Effect of Gibberellin Responsive Reduced Height Allele Rht13 on Agronomic Traits in Spring Bread Wheat in Field Experiment in Non-Black Soil Zone

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Molecular Analysis
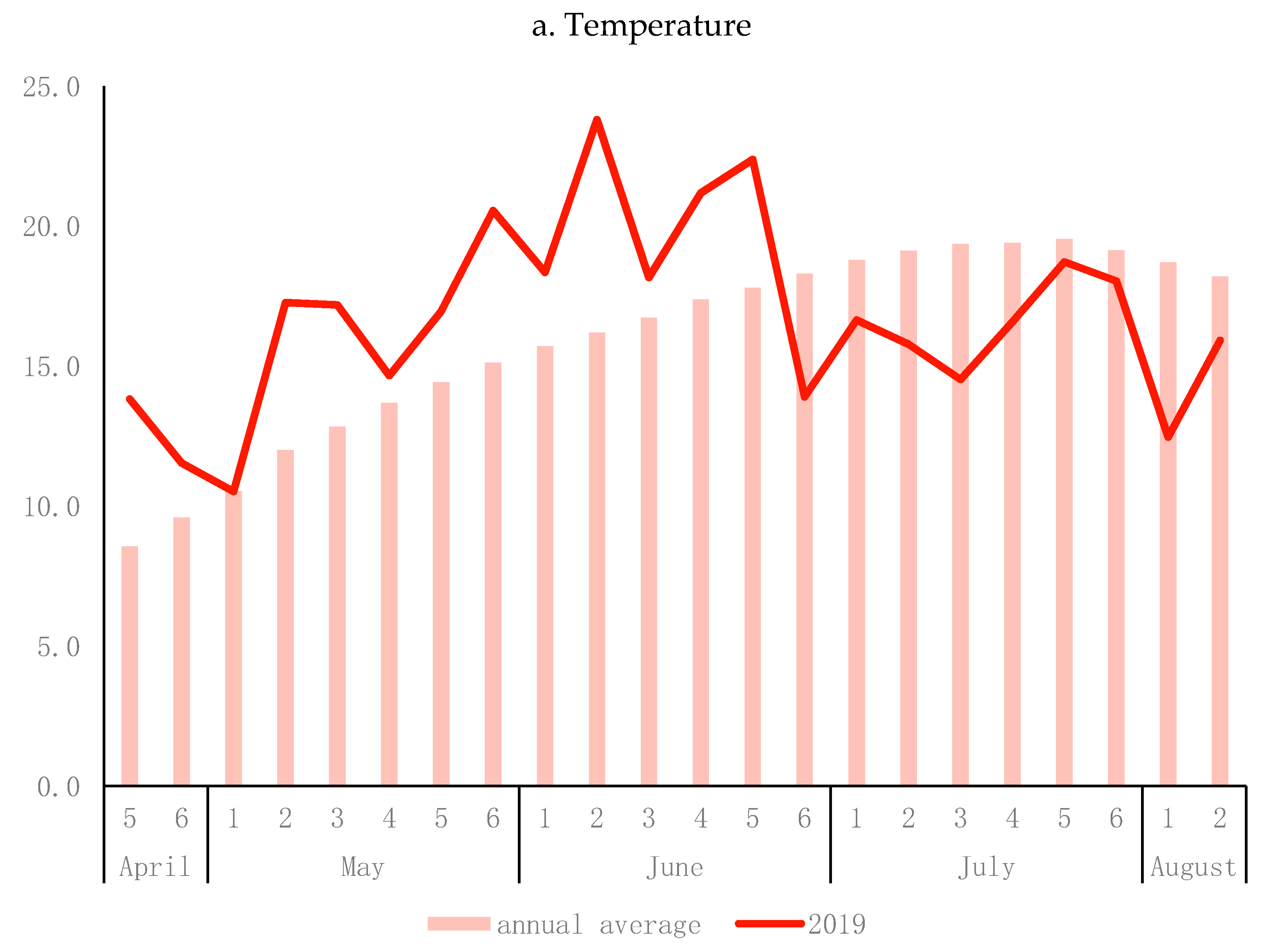
