Potassium Positively Affects Skin Characteristics of Sweet Potato Storage Roots

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Soil, Growth Management, and Fertilization Treatments
2.2. Skin Anatomical Study
2.3. RNA Extraction and Quantitative Real-Time PCR (qPCR)
2.4. Data Analysis
3. Results
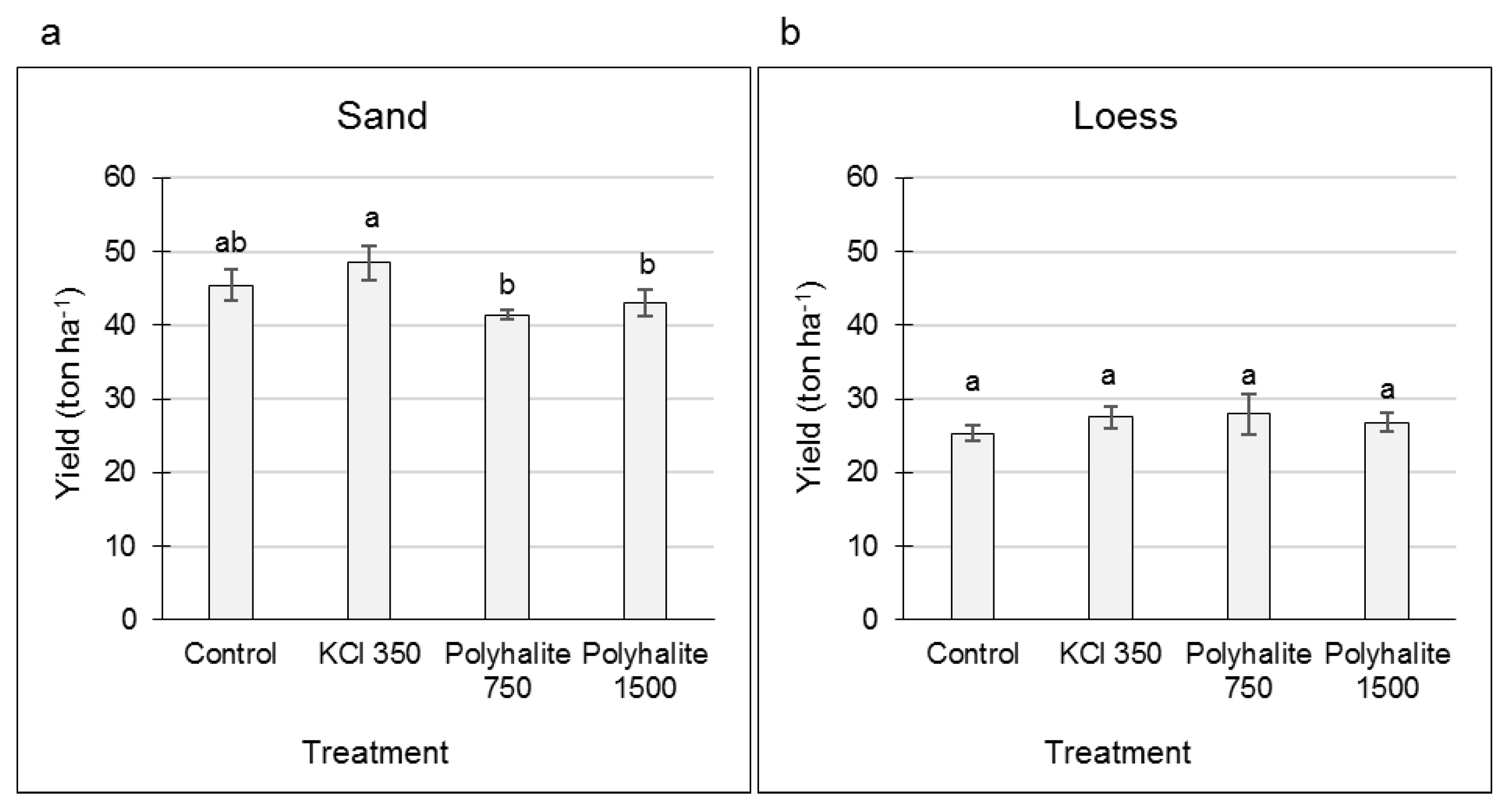
3.1. Effects of KCl and Polyhalite Fertilizers on SR Yield
3.2. Effects of KCl and Polyhalite Fertilizers on SR Skin Anatomy
3.3. Expression of Skin Molecular Markers and Effect of K Fertilizers
4. Discussion
4.1. Effects of K Fertilization on Yield
4.2. SR Uniformity
4.3. K Fertilization Affects Skin Characteristics
4.4. Expression of Skin-Related Genes
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Firon, N.; LaBonte, D.; Villordon, A.; McGregor, C.; Kfir, Y.; Pressman, E. Botany and physiology: Storage root formation and development. In The Sweetpotato; Loebenstein, G., Thottappilly, G., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 13–26. [Google Scholar] [CrossRef]
- Ma, J.; Aloni, R.; Villordon, A.; Labonte, D.; Kfir, Y.; Zemach, H.; Schwartz, A.; Althan, L.; Firon, N. Adventitious root primordia formation and development in stem nodes of ‘Georgia Jet’ sweetpotato, Ipomoea batatas. Am. J. Bot. 2015, 102, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Villordon, A.; LaBonte, D.; Solis, J.; Firon, N. Characterization of lateral root development at the onset of storage root initiation in ‘Beauregard’ sweetpotato adventitious roots. Hortscience 2012, 47, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Afek, U.; Orenstein, J.; Nuriel, E. Increased quality and prolonged storage of sweet potatoes in Israel. Phytoparasitica 1998, 26, 307–312. [Google Scholar] [CrossRef]
- Reeve, R.M.; Hautala, E.; Weaver, M.L. Anatomy and compositional variation within potatoes 1. Developmental histology of the tuber. Am. Potato J. 1969, 46, 361–373. [Google Scholar] [CrossRef]
- Lulai, E.C.; Freeman, T.P. The importance of phellogen cells and their structural characteristics in susceptibility and resistance to excoriation in immature and mature potato tuber (Solanum tuberosum L.) periderm. Ann. Bot. 2001, 88, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Bernards, M.A.; Lewis, N.G. The macromolecular aromatic domain in suberized tissue: A changing paradigm. Phytochemistry 1998, 47, 915–933. [Google Scholar] [CrossRef]
- Yan, B.; Stark, R.E. Biosynthesis, molecular structure, and domain architecture of potato suberin: A C-13 NMR study using isotopically labeled precursors. J. Agric. Food Chem. 2000, 48, 3298–3304. [Google Scholar] [CrossRef]
- Graça, J. Suberin: The biopolyester at the frontier of plants. Front. Chem. 2015, 3, 62. [Google Scholar] [CrossRef]
- Graça, J.; Pereira, H. Suberin structure in potato periderm: Glycerol, long-chain monomers, and glyceryl and feruloyl dimers. J. Agric. Food Chem. 2000, 48, 5476–5483. [Google Scholar] [CrossRef]
