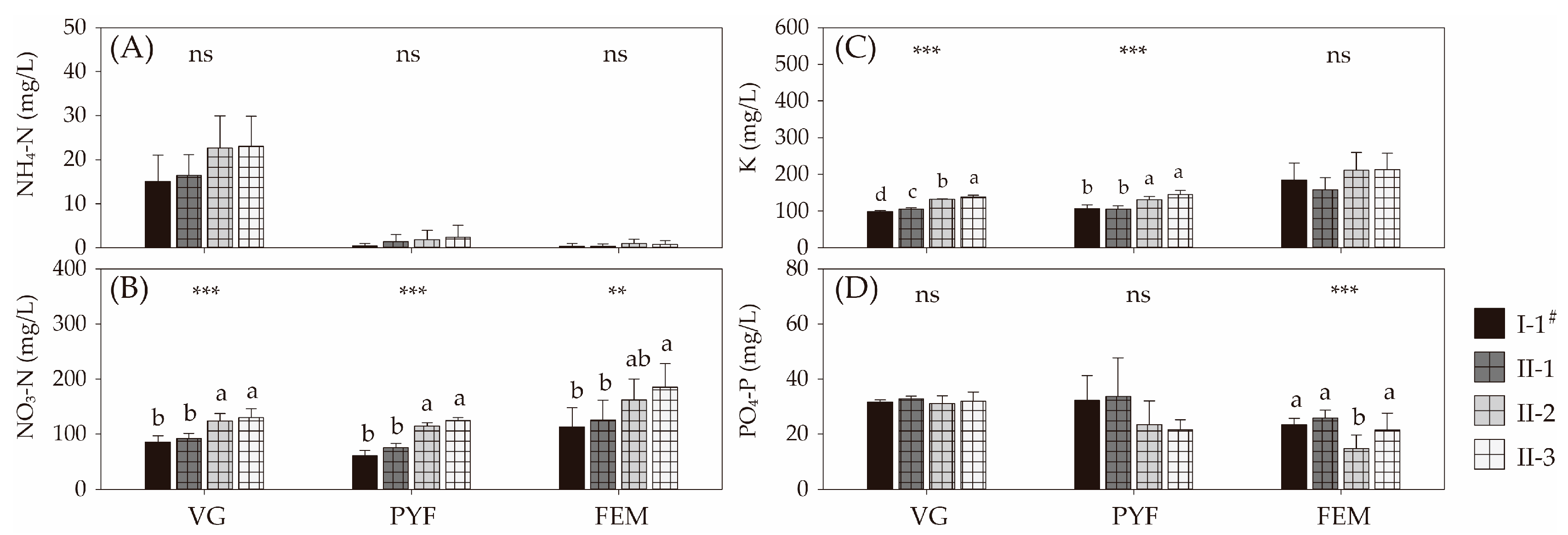
3.1. Changes in Water Chemistry of Nutrient Solution for the Hydroponic Culture
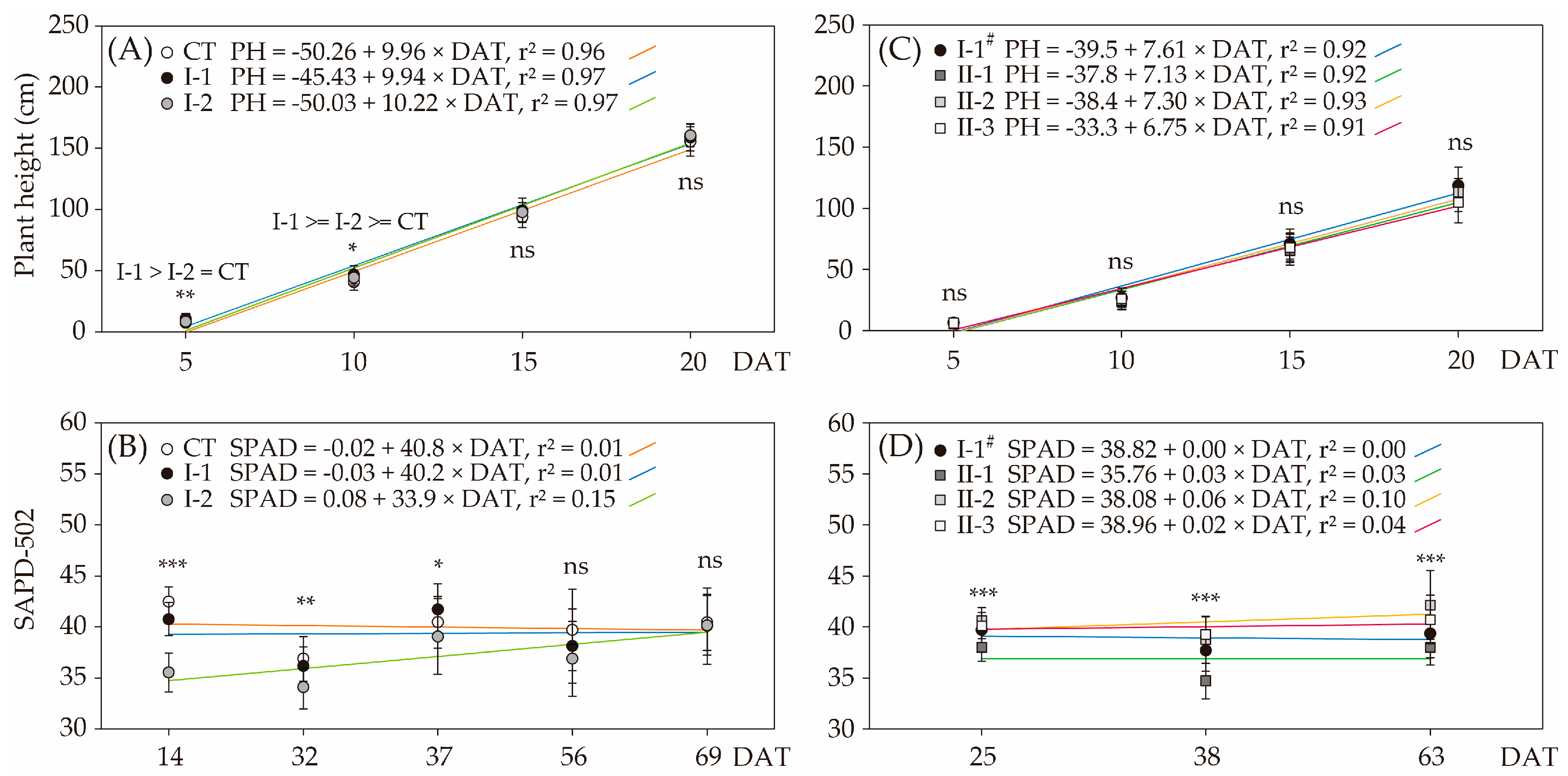
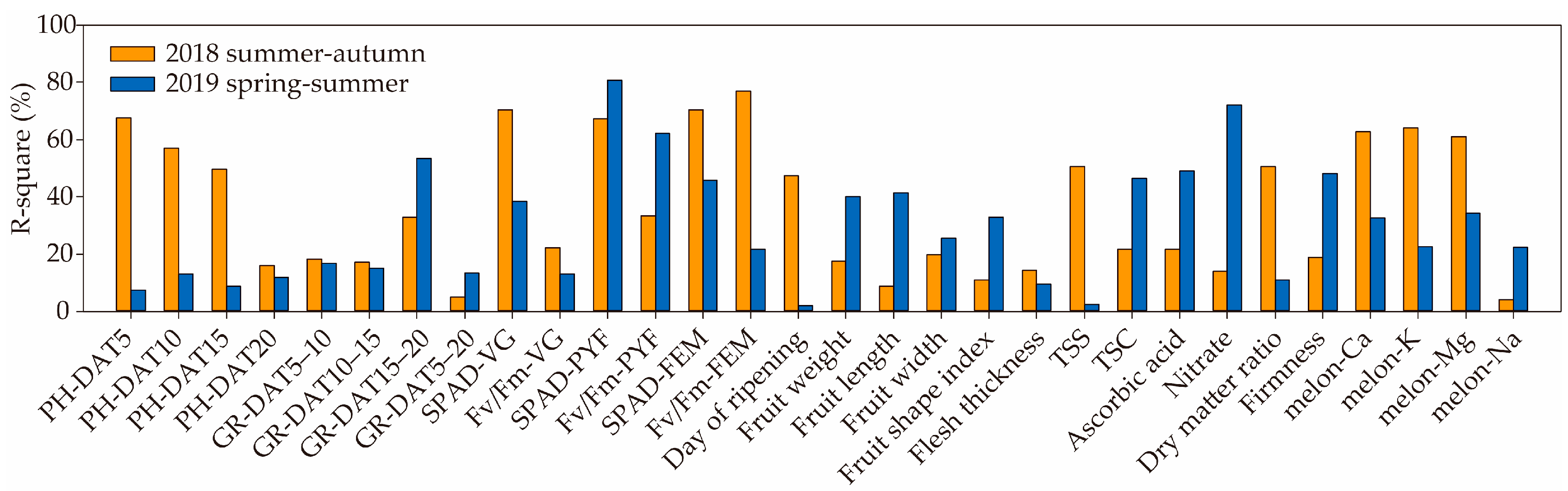
3.2. Plant Growth and Fruit Quality
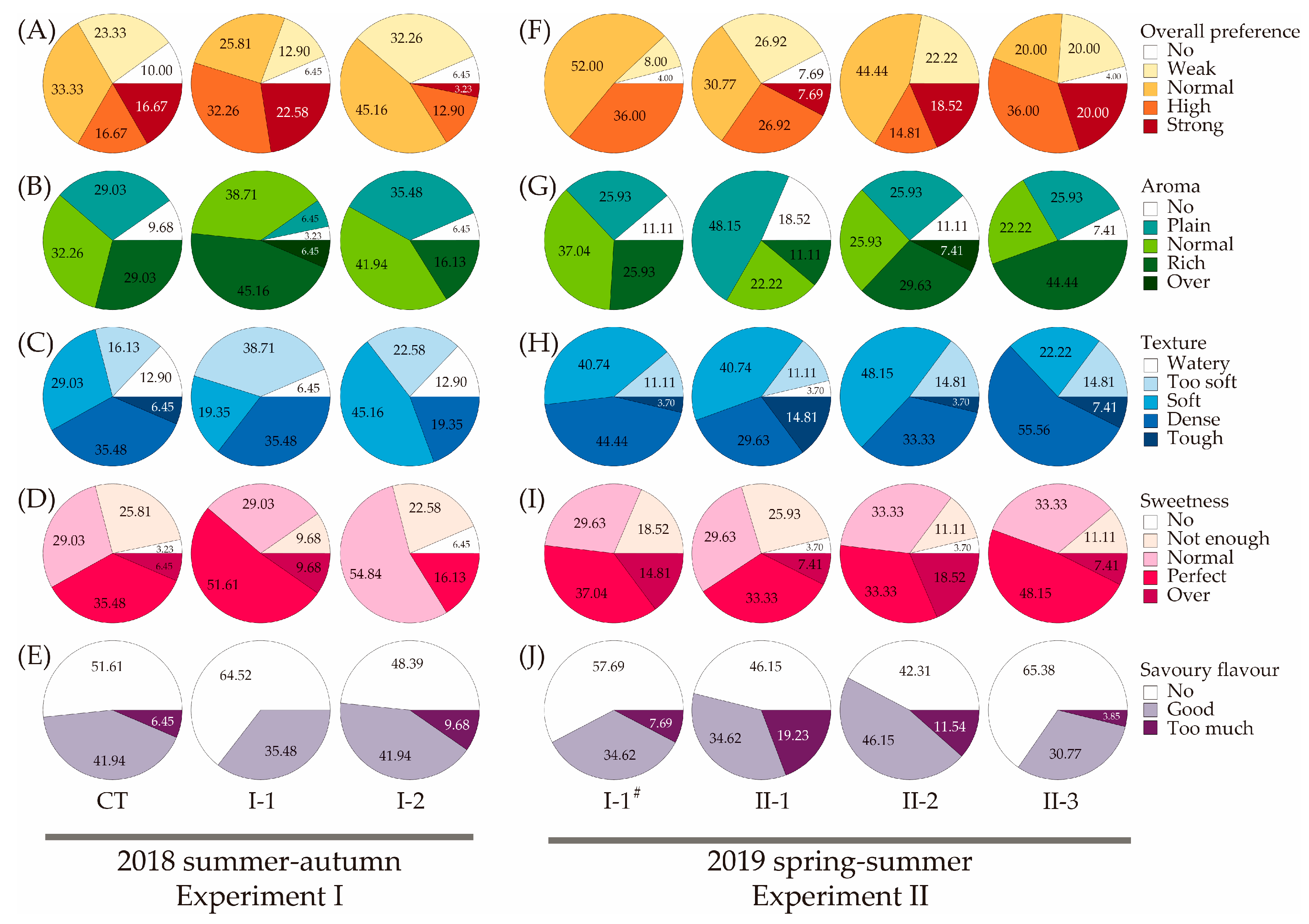
3.3. Blind Tasting Tests for Customers’ Preferences
4. Discussion
4.1. Changes in N Levels of Nutrient Solutions for Hydroponic Melon Culture
4.2. Plant Growth and Nutrient Manipulations
4.3. Fruit Quality and Nutrient Manipulations
4.4. Blind Tasting Tests for Customers’ Preferences
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rouphael, Y.; Colla, G. Growth, yield, fruit quality and nutrient uptake of hydroponically cultivated zucchini squash as affected by irrigation systems and growing seasons. Sci. Hortic. 2005, 105, 177–195. [Google Scholar] [CrossRef]
- Amalfitano, C.; Del Vacchio, L.; Somma, S.; Cuciniello, A.; Caruso, G. Effects of cultural cycle and nutrient solution electrical conductivity on plant growth, yield and fruit quality of ‘Friariello’ pepper grown in hydroponics. Hortic. Sci. 2017, 44, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Croft, M.M.; Hallett, S.G.; Marshall, M.I. Hydroponic production of vegetable amaranth (Amaranthus cruentus) for improving nutritional security and economic viability in Kenya. Renew. Agric. Food Syst. 2017, 32, 552–561. [Google Scholar] [CrossRef]
- Cardoso, F.B.; Martinez, H.E.P.; Da Silva, D.J.H.; Do Carmo Milagres, C.; Barbosa, J.G. Yield and quality of tomato grown in a hydroponic system, with different planting densities and number of bunches per plant. Pesq. Agropec. Trop. 2018, 48, 340–349. [Google Scholar] [CrossRef]
- Miceli, A.; Moncada, A.; Sabatino, L.; Vetrano, F. Effect of gibberellic acid on growth, yield, and quality of leaf lettuce and rocket grown in a floating system. Agronomy 2019, 9, 382. [Google Scholar] [CrossRef] [Green Version]