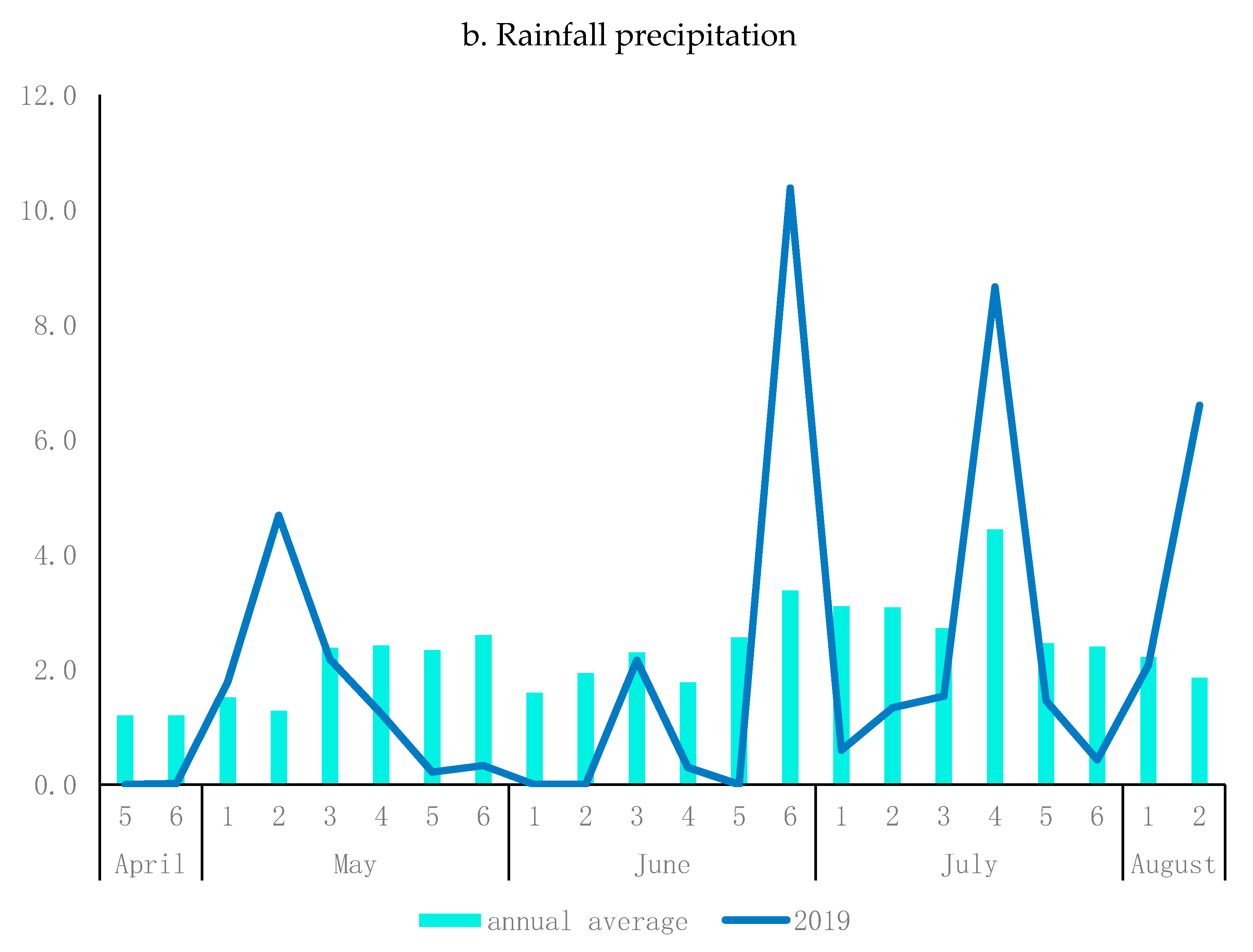
2.3. Field Experiment
2.4. Phenotyping
2.5. Statistical Analysis
3. Results
3.1. Anatomy of the Main Stem
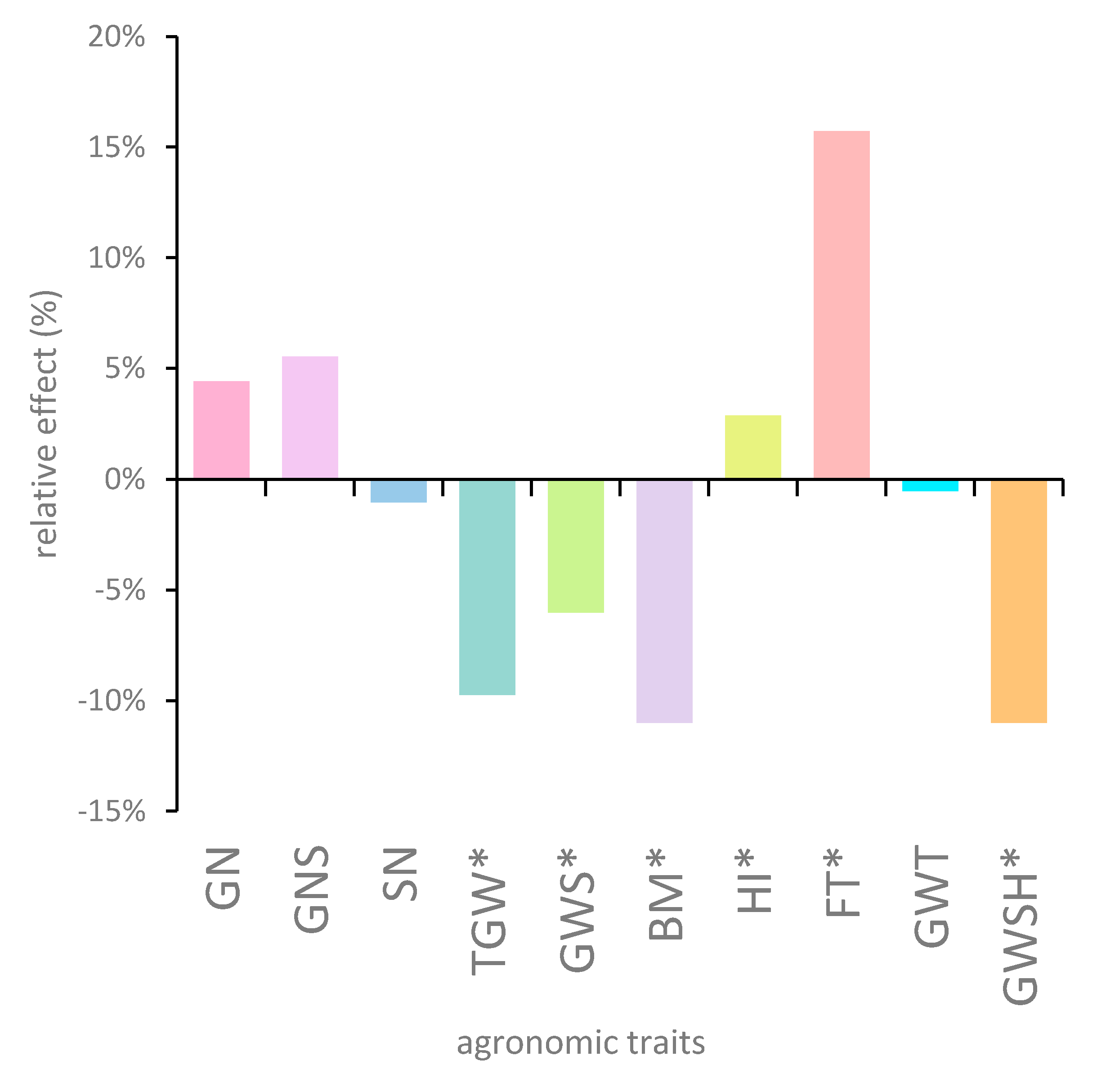
3.2. Spike Anatomy and Productivity