- Vishwanath, S.J.; Delude, C.; Domergue, F.; Rowland, O. Suberin: Biosynthesis, regulation, and polymer assembly of a protective extracellular barrier. Plant Cell Rep. 2015, 34, 573–586. [Google Scholar] [CrossRef]
- Serra, O.; Soler, M.; Hohn, C.; Franke, R.; Schreiber, L.; Prat, S.; Molinas, M.; Figueras, M. Silencing of StKCS6 in potato periderm leads to reduced chain lengths of suberin and wax compounds and increased peridermal transpiration. J. Exp. Bot. 2009, 60, 697–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, O.; Soler, M.; Hohn, C.; Sauveplane, V.; Pinot, F.; Franke, R.; Schreiber, L.; Prat, S.; Molinas, M.; Figueras, M. CYP86A33-targeted gene silencing in potato tuber alters suberin composition, distorts suberin lamellae, and impairs the periderm’s water barrier function. Plant Physiol. 2009, 149, 1050–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, O.; Hohn, C.; Franke, R.; Prat, S.; Molinas, M.; Figueras, M. A feruloyl transferase involved in the biosynthesis of suberin and suberin-associated wax is required for maturation and sealing properties of potato periderm. Plant J. 2010, 62, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Vulavala, V.K.; Fogelman, E.; Faigenboim, A.; Shoseyov, O.; Ginzberg, I. The transcriptome of potato tuber phellogen reveals cellular functions of cork cambium and genes involved in periderm formation and maturation. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vulavala, V.K.; Fogelman, E.; Rozental, L.; Faigenboim, A.; Tanami, Z.; Shoseyov, O.; Ginzberg, I. Identification of genes related to skin development in potato. Plant Mol. Biol. 2017, 94, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Ginzberg, I.; Minz, D.; Faingold, I.; Soriano, S.; Mints, M.; Fogelman, E.; Warshavsky, S.; Zig, U.; Yermiyahu, U. Calcium mitigated potato skin physiological disorder. Am. J. Potato Res. 2012, 89, 351–362. [Google Scholar] [CrossRef]
- Keren-Keiserman, A.; Baghel, R.S.; Fogelman, E.; Faingold, I.; Zig, U.; Yermiyahu, U.; Ginzberg, I. Effects of polyhalite fertilization on skin quality of potato tuber. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Vulavala, V.K.; Elbaum, R.; Yermiyahu, U.; Fogelman, E.; Kumar, A.; Ginzberg, I. Silicon fertilization of potato: Expression of putative transporters and tuber skin quality. Planta 2016, 243, 217–229. [Google Scholar] [CrossRef]
- Olsson, K. Resistance to gangrene (Phoma exigua var.foveata) and dry rot (Fusarium solani var.coeruleum) in potato tubers. 1. The influence of pectin-bound magnesium and calcium. Potato Res. 1988, 31, 413–422. [Google Scholar] [CrossRef]
- Palta, J.P. Role of calcium in plant responses to stresses: Linking basic research to the solution of practical problems. HortScience 1996, 31, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, M.; Briggs, S.; Knox, J. Intercellular adhesion and cell separation in plants. Plantcell Environ. 2003, 26, 977–989. [Google Scholar] [CrossRef]
