The Influence of UV on the Production of Free Terpenes in Vitis vinifera cv. Shiraz


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Vineyard Site
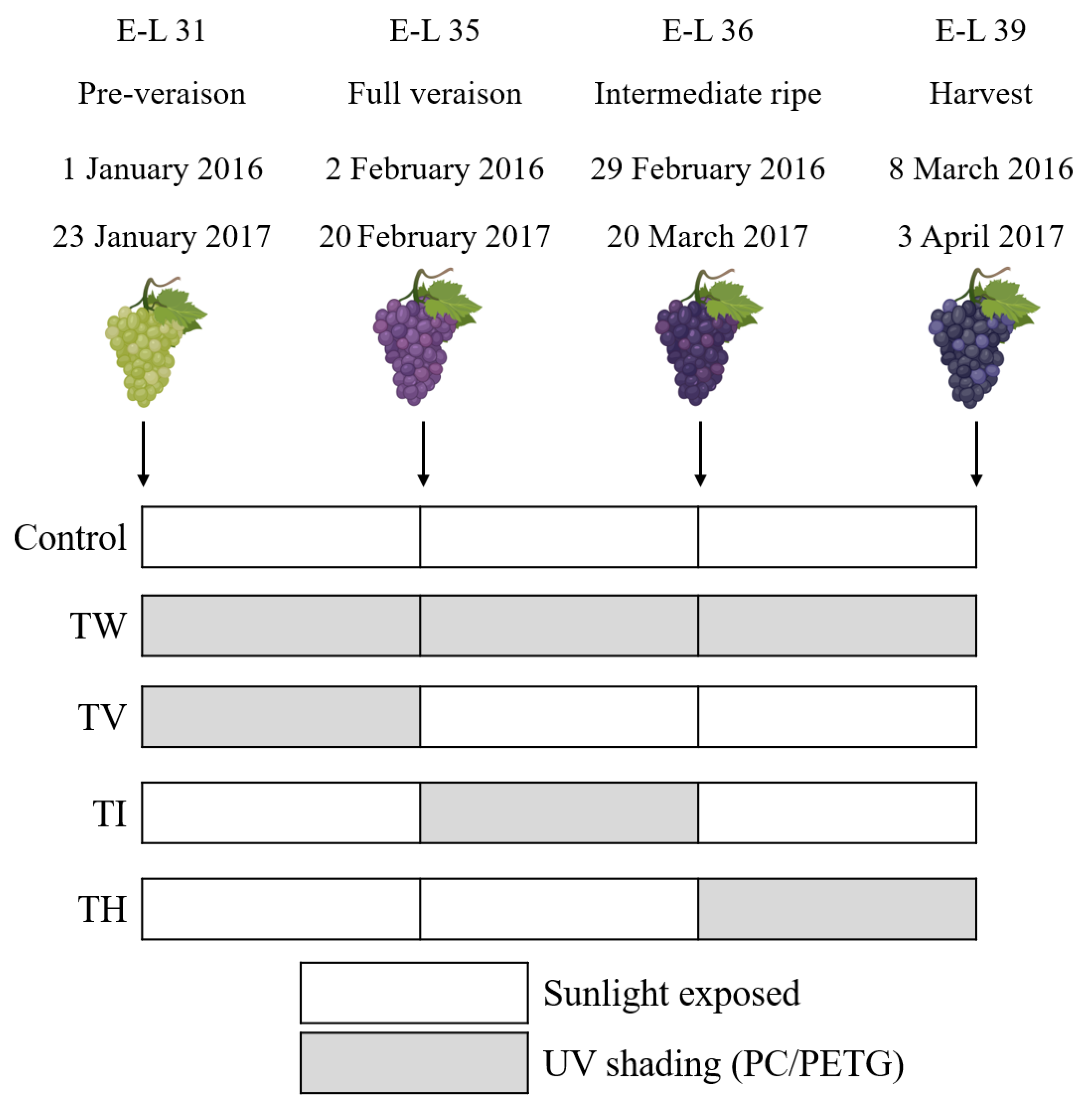
2.3. Experimental Design and Sampling
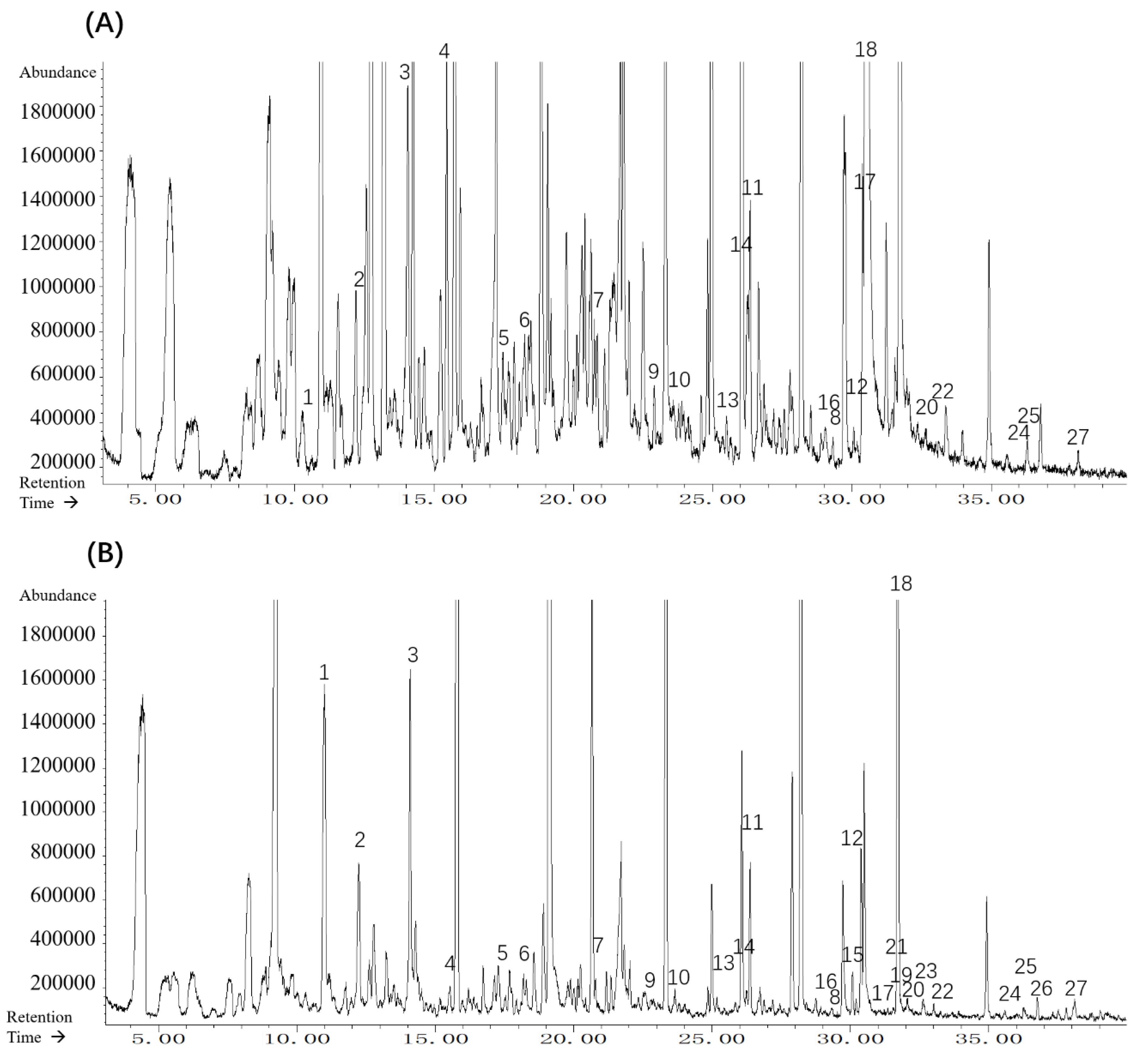
2.4. Headspace Solid Phase Microextraction Gas Chromatography–Mass Spectrometry (HS–SPME–GC–MS) Analysis
2.5. Statistical Analysis
3. Results and Discussion
3.1. Basic Chemical Parameters of the Shiraz Grapes
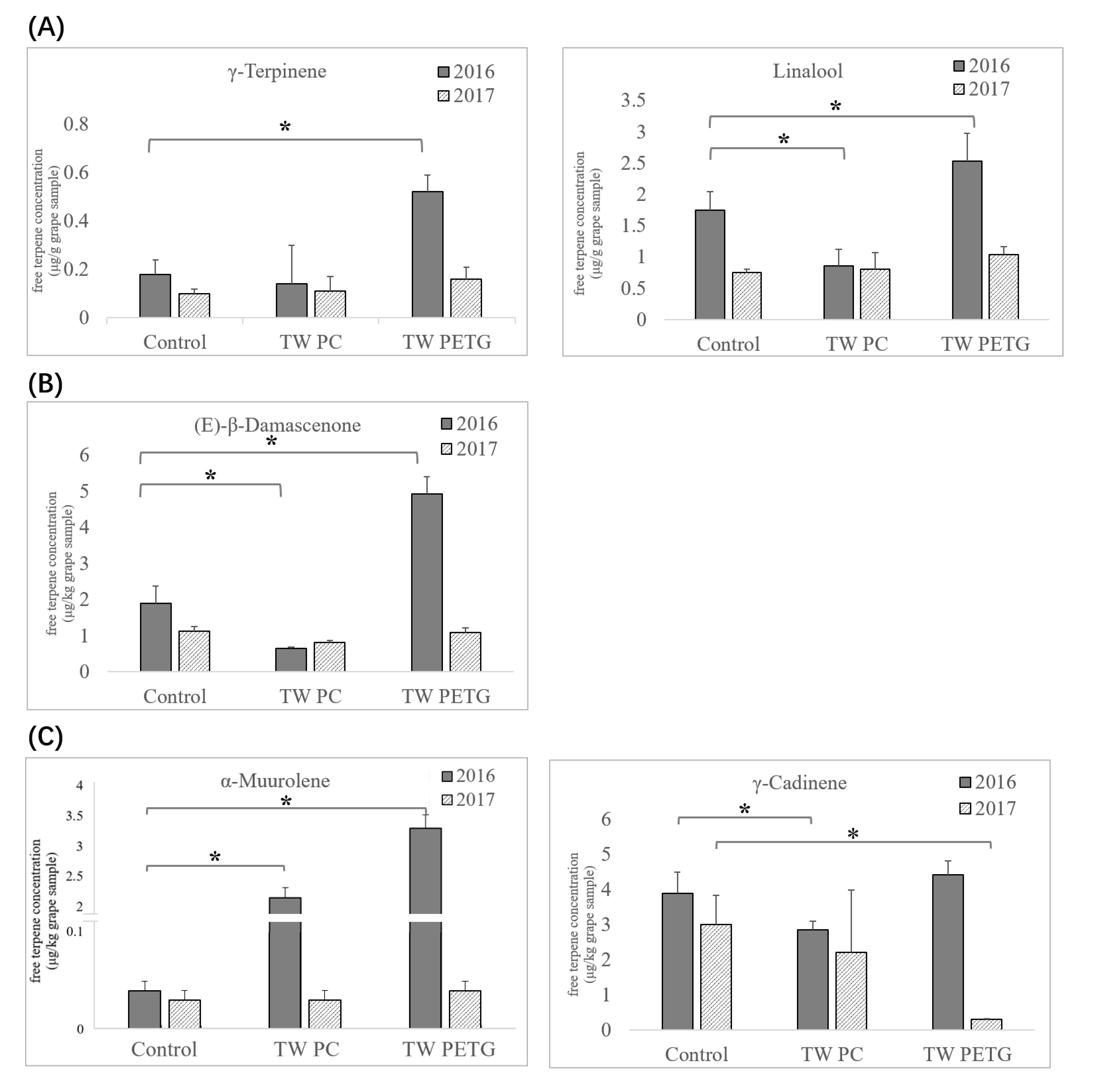
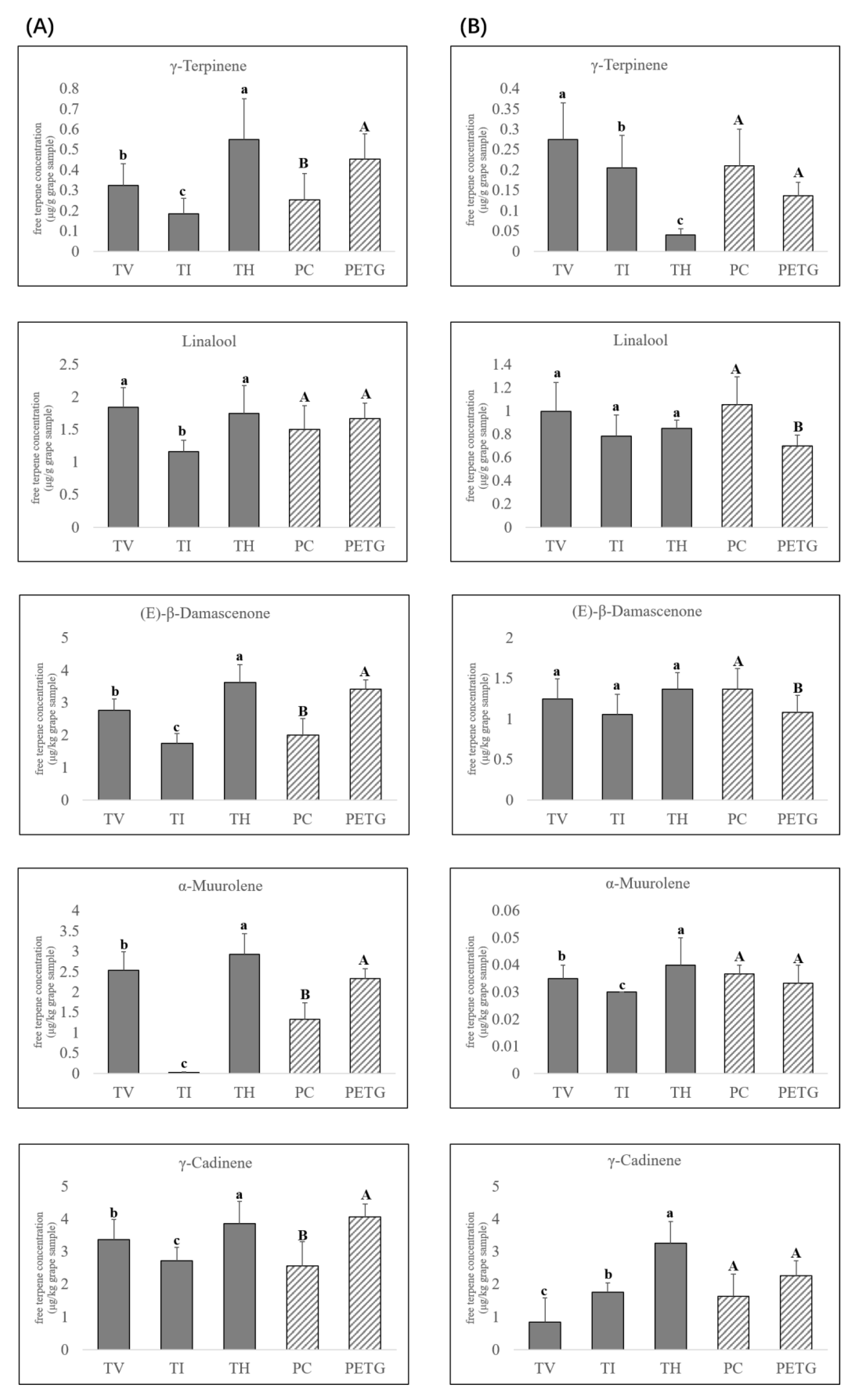
3.2. Response of Terpene Compounds to UV-Attenuated Treatments
3.2.1. Effects on the Concentrations of Free Monoterpene Compounds