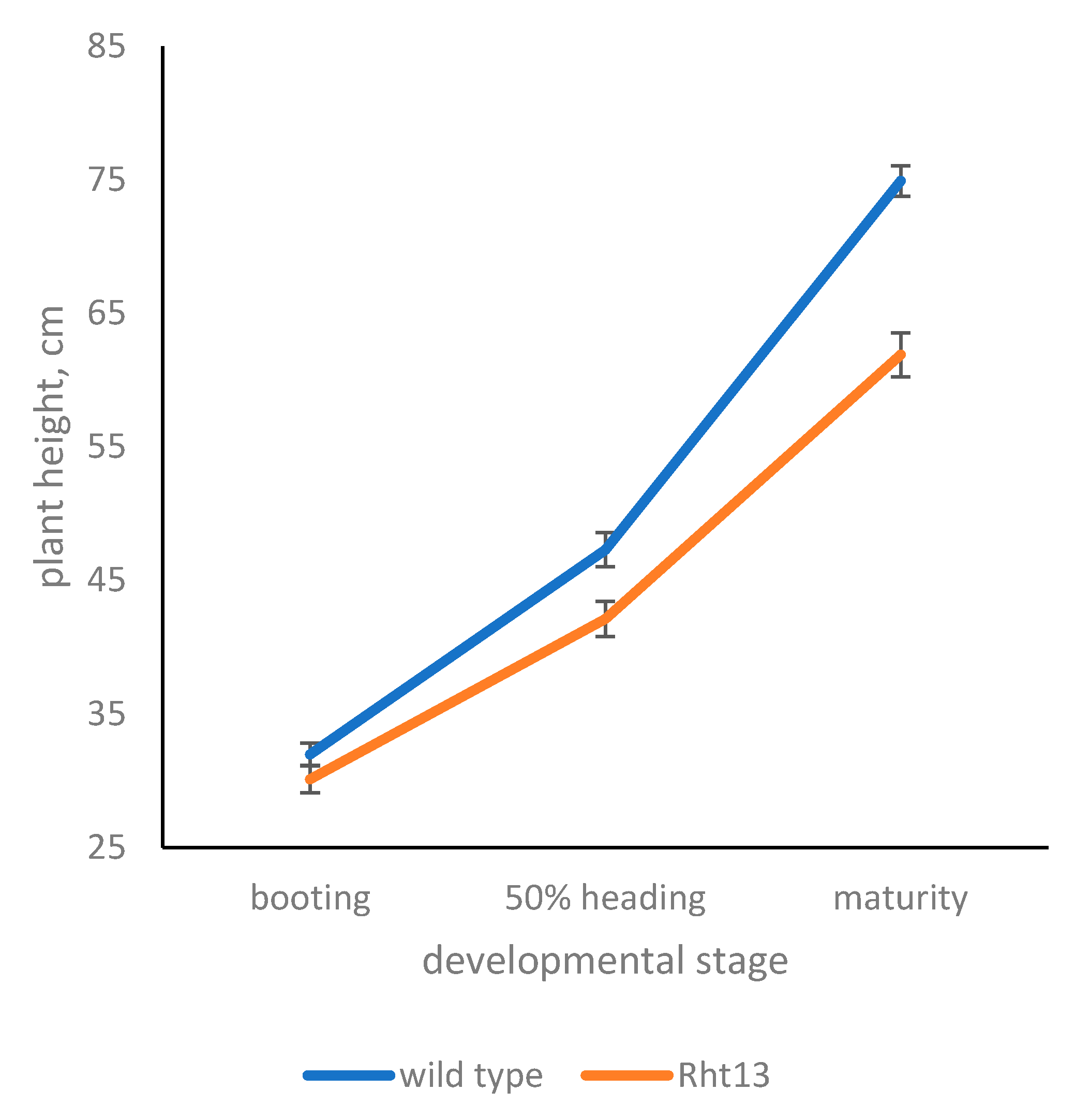
3.3. Harvest Index and Plant Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nhemachena, C.; Kirsten, J. A historical assessment of sources and uses of wheat varietal innovations in South Africa. S. Afr. J. Sci. 2017, 113, 1–8. [Google Scholar] [CrossRef]
- Catalogue of Gene Symbols//National BioResource Project. Komugi Wheat Genetic Resource Database. Available online: https://shigen.nig.ac.jp/wheat/komugi/genes/symbolClassList.jsp (accessed on 26 June 2020).