- Vincken, J.-P.; Borkhardt, B.; Bush, M.; Doeswijk-Voragen, C.; Dopico, B.; Labrador, E.; Lange, L.; McCann, M.; Morvan, C.; Muñoz, F. Remodelling pectin structure in potato. In Developments in Plant Genetics and Breeding; Elsevier: Amsterdam, The Netherlands, 2000; Volume 6, pp. 245–256. [Google Scholar]
- Zörb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture—Status and perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Haberman, A.; Dag, A.; Shtern, N.; Zipori, I.; Erel, R.; Ben-Gal, A.; Yermiyahu, U. Long-term impact of potassium fertilization on soil and productivity in intensive olive cultivation. Agronomy 2019, 9, 525. [Google Scholar] [CrossRef] [Green Version]
- Yermiyahu, U.; Zipori, I.; Faingold, I.; Yusopov, L.; Faust, N.; Bar-Tal, A. Polyhalite as a multi nutrient fertilizer—Potassium, magnesium, calcium and sulfate. Isr. J. Plant Sci. 2017, 64, 145–157. [Google Scholar]
- Penman, H.L. Natural evaporation from open water, bare soil and grass. Proc. R. Soc. Lond. Ser. A Math. Phys. Sci. 1948, 193, 120–145. [Google Scholar]
- Ruzin, S.E. Plant Microtechnique and Microscopy; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Johansen, D.A. Plant Microtechniques; McGraw-Hill Book Company, Inc.: New York, NY, USA, 1940. [Google Scholar]
- Ginzberg, I.; Barel, G.; Ophir, R.; Tzin, E.; Tanami, Z.; Muddarangappa, T.; de Jong, W.; Fogelman, E. Transcriptomic profiling of heat-stress response in potato periderm. J. Exp. Bot. 2009, 60, 4411–4421. [Google Scholar] [CrossRef]
- Chua, K.; Kays, S.J. Effect of soil oxygen concentration on sweet potato storage root induction and/or development. HortScience 1981, 16, 71–73. [Google Scholar]
- Byju, G.; George, J. Potassium nutrition of sweet potato. Adv. Hortic. Sci. 2005, 19, 221–239. [Google Scholar]
- Fageria, N.K. Absorption of magnesium and its influence on the uptake of phosphorus, potassium, and calcium by intact groundnut plants. Plant Soil 1974, 40, 313–320. [Google Scholar] [CrossRef]
- Villordon, A.Q.; Bonte, D.R.L.; Firon, N.; Kfir, Y.; Pressman, E.; Schwartz, A. Characterization of Adventitious Root Development in Sweetpotato. HortScience 2009, 44, 651. [Google Scholar] [CrossRef]
- Firon, N.; LaBonte, D.; Villordon, A.; Kfir, Y.; Solis, J.; Lapis, E.; Perlman, T.S.; Doron-Faigenboim, A.; Hetzroni, A.; Althan, L. Transcriptional profiling of sweetpotato (Ipomoea batatas) roots indicates down-regulation of lignin biosynthesis and up-regulation of starch biosynthesis at an early stage of storage root formation. BMC Genom. 2013, 14, 460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, L.; Lowe, S. The anatomy of the root system in West Indian sweet potato (Ipomoea batatas (L.) Lam.) cultivars. Ann. Bot. 1973, 37, 633–643. [Google Scholar] [CrossRef]
- Togari, Y. A study of tuberous root formation in sweet potato. Bull. Nat. Agric. Exp. Stn. 1950, 68, 1–96. [Google Scholar]
- Gajanayake, B.; Reddy, K.R.; Shankle, M.W.; Arancibia, R.A.; Villordon, A.O. Quantifying storage root initiation, growth, and developmental responses of sweetpotato to early season temperature. Agron. J. 2014, 106, 1795–1804. [Google Scholar] [CrossRef]