- Cuquel, F.L.; Motta, A.C.V.; Tutida, I.; De Mio, L.L.M. Nitrogen and potassium fertilization affecting the plum postharvest quality. Rev. Bras. Frutic. 2011, 33, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.C.; Smith, M.W.; Penn, C.J.; Cheary, B.S.; Conaghan, K.J. Nitrogen, phosphorus, calcium, and magnesium applied individually or as a slow release or controlled release fertilizer increase growth and yield and affect macronutrient and micronutrient concentration and content of field-grown tomato plants. Sci. Hortic. 2016, 211, 420–430. [Google Scholar] [CrossRef] [Green Version]
- Asaduzzaman, M.; Talukder, M.R.; Tanaka, H.; Ueno, M.; Kawaguchi, M.; Yano, S.; Ban, T.; Asao, T. Production of low-potassium content melon through hydroponic nutrient management using perlite substrate. Front. Plant Sci. 2018, 9, 1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambo, P.; Nicoletto, C.; Giro, A.; Pii, Y.; Valentinuzzi, F.; Mimmo, T.; Lugli, P.; Orzes, G.; Mazzetto, F.; Astolfi, S.; et al. Hydroponic solutions for soilless production systems: Issues and opportunities in a smart agriculture perspective. Front. Plant Sci. 2019, 10, 923. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, S.; Sapkota, S.; Liu, Z. Effects of nutrient composition and lettuce cultivar on crop production in hydroponic culture. Horticulturae 2019, 5, 72. [Google Scholar] [CrossRef] [Green Version]
- Sanjuan-Delmás, D.; Josa, A.; Muñoz, P.; Gassó, S.; Rieradevall, J.; Gabarrell, X. Applying nutrient dynamics to adjust the nutrient-water balance in hydroponic crops. A case study with open hydroponic tomato crops from Barcelona. Sci. Hortic. 2020, 261, 108908. [Google Scholar] [CrossRef]
- Kaur, H.; Bedi, S.; Sethi, V.P.; Dhatt, A.S. Effects of substrate hydroponic systems and different N and K ratios on yield and quality of tomato fruit. J. Plant Nutr. 2018, 41, 1547–1554. [Google Scholar] [CrossRef]