- Wang, Y.; Chen, L.; Du, Y.; Yang, Z.; Condon, A.; Hu, Y. Genetic effect of dwarfing gene Rht13 compared with Rht-D1b on plant height and some agronomic traits in common wheat (Triticum aestivum L.). Field Crop. Res. 2014, 162, 39–47. [Google Scholar] [CrossRef]
- Rebetzke, G.; Ellis, M.; Bonnett, D.; Richards, R. Molecular mapping of genes for coleoptile growth in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 114, 1173–1183. [Google Scholar] [CrossRef]
- Li, X.; Lan, S.; Liu, Y.; Gale, M.; Worland, T. Effects of different Rht-B1b, Rht-D1b and Rht-B1c dwarfing genes on agronomic characteristics in wheat. Cereal Res. Commun. 2006, 34, 919–924. [Google Scholar] [CrossRef]
- Ellis, M.; Rebetzke, G.; Chandler, P.; Bonnett, D.; Spielmeyer, W.; Richards, R. The effect of different height reducing genes on the early growth of wheat. Funct. Plant Biol. 2004, 31, 583. [Google Scholar] [CrossRef]
- Rebetzke, G.; Ellis, M.; Bonnett, D.; Condon, A.; Falk, D.; Richards, R. The Rht13 Dwarfing gene reduces peduncle length and plant height to increase grain number and yield of wheat. Field Crop. Res. 2011, 124, 323–331. [Google Scholar] [CrossRef]
- Rebetzke, G.; Ellis, M.; Bonnett, D.; Mickelson, B.; Condon, A.; Richards, R. Height reduction and agronomic performance for selected gibberellin-responsive dwarfing genes in bread wheat (Triticum aestivum L.). Field Crop. Res. 2012, 126, 87–96. [Google Scholar] [CrossRef]
- Chen, L.; Du, Y.; Lu, Q.; Chen, H.; Meng, R.; Cui, C.; Lu, S.; Yang, Y.; Chai, Y.; Li, J.; et al. The photoperiod-insensitive allele Ppd-D1a promotes earlier flowering in Rht12 dwarf plants of bread wheat. Front. Plant Sci. 2018, 9, 1312. [Google Scholar] [CrossRef]
- Mo, Y.; Vanzetti, L.; Hale, I.; Spagnolo, E.; Guidobaldi, F.; Al-Oboudi, J.; Odle, N.; Pearce, S.; Helguera, M.; Dubcovsky, J. Identification and characterization of Rht25, a locus on chromosome arm 6AS affecting wheat plant height, heading time, and spike development. Theor. Appl. Genet. 2018, 131, 2021–2035. [Google Scholar] [CrossRef] [Green Version]
- Divashuk, M.; Bespalova, L.; Vasilyev, A.; Fesenko, I.; Puzyrnaya, O.; Karlov, G. Reduced height genes and their importance in winter wheat cultivars grown in Southern Russia. Euphytica 2012, 190, 137–144. [Google Scholar] [CrossRef]
- Loskutova, N. The Use of Dwarfing Genes in the Selection of Spring Common Wheat. Ph.D. Thesis, N.I.Vavilov Institute of Plant Genetic Resources (VIR), Saint-Petersburg, Russia, 6 June 1996. Available online: http://earthpapers.net/preview/536914/a#?page=1 (accessed on 13 May 2020).
- Wang, Y.; Du, Y.; Yang, Z.; Chen, L.; Condon, A.; Hu, Y. Comparing the effects of GA-responsive dwarfing genes Rht13 and Rht8 on plant height and some agronomic traits in common wheat. Field Crop. Res. 2015, 179, 35–43. [Google Scholar] [CrossRef]
- Mutlucan, M.; Tosun, M.; Ilker, E.; Aykut Tonk, F.; Iştipliler, D.; Byrne, P.; Haley, S.; Maragues, M. Genetic Analysis of Plant Height, Coleoptile Length and Their Relationships With Rht12 and Rht13 Genes in Wheat (Conference Paper). Available online: https://www.researchgate.net/publication/303370625_Genetic_Analysis_of_Plant_Height_Coleoptile_Length_and_their_Relationships_with_Rht12_and_Rht13_Genes_in_Wheat (accessed on 13 May 2020).