- Villordon, A.; LaBonte, D.; Firon, N.; Carey, E. Variation in nitrogen rate and local availability alter root architecture attributes at the onset of storage root initiation in ‘Beauregard’ sweetpotato. HortScience 2013, 48, 808–815. [Google Scholar] [CrossRef] [Green Version]
- Kellermeier, F.; Chardon, F.; Amtmann, A. Natural variation of arabidopsis root architecture reveals complementing adaptive strategies to potassium starvation. Plant Physiol. 2013, 161, 1421–1432. [Google Scholar] [CrossRef] [Green Version]
- Du, F.; Liu, H.; Yin, X.; Zhao, Q.; Shi, C. Potassium-mediated regulation of sucrose metabolism and storage root formation in sweet potato. Arch. Agron. Soil Sci. 2020. [Google Scholar] [CrossRef]
- Villavicencio, L.E.; Blankenship, S.M.; Yencho, G.C.; Thomas, J.F.; Raper, C.D. Temperature effect on skin adhesion, cell wall enzyme activity, lignin content, anthocyanins, growth parameters, and periderm histochemistry of sweetpotato. J. Am. Soc. Hortic. Sci. 2007, 132, 729–738. [Google Scholar] [CrossRef]
- Effendy, J.; La Bonte, D.R.; Baisakh, N. Identification and expression of skinning injury-responsive genes in sweetpotato. J. Am. Soc. Hortic. Sci. 2013, 138, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Davies, H.; Millard, P. Fractionation and distribution of calcium in sprouting and non-sprouting potato tubers. Ann. Bot. 1985, 56, 745–754. [Google Scholar]
- Subramanian, N.K.; White, P.J.; Broadley, M.R.; Ramsay, G. The three-dimensional distribution of minerals in potato tubers. Ann. Bot. 2011, 107, 681–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, N.L.; Hiller, L.K.; Mikitzel, L.J. The dependence of internal brown spot development upon calcium fertility in potato tubers. Potato Res. 1996, 39, 165–178. [Google Scholar] [CrossRef]
- Busse, J.S.; Palta, J.P. Investigating the in vivo calcium transport path to developing potato tuber using 45Ca: A new concept in potato tuber calcium nutrition. Physiol. Plant. 2006, 128, 313–323. [Google Scholar] [CrossRef]
- Habib, A.; Donnelly, D.J. Calcium translocation and accumulation into potato tubers. Potato Res. 2002, 45, 17–24. [Google Scholar] [CrossRef]
- Shomer, I.; Novacky, A.J.; Pike, S.M.; Yermiyahu, U.; Kinraide, T.B. Electrical potentials of plant cell walls in response to the ionic environment. Plant Physiol. 2003, 133, 411–422. [Google Scholar] [CrossRef] [Green Version]
- Lobato, M.C.; Daleo, G.R.; Andreu, A.B.; Olivieri, F.P. Cell wall reinforcement in the potato tuber periderm after crop treatment with potassium phosphite. Potato Res. 2018, 61, 19–29. [Google Scholar] [CrossRef]
- Beisson, F.; Li-Beisson, Y.; Pollard, M. Solving the puzzles of cutin and suberin polymer biosynthesis. Curr. Opin. Plant Biol. 2012, 15, 329–337. [Google Scholar] [CrossRef]
- Domergue, F.; Vishwanath, S.J.; Joubes, J.; Ono, J.; Lee, J.A.; Bourdon, M.; Alhattab, R.; Lowe, C.; Pascal, S.; Lessire, R.; et al. Three Arabidopsis fatty acyl-coenzyme a reductases, FAR1, FAR4, and FAR5, generate primary fatty alcohols associated with suberin deposition. Plant Physiol. 2010, 153, 1539–1554. [Google Scholar] [CrossRef] [Green Version]