- Janpen, C.; Kanthawang, N.; Inkham, C.; Tsan, F.Y.; Sommano, S.R. Physiological responses of hydroponically-grown Japanese mint under nutrient deficiency. PeerJ 2019, 7, e7751. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chen, L.; Cao, M.; Zhang, X.; Li, S. Effects of leaf potassium content on growth dynamics of muskmelon and developing a coupling model of nitrogen and potassium. HortScience 2019, 54, 1397–1408. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Longnecker, N.; Dracup, M. Nitrogen deficiency slows leaf development and delays flowering in narrow-leafed lupin. Ann. Bot. 1997, 79, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Souri, M.K.; Dehnavard, S. Tomato plant growth, leaf nutrient concentrations and fruit quality under nitrogen foliar applications. Adv. Hortic. Sci. 2017, 32, 41–47. [Google Scholar] [CrossRef]
- Xiong, Q.; Tang, G.; Zhong, L.; He, H.; Chen, X. Response to nitrogen deficiency and compensation on physiological characteristics, yield formation, and nitrogen utilization of rice. Front. Plant Sci. 2018, 9, 1075. [Google Scholar] [CrossRef] [Green Version]
- Roorda van Eysinga, J.P.N.L.; Smilde, K.W. Nutritional Disorders in Glasshouse Tomatoes, Cucumbers and Lettuce; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1981; p. 130. [Google Scholar]
- Castellanos, M.T.; Cabello, M.J.; Cartagena, M.D.C.; Tarquis, A.M.; Arce, A.; Ribas, F. Growth dynamics and yield of melon as influenced by nitrogen fertiliser. Sci Agric. 2011, 68, 191–199. [Google Scholar] [CrossRef]
- Omotoso, S.O.; Akinrinde, E.A. Effect of nitrogen fertiliser on some growth, yield and fruit quality parameters in pineapple (Ananas comosus L. Merr.) plant at Ado-Ekiti Southwestern, Nigeria. IRJAS 2013, 3, 11–16. [Google Scholar]
- Peña-Fleitas, M.T.; Gallardo, M.; Thompson, R.B.; Farneselli, M.; Padilla, F.M. Assessing crop N status of fertigated vegetable crops using plant and soil monitoring techniques. Ann. Appl. Biol. 2015, 167, 387–405. [Google Scholar] [CrossRef]
- Mendoza-Tafolla, R.O.; Juarez-Lopez, P.; Ontiveros-Capurata, R.-E.; Sandoval-Villa, M.; Alia-Tejacal, I.; Alejo-Santiago, G. Estimating nitrogen and chlorophyll status of romaine lettuce using SPAD and at LEAF readings. Not. Bot. Horti. Agrobot. Cluj-Napoca 2019, 47. [Google Scholar] [CrossRef] [Green Version]