- Maddocks, L.M. Molecular and Physiological Characterisation of Alternative Dwarfing Genes Rht12 and Rht13 in Bread Wheat (Triticum aestivum) and Rhtl8 in Durum Wheat (T. turgidum spp. durum). Master’s Thesis, Australian National University, Canberra, Australia, 2008. [Google Scholar]
- Zhang, N.; Yan, J.; Zhang, S. Rht13 dwarfing gene delays foliar senescence in wheat induced by nitrogen deficiency. Pak. J. Bot. 2019, 51, 143–147. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, N.; Wang, X.; Zhang, S. Differences in the physiological responses of Rht13 and rht wheat lines to short-term osmotic stress. Cereal Res. Commun. 2020, 48, 41–47. [Google Scholar] [CrossRef]
- Litvinov, D.; Chernook, A.; Kroupin, P.; Bazhenov, M.; Karlov, G.; Avdeev, S.; Divashuk, M. A Convenient co-dominant marker for height-reducing Ddw1 allele useful for marker-assisted selection. Agriculture 2020, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Komyshev, E.; Genaev, M.; Afonnikov, D. Evaluation of the SeedCounter, a mobile application for grain phenotyping. Front. Plant Sci. 2017, 7, 1990. [Google Scholar] [CrossRef] [Green Version]
- Ihaka, R.; Gentleman, R. R: A language for data analysis and graphics. J. Comp. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- Tshikunde, N.; Mashilo, J.; Shimelis, H.; Odindo, A. Agronomic and physiological traits, and associated quantitative trait loci (QTL) affecting yield response in wheat (Triticum aestivum L.): A review. Front. Plant Sci. 2019, 10, 1428. [Google Scholar] [CrossRef] [Green Version]
- Chapman, S.; Mathews, K.; Trethowan, R.; Singh, R. Relationships between height and yield in near-isogenic spring wheats that contrast for major reduced height genes. Euphytica 2006, 157, 391–397. [Google Scholar] [CrossRef]
- Rebetzke, G.; Bonnett, D.; Ellis, M. Combining gibberellic acid-sensitive and insensitive dwarfing genes in breeding of higher-yielding, sesqui-dwarf wheats. Field Crop. Res. 2012, 127, 17–25. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, J.; Hu, Y.; Chen, J. Dwarfing genes Rht4 and Rht-B1b affect plant height and key agronomic traits in common wheat under two water regimes. Field Crop. Res. 2017, 204, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Chernook, A.; Kroupin, P.; Bespalova, L.; Panchenko, V.; Kovtunenko, V.; Bazhenov, M.; Nazarova, L.; Karlov, G.; Kroupina, A.; Divashuk, M. Phenotypic effects of the dwarfing gene Rht-17 in spring durum wheat under two climatic conditions. Vavilov J. Genet. Breed. 2019, 23, 916–925. [Google Scholar] [CrossRef]
- Li, S.; Tian, Y.; Wu, K.; Ye, Y.; Yu, J.; Zhang, J.; Liu, Q.; Hu, M.; Li, H.; Tong, Y.; et al. Modulating plant growth–metabolism coordination for sustainable agriculture. Nature 2018, 560, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Kroupin, P.; Chernook, A.; Bazhenov, M.; Karlov, G.; Goncharov, N.; Chikida, N.; Divashuk, M. Allele mining of TaGRF-2D gene 5′-UTR in Triticum aestivum and Aegilops tauschii genotypes. PLoS ONE 2020, 15, e0231704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drouyer, G.J.-P.; Bonnett, D.G.; Ellis, M.H.; Sirault, X.R.R. Unravelling the Effects of GA-Responsive Dwarfing Gene Rht13 on Yield and Grain Size. Available online: https://ses.library.usyd.edu.au/bitstream/handle/2123/3439/P306.pdf?sequence=1 (accessed on 10 June 2020).