- Lulai, E.C.; Orr, P.H. Techniques for detecting and measuring developmental and maturational changes in tuber native periderm. Am. Potato J. 1994, 71, 489–505. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Type | ||
|---|---|---|
| Sand | Loess | |
| Sand (%) | 83.2 ± 3.3 | 66.8 ± 3.5 |
| Silt (%) | 2.3 ± 0.8 | 9.9 ± 1.4 |
| Clay (%) | 14.5 ± 1.2 | 23.3 ± 2 |
| Electrical conductivity (dS m−1) * | 1.20 ± 0.27 | 2.03 ± 0.13 |
| pH * | 7.63 ± 0.04 | 7.63 ± 0.04 |
| Minerals (mg kg−1 dry soil) | ||
| N-NO3 * | 8.62 ± 3.91 | 35.32 ± 9.50 |
| P * | 0.50 ± 0.09 | 0.02 ± 00 |
| K * | 22.27 ± 4.19 | 26.24 ± 2.76 |
| Mg * | 7.45 ± 1.40 | 16.43 ± 0.44 |
| Ca * | 36.38 ± 8.83 | 83.80 ± 4.19 |
| Cl * | 16.94 ± 6.53 | 64.05 ± 24.50 |
| Exchangeable K * | 217.2 ± 32.6 | 514.1 ± 77.1 |
| Gene Code | Gene | Potato ID | TAIR ID | Sweet potato ID * | Forward primer 5′→3′ | Reverse primer 5′→3′ | Length (bp) |
|---|---|---|---|---|---|---|---|
| CYP86A33 | FATTY ACID OMEGA-HYDROXYLASE | Sotub06g032570 | AT5G58860 | itf04g28930.t1 | AAACGCATCGCTCTCAACTT | GCTAATGCCGCCTTGAGATA | 227 |
| FAR3 | FATTY ACYL-COA REDUCTASE 3 | Sotub06g031240 | AT4G33790 | itf14g11580.t1 | TTTGCAAGTATTTCCAGACAACA | CCCCCTTGAAGAATAAGTATGG | 101 |
| FHT | FERULOYL TRANSFERASE | Sotub03g018220 | AT5G41040 | itf02g04740.t1 | AATACCAAGCCTGATCCTGT | GATGCTTCTGTCCAGGAATG | 230 |
| GPAT3 | GLYCEROL-3-PHOSPHATE 2-O-ACYLTRANSFERASE 4 | Sotub01g032090 | AT1G01610 | itf09g20300.t1 | CGATTGAAAAGCCGATTGAT | GTTGATTCCGAGCATGAGGT | 183 |
| KSC6 | 3-KETOACYL-COA SYNTHASE 6 | Sotub02g029020 | AT1G68530 | itf13g22850.t1 | CTCATCGTCAACTGCAGCAT | CAAATCTCGGGCCAAATCTA | 153 |
| OSP | ORGAN-SPECIFIC PROTEIN S2-LIKE | Sotub07g011080 | AT2G48030 | itf10g24760.t1 | GAGTATTGGGACGCGATGAT | CTGCTGAACATCCTTGGC | 300 |
| PLD | PHOSPHOLIPASE D1A | itf10g15190.t1 | ATCGGAATCAGCAGTGATGG | ATGATGAGGCAAGCAGTGTG | 160 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klipcan, L.; van Oss, R.; Keren-Kieserman, A.; Yermiyahu, U.; Ginzberg, I. Potassium Positively Affects Skin Characteristics of Sweet Potato Storage Roots. Agronomy 2020, 10, 1385. https://doi.org/10.3390/agronomy10091385
Klipcan L, van Oss R, Keren-Kieserman A, Yermiyahu U, Ginzberg I. Potassium Positively Affects Skin Characteristics of Sweet Potato Storage Roots. Agronomy. 2020; 10(9):1385. https://doi.org/10.3390/agronomy10091385
Chicago/Turabian StyleKlipcan, Liron, Ruth van Oss, Alexandra Keren-Kieserman, Uri Yermiyahu, and Idit Ginzberg. 2020. "Potassium Positively Affects Skin Characteristics of Sweet Potato Storage Roots" Agronomy 10, no. 9: 1385. https://doi.org/10.3390/agronomy10091385
APA StyleKlipcan, L., van Oss, R., Keren-Kieserman, A., Yermiyahu, U., & Ginzberg, I. (2020). Potassium Positively Affects Skin Characteristics of Sweet Potato Storage Roots. Agronomy, 10(9), 1385. https://doi.org/10.3390/agronomy10091385