- Cavalcante, V.S.; Prado, R.D.M.; Vasconcelos, R.D.L.; Almeida, H.J.D.; Silva, T.R.D. Growth and nutritional efficiency of watermelon plants grown under macronutrient deficiencies. HortScience 2019, 54, 738–742. [Google Scholar] [CrossRef] [Green Version]
- Andriolo, J.L.; Erpen, L.; Cardoso, F.L.; Cocco, C.; Casagrande, G.S.; Jänisch, D.I. Nitrogen levels in the cultivation of strawberries in soilless culture. Hortic. Bras. 2011, 29, 516–519. [Google Scholar] [CrossRef] [Green Version]
- Neilsen, G.; Kappel, F.; Neilsen, D. Fertigation and crop load affect yield, nutrition, and fruit quality of ‘Lapins’ sweet cherry on Gisela 5 rootstock. HortScience 2007, 42, 1456–1462. [Google Scholar] [CrossRef] [Green Version]
- Goyal, S.S.; Huffaker, R.C. Nitrogen Toxicity in Plants. In Nitrogen in Crop Production; Hauck, R.D., Ed.; American Society of Agronomy: Madison, WI, USA, 1984; pp. 97–118. [Google Scholar] [CrossRef]
- Fernández-Escobar, R.; Beltrán, G.; Sánchez-Zamora, M.A.; García-Novelo, J.; Aguilera, M.P.; Uceda, M. Olive oil quality decreases with nitrogen over-fertilisation. HortScience 2006, 41, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, T.T.; Cochrane, T.A. The vital role of potassium in the osmotic mechanism of stomata aperture modulation and its link with potassium deficiency. Plant Signal. Behav. 2009, 4, 240–243. [Google Scholar] [CrossRef] [Green Version]
- Lester, G.E.; Jifon, J.L.; Makus, D.J. Impact of potassium nutrition on postharvest fruit quality: Melon (Cucumis melo L) case study. Plant Soil 2010, 335, 117–131. [Google Scholar] [CrossRef]
- Mondal, M.F.; Asaduzzaman, M.; Ueno, M.; Kawaguchi, M.; Yano, S.; Ban, T.; Tanaka, H.; Asao, T. Reduction of potassium (K) content in strawberry fruits through KNO3 management of hydroponics. Hort. J. 2017, 86, 26–36. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Reddy, V.R. Potassium starvation limits soybean growth more than the photosynthetic processes across CO2 levels. Front. Plant Sci. 2017, 8, 991. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Hossain, M.S.; Mahmud, J.A.; Hossen, M.S.; Masud, A.A.C.; Fujita, M. Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.; Huang, D.; Wang, S. Effects of potassium levels on fruit quality of muskmelon in soilless medium culture. Sci. Hortic. 2004, 102, 53–60. [Google Scholar] [CrossRef]
- Lester, G.E.; Jifon, J.L.; Rogers, G. Supplemental foliar potassium applications during muskmelon fruit development can improve fruit quality, ascorbic acid, and beta-carotene contents. J. Am. Soc. Hort. Sci. 2005, 130, 649–653. [Google Scholar] [CrossRef]
- Çolpan, E.; Zengin, M.; Özbahçe, A. The effects of potassium on the yield and fruit quality components of stick tomato. Hortic. Environ. Biotechnol. 2013, 54, 20–28. [Google Scholar] [CrossRef]
- Corrêa, C.V.; de Sousa Gouveia, A.M.; Martins, B.N.M.; Lanna, N.D.B.L.; Tavares, A.E.B.; Mendonça, V.Z.; Jorge, L.G.; Inácio Cardoso, A.I.; Evangelista, R.M. Effect of top-dressed potassium fertilisation on the yield and quality of cucumber. J. Plant Nutr. 2018, 41, 1345–1350. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Hoopen, F.T.; Cuin, T.A.; Pedas, P.; Hegelund, J.N.; Shabala, S.; Schjoerring, J.K.; Jahn, T.P. Competition between uptake of ammonium and potassium in barley and arabidopsis roots: Molecular mechanisms and physiological consequences. J. Exp. Bot. 2010, 61, 2303–2315. [Google Scholar] [CrossRef] [Green Version]
- Berdiyev, M.; Arslan, D.; Özcan, M.M. Nutritional composition, microbiological and sensory properties of dried melon: A traditional turkmen product. Int. J. Food Sci. Nutr. 2009, 60, 60–68. [Google Scholar] [CrossRef]
- Schaffer, A.A.; Paris, H.S. Melons, Squashes, and Gourds. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- MAFF. The 93rd Statistical Yearbook of Ministry of Agriculture, Forestry and Fisheries; Ministry of Agriculture, Forestry and Fisheries: Tokyo, Japan, 2019. Available online: https://www.maff.go.jp/e/data/stat/93th/index.html (accessed on 13 March 2020).