{kind=link}
{kind=link}
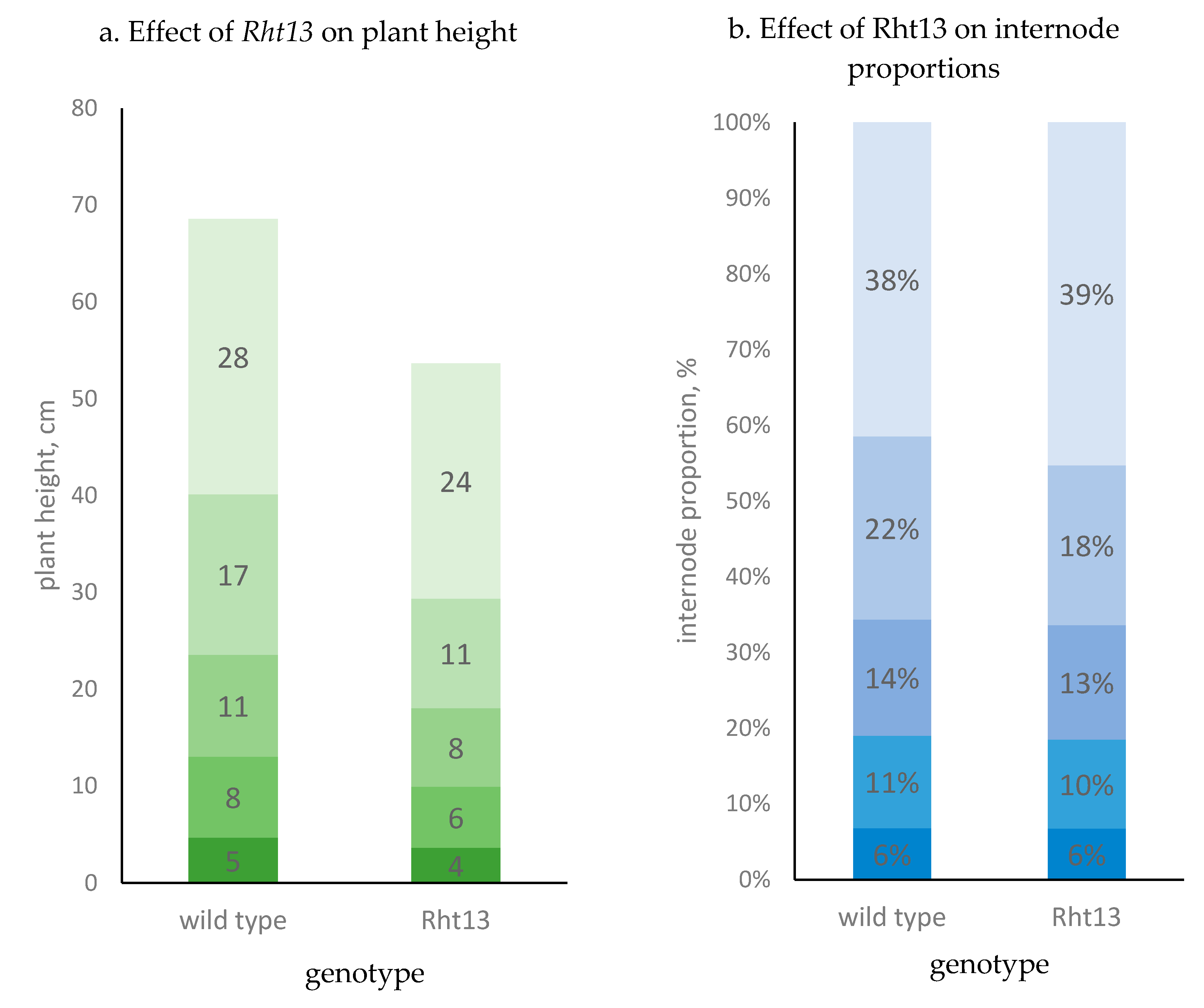{kind=link}
{kind=link}
{kind=link}
| Trait | Allele | Mean ± Standard Deviation | Contrast Rht13Rht13 vs. rht13rht13 (%) |
|---|---|---|---|
| Plant height (PH), cm | rht13 | 74.9 ± 6.0 | −13.0 (−17.4%) * |
| Rht13 | 61.9 ± 4.6 | ||
| 1st upper internode (peduncle), cm | rht13 | 28.4 ± 4.2 | −4.1 (−14.4%) * |
| Rht13 | 24.3 ± 3.5 | ||
| 2nd upper internode, cm | rht13 | 16.6 ± 2.1 | −5.3 (−31.9%) * |
| Rht13 | 11.3 ± 1.7 | ||
| 3rd upper internode, cm | rht13 | 10.5 ± 1.1 | −2.4 (−22.9%) * |
| Rht13 | 8.1 ± 1.3 | ||
| 2nd lower internode, cm | rht13 | 8.4 ± 1.5 | −2.1 (−25.0%) * |
| Rht13 | 6.3 ± 1.3 | ||
| 1st lower internode, cm | rht13 | 4.7 ± 1.9 | −1.1 (23.4%) * |
| Rht13 | 3.6 ± 1.5 | ||
| Internode number | rht13 | 4.7 ± 0.5a | 0.2 (4.3%) |
| Rht13 | 4.9 ± 0.4b | ||
| Grain number per main spike (GN), pcs. | rht13 | 28.9 ± 8.0 | 1.3 (4.5%) |
| Rht13 | 30.2 ± 7.9 | ||
| Grain number per spikelet (GNS), pcs. | rht13 | 1.9 ± 0.5 | 0.1 (5.3%) |
| Rht13 | 2.0 ± 0.5 | ||
| Main spike length (MSL), cm | rht13 | 9.0 ± 1.0 | 0.2 (2.2%) |
| Rht13 | 9.2 ± 1.0 | ||
| Spikelet number per spike (SN), pcs. | rht13 | 15.2 ± 1.4 | −0.2 (−1.3%) |
| Rht13 | 15.0 ± 1.3 | ||
| Spike compactness (SC) | rht13 | 16.9 ± 1.4 | −0.4 (−2.5%) |
| Rht13 | 16.5 ± 1.4 | ||
| Thousand grain weight (TGW), g | rht13 | 52.1 ± 3.6 | −5.1 (−9.8%) * |
| Rht13 | 47.0 ± 4.8 | ||
| Grain weight per main spike (GWS), g | rht13 | 1.5 ± 0.4 | −0.1 (−6.7%) * |
| Rht13 | 1.4 ± 0.4 | ||
| Number of fertile tillers per plant (FT), pcs. | rht13 | 2.5 ± 1.3 | 0.4 (16.0%) * |
| Rht13 | 2.9 ± 1.5 | ||
| Grain weight in side tillers (GWT), g | rht13 | 1.87 ± 1.14 | 0.01 (0.5%) |
| Rht13 | 1.86 ± 1.20 | ||
| Grain weight per shoot (GWSH), g | rht13 | 1.18 ± 0.42 | −0.13 (11.0%) * |
| Rht13 | 1.05 ± 0.35 | ||
| Main culm weight (MCW), g | rht13 | 0.74 ± 0.17 | −0.12 (−16.2%) * |
| Rht13 | 0.62 ± 0.15 | ||
| Main shoot biomass (BM), g | rht13 | 2.6 ± 0.6 | −0.2 (−7.7%) * |
| Rht13 | 2.4 ± 0.6 | ||
| Harvest index (HI) | rht13 | 0.556 ± 0.085 | 0.016 (2.9%) * |
| Rht13 | 0.572 ± 0.082 | ||
| Days to 80%-heading | rht13 | 50.8 ± 1.9 | −1.5 (−3.0%) |
| Rht13 | 49.3 ± 2.7 | ||
| Days to 80%-anthesis | rht13 | 52.1 ± 1.8 | −1.6 (−3.1%) |
| Rht13 | 50.5 ± 2.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Divashuk, M.G.; Kroupin, P.Y.; Shirnin, S.Y.; Vukovic, M.; Kroupina, A.Y.; Karlov, G.I. Effect of Gibberellin Responsive Reduced Height Allele Rht13 on Agronomic Traits in Spring Bread Wheat in Field Experiment in Non-Black Soil Zone. Agronomy 2020, 10, 927. https://doi.org/10.3390/agronomy10070927
Divashuk MG, Kroupin PY, Shirnin SY, Vukovic M, Kroupina AY, Karlov GI. Effect of Gibberellin Responsive Reduced Height Allele Rht13 on Agronomic Traits in Spring Bread Wheat in Field Experiment in Non-Black Soil Zone. Agronomy. 2020; 10(7):927. https://doi.org/10.3390/agronomy10070927
Chicago/Turabian StyleDivashuk, Mikhail G., Pavel Yu. Kroupin, Sergey Yu. Shirnin, Milena Vukovic, Aleksandra Yu. Kroupina, and Gennady I. Karlov. 2020. "Effect of Gibberellin Responsive Reduced Height Allele Rht13 on Agronomic Traits in Spring Bread Wheat in Field Experiment in Non-Black Soil Zone" Agronomy 10, no. 7: 927. https://doi.org/10.3390/agronomy10070927
APA StyleDivashuk, M. G., Kroupin, P. Y., Shirnin, S. Y., Vukovic, M., Kroupina, A. Y., & Karlov, G. I. (2020). Effect of Gibberellin Responsive Reduced Height Allele Rht13 on Agronomic Traits in Spring Bread Wheat in Field Experiment in Non-Black Soil Zone. Agronomy, 10(7), 927. https://doi.org/10.3390/agronomy10070927