- Agriculture and Food Agency. Basic Agricultural Statistics 2018; Agriculture and Food Agency, Council of Agriculture: Taiwan, 2019. Available online: https://agrstat.coa.gov.tw/sdweb/public/book/Book.aspx (accessed on 13 March 2020). (In Chinese)
- Bouwkamp, J.C.; Angell, F.F.; Schales, F.D. Effects of weather conditions on soluble solids of muskmelon. Sci. Hortic. 1978, 8, 265–271. [Google Scholar] [CrossRef]
- Baker, J.T.; Reddy, V.R. Temperature effects on phenological development and yield of muskmelon. Ann. Bot. 2001, 87, 605–613. [Google Scholar] [CrossRef] [Green Version]
- Wiangsamut, B.; Koolpluksee, M.; Makhonpas, C. Yield, fruit quality, and growth of 4 cantaloupe varieties grown in hydroponic system and drip irrigation systems of substrate and soil culture. Int. J. Agric. Technol. 2017, 13, 1381–1394. [Google Scholar]
- Rodriguez, J.C.; Cantliffe, D.J.; Shaw, N.L.; Karchi, Z. Soilless media and containers for greenhouse production of ‘Galia’ type muskmelon. HortScience 2006, 41, 1200–1205. [Google Scholar] [CrossRef] [Green Version]
- Singer, S.M.; El-Tanahy, A.M.M.; El-Behairy, U.A.; El-Samad, E.H.A. Growth and productivity of cantaloupe plants grown under different soilless culture systems. J. Appl. Sci. Res. 2013, 9, 5294–5302. [Google Scholar]
- Yamasaki, K.; Suzuki, Y.; Shinohara, Y. Studies on water culture of several vegetables with special reference to the control of nutrient solution and ratio of nutrients absorption per water consumption. In Memoirs of the Faculty of Agriculture; Tokyo University of Education: Ibaraki, Japan, 1976. [Google Scholar]
- Sheen, T.F. Effects of NO3/NH4 ratio and depth of nutrient solution on growth of greenhouse muskmelon. J. Agric. Res. China 1989, 38, 53–71. (In Chinese) [Google Scholar]
- Sheen, T.F.; Hsu, M.M. Studies on nutrient uptake of greenhouse muskmelon (Cucumis melo L.) by hydroponics. J. Agric. Res. China 1990, 39, 55–64. (In Chinese) [Google Scholar] [CrossRef]
- Lobet, G.; Draye, X.; Périlleux, C. An online database for plant image analysis software tools. Plant Methods 2013, 9, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, K.K.; Price, T.V.; Hanger, B.C. Nutritional requirements for growth and yield of strawberry in deep flow hydroponic systems. Sci. Hortic. 1992, 52, 95–104. [Google Scholar] [CrossRef]
- Guo, X.; van Iersel, M.W.; Chen, J.; Brackett, R.E.; Beuchat, L.R. Evidence of association of salmonellae with tomato plants grown hydroponically in inoculated nutrient solution. Appl. Environ. Microbiol. 2002, 68, 3639. [Google Scholar] [CrossRef] [Green Version]
- Akl, I.A.; Savvas, D.; Papadantonakis, N.; Lydakis-Simantiris, N.; Kefalas, P. Influence of ammonium to total nitrogen supply ratio on growth, yield and fruit quality of tomato grown in a closed hydroponic system. Eur. J. Hortic. Sci. 2003, 68, 204–211. [Google Scholar]
- Hoque, M.M.; Ajwa, H.A.; Smith, R. Nitrite and ammonium toxicity on lettuce grown under hydroponics. Commun. Soil Sci. Plant Anal. 2007, 39, 207–216. [Google Scholar] [CrossRef]
- Erabadupitiya, H.R.U.T.; Weerakkody, W.A.P.; Nandasena, K.A. Determination of optimum nitrogen concentrations in hydroponics for tomato grown in coir medium in tropical greenhouse. Trop. Agric. Res. 2019, 30, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, H.; Osawa, T. Nitrate- and ammonium-N absorption by vegetables from nutrient solution containing ammonium nitrate and the resultant change of solution pH. J. Jpn. Soc. Hortic. Sci 1981, 50, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Zhou, Y.; Shen, Q.; Zhang, F. Effect of ammonium and nitrate nutrition on some physiological processes in higher plants-growth, photosynthesis, photorespiration, and water relations. Plant Biol. 2007, 9, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef] [Green Version]
- Huffman, J.R. Effects of enhanced ammonium nitrogen availability for corn. J. Agron. Educ. 1989, 18, 93–97. [Google Scholar] [CrossRef]
- Claussen, W.; Lenz, F. Effect of ammonium and nitrate on net photosynthesis, flower formation, growth and yield of eggplants (Solanum melongena L.). Plant Soil 1995, 171, 267–274. [Google Scholar] [CrossRef]
- Central Weather Bureau, Taiwan. Climate Statistics. Available online: https://www.cwb.gov.tw/V8/C/ (accessed on 10 March 2020).
- Ferrante, A.; Spinardi, A.; Maggiore, T.; Testoni, A.; Gallina, P.M. Effect of nitrogen fertilisation levels on melon fruit quality at the harvest time and during storage. J. Sci. Food Agric. 2008, 88, 707–713. [Google Scholar] [CrossRef]
- Sete, P.B.; Comin, J.J.; Nara Ciotta, M.; Almeida Salume, J.; Thewes, F.; Brackmann, A.; Toselli, M.; Nava, G.; Rozane, D.E.; Loss, A.; et al. Nitrogen fertilization affects yield and fruit quality in pear. Sci. Hortic. 2019, 258, 108782. [Google Scholar] [CrossRef]
- Asao, T.; Asaduzzaman, M.; Mondal, M.F.; Tokura, M.; Adachi, F.; Ueno, M.; Kawaguchi, M.; Yano, S.; Ban, T. Impact of reduced potassium nitrate concentrations in nutrient solution on the growth, yield and fruit quality of melon in hydroponics. Sci. Hortic. 2013, 164, 221–231. [Google Scholar] [CrossRef]
- Padilla, F.M.; de Souza, R.; Peña-Fleitas, M.T.; Gallardo, M.; Giménez, C.; Thompson, R.B. Different responses of various chlorophyll meters to increasing nitrogen supply in sweet pepper. Front. Plant Sci. 2018, 9, 1752. [Google Scholar] [CrossRef] [Green Version]
- Wahocho, N.A.; Maitlo, A.A.; Baloch, Q.B.; Kaleri, A.A.; Rajput, L.B.; Talpur, N.A.; Sheikh, Z.A.; Mengal, F.H.; Wahocho, S.A. Effect of varying levels of nitrogen on the growth and yield of muskmelon (Cucumis melo L.). J. Basic Appl. Sci. 2017, 13, 448–453. [Google Scholar]
- Johansen, C.; Edwards, D.G.; Loneragan, J.F. Interactions between potassium and calcium in their absorption by intact barley plants. I. Effects of potassium on calcium absorption. Plant Physiol. 1968, 43, 1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voogt, W. The growth of beefsteak tomato as affected by K/Ca ratios in the nutrient solution. Acta Hortic. 1988, 155–166. [Google Scholar] [CrossRef]
- Bar-Tal, A.; Pressman, E. Root restriction and potassium and calcium solution concentrations affect dry-matter production, cation uptake, and blossom-end rot in greenhouse tomato. J. Am. Soc. Hort. Sci. 1996, 121, 649–655. [Google Scholar] [CrossRef] [Green Version]
- Preciado-Rangel, P.; Salas-Pérez, L.; Gallegos-Robles, M.Á.; Ruiz-Espinoza, F.H.; Ayala-Garay, A.V.; Fortis-Hernández, M.; Murillo-Amador, B. Increasing doses of potassium increases yield and quality of muskmelon fruits under greenhouse. Hortic. Bras. 2018, 36, 184–188. [Google Scholar] [CrossRef]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of Macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 135–189. [Google Scholar] [CrossRef]
- Sams, C.E. Preharvest factors affecting postharvest texture. Postharvest Biol. Technol. 1999, 15, 249–254. [Google Scholar] [CrossRef]
- Wiesler, F. Nutrition and Quality. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 271–282. [Google Scholar] [CrossRef]
- Elbashier, M.A.M.; Xiaohou, S.; Ali, A.S.A.; Mohmmed, A. Effect of digestate and biochar amendments on photosynthesis rate, growth parameters, water use efficiency and yield of Chinese melon (Cucumis melo L.) under saline irrigation. Agronomy 2018, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Ortega, W.M.; Martínez, V.; Nieves, M.; Simón, I.; Lidón, V.; Fernandez-Zapata, J.C.; Martinez-Nicolas, J.J.; Cámara-Zapata, J.M.; García-Sánchez, F. Agricultural and physiological responses of tomato plants grown in different soilless culture systems with saline water under greenhouse conditions. Sci. Rep. 2019, 9, 6733. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Developmental Stage | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| VG | PYF | FEM | |||||||
| N | P | K | N | P | K | N | P | K | |
| CT | 168 (100) | 31 (100) | 195 (100) | 168 (100) | 31 (100) | 195 (100) | 168 (100) | 31 (100) | 195 (100) |
| I-1 | 126 (75) | 31 (100) | 146.25 (75) | 126 (75) | 31 (100) | 243.75 (125) | 168 (100) | 31 (100) | 195 (100) |
| I-2 | 126 (75) | 31 (100) | 146.25 (75) | 126 (75) | 31 (100) | 243.75 (125) | 126 (75) | 31 (100) | 243.75 (125) |
| Treatments | Plant Developmental Stage | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| VG | PYF | FEM | |||||||
| N | P | K | N | P | K | N | P | K | |
| I-1# | 126 (75) | 31 (100) | 146.25 (75) | 126 (75) | 31 (100) | 243.75 (125) | 168 (100) | 31 (100) | 195 (100) |
| II-1 | 126 (75) | 31 (100) | 146.25 (75) | 126 (75) | 31 (100) | 243.75 (125) | 168 (100) | 31 (100) | 146.25 (75) |
| II-2 | 168 (100) | 31 (100) | 195 (100) | 126 (75) | 31 (100) | 243.75 (125) | 168 (100) | 31 (100) | 195 (100) |
| II-3 | 168 (100) | 31 (100) | 195 (100) | 126 (75) | 31 (100) | 243.75 (125) | 168 (100) | 31 (100) | 146.25 (75) |
| Treatments | Days to Harvest (d) | Fruit Weight (g) | Fruit Shape Index (Length/Width) | Flesh Thickness (mm) | Pulp Firmness (N·m2) | Dry Matter Ratio (%) | TSS (°Brix) | TSC (mg/g) | Ascorbic Acid (mg/L) | Fruit K (mg/100 g FW) | Nitrate (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Experiment I, summer–autumn 2018 | |||||||||||
| Day length: decreasing trend from 13.18 to 11.28 h/d | |||||||||||
| CT | 54.0 c ± 1.3 | 1383.1 ab ± 118.8 | 0.99 ± 0.04 | 35.95 b ± 1.92 | 2.58 ± 0.44 | 7.63 c ± 0.61 | 12.51 b ± 1.19 | 0.49 b ± 0.06 | 76.8 ± 18.1 | 327.8 b ± 33.6 | 3.0 ± 6.2 |
| I-1 | 56.4 a ± 0.7 | 1506.9 a ± 143.7 | 1.00 ± 0.03 | 39.28 a ± 3.24 | 2.51 ± 0.88 | 10.76 a ± 0.35 | 14.09 a ± 0.93 | 0.50 ab ± 0.03 | 86.7 ± 26.2 | 410.3 a ± 22.0 | 1.3 ± 4.6 |
| I-2 | 55.2 b ± 1.1 | 1356.4 b ± 131.1 | 0.99 ± 0.02 | 36.50 b ± 2.90 | 2.15 ± 0.78 | 9.45 b ± 1.02 | 13.55 a ± 0.57 | 0.53 a ± 0.05 | 82.5 ± 29.1 | 402.0 a ± 22.3 | 3.8 ± 5.8 |
| F ratio | 14.99 *** | 4.47 * | 0.09 ns | 5.08 * | 1.14 ns | 33.94 *** | 9.00 ** | 3.36 * | 0.47 ns | 8.80 * | 0.59 ns |
| Experiment II, spring–summer 2019 | |||||||||||
| Day length: increasing trend from 12.60 to 13.68 h/d | |||||||||||
| I-1# | 54.7 ± 2.1 | 1192.0 ± 88.6 | 1.02 ± 0.03 | 35.43 ± 1.95 | 5.77 ab ± 0.73 | 11.27 ± 0.74 | 0.49 ± 0.04 | 176.3 ± 37.5 | 523.8 ± 46.0 | 12.4 ± 14.8 | |
| II-1 | 54.4 ± 3.0 | 1107.4 ± 84.6 | 1.00 ± 0.03 | 34.22 ± 2.26 | 6.07 a ± 0.62 | 10.78 ± 0.72 | 13.52 ± 0.71 | 0.53 ± 0.05 | 156.1 ± 42.1 | 508.9 ± 71.4 | 8.8 ± 6.0 |
| II-2 | 54.3 ± 2.1 | 1194.8 ± 63.7 | 1.02 ± 0.04 | 35.31 ± 3.07 | 5.27 ab ± 0.41 | 10.44 ± 1.12 | 13.44 ± 0.82 | 0.53 ± 0.02 | 135.6 ± 20.8 | 598.9 ± 119.4 | 4.4 ± 5.3 |
| II-3 | 55.1 ± 2.1 | 1203.2 ± 122.1 | 1.03 ± 0.02 | 34.71 ± 2.40 | 5.07 b ± 0.35 | 10.93 ± 1.12 | 13.61 ± 0.98 | 0.50 ± 0.05 | 140.3 ± 33.1 | 554.9 ± 25.2 | 8.0 ± 9.0 |
| F ratio | 0.19 ns | 2.13 ns | 0.97 ns | 0.47 ns | 4.79 ** | 1.18 ns | 0.33 ns | 2.07 ns | 2.59 ns | 0.86 ns | 1.06 ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yam, R.S.W.; Fan, Y.-T.; Lin, J.-T.; Fan, C.; Lo, H.-F. Quality Improvement of Netted Melon (Cucumis melo L. var. reticulatus) through Precise Nitrogen and Potassium Management in a Hydroponic System. Agronomy 2020, 10, 816. https://doi.org/10.3390/agronomy10060816
Yam RSW, Fan Y-T, Lin J-T, Fan C, Lo H-F. Quality Improvement of Netted Melon (Cucumis melo L. var. reticulatus) through Precise Nitrogen and Potassium Management in a Hydroponic System. Agronomy. 2020; 10(6):816. https://doi.org/10.3390/agronomy10060816
Chicago/Turabian StyleYam, Rita S.W., Yen-Tzu Fan, Jing-Tian Lin, Chihhao Fan, and Hsiao-Feng Lo. 2020. "Quality Improvement of Netted Melon (Cucumis melo L. var. reticulatus) through Precise Nitrogen and Potassium Management in a Hydroponic System" Agronomy 10, no. 6: 816. https://doi.org/10.3390/agronomy10060816
APA StyleYam, R. S. W., Fan, Y. -T., Lin, J. -T., Fan, C., & Lo, H. -F. (2020). Quality Improvement of Netted Melon (Cucumis melo L. var. reticulatus) through Precise Nitrogen and Potassium Management in a Hydroponic System. Agronomy, 10(6), 816. https://doi.org/10.3390/agronomy10060816