3.2.2. Effects on the Concentrations of Free Norisoprenoid Compounds
3.2.3. Effects on the Concentrations of Sesquiterpene Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Howell, K.; Fang, Z.; Zhang, P. Sesquiterpenes in grapes and wines: Occurrence, biosynthesis, functionality, and influence of winemaking processes. Compr. Rev. Food Sci. Food Saf. 2020, 19, 247–281. [Google Scholar] [CrossRef]
- Black, C.A.; Parker, M.; Siebert, T.E.; Capone, D.L.; Francis, I.L. Terpenoids and their role in wine flavour: Recent advances. Aust. J. Grape Wine Res. 2015, 21, 582–600. [Google Scholar] [CrossRef]
- Vilanova, M.; Sieiro, C. Determination of free and bound terpene compounds in albariño wine. J. Food Compos. Anal. 2006, 19, 694–697. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Utsumi, R. Diversity, regulation, and genetic manipulation of plant mono- and sesquiterpenoid biosynthesis. Cell. Mol. Life Sci. 2009, 66, 3043–3052. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.S. 6-chemical constituents of grapes and wine. In Wine Science, 3rd ed.; Jackson, R.S., Ed.; Academic Press: San Diego, CA, USA, 2008; pp. 27–331. [Google Scholar]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial activity of terpenes and terpenoids present in essential oils. Molecules (Basel Switz.) 2019, 24, 2471. [Google Scholar] [CrossRef] [Green Version]
- Dahham, S.S.; Tabana, Y.M.; Iqbal, M.A.; Ahamed, M.B.; Ezzat, M.O.; Majid, A.S.; Majid, A.M. The anticancer, antioxidant and antimicrobial properties of the sesquiterpene β-caryophyllene from the essential oil of aquilaria crassna. Molecules (Basel Switz.) 2015, 20, 11808–11829. [Google Scholar] [CrossRef]
- González-Burgos, E.; Gómez-Serranillos, M.P. Terpene compounds in nature: A review of their potential antioxidant activity. Curr. Med. Chem. 2012, 19, 5319–5341. [Google Scholar] [CrossRef]
- Kalua, C.M.; Boss, P.K. Comparison of major volatile compounds from riesling and cabernet sauvignon grapes (Vitis vinifera L.) from fruitset to harvest. Aust. J. Grape Wine Res. 2010, 16, 337–348. [Google Scholar] [CrossRef]
- Coelho, E.; Rocha, S.M.; Delgadillo, I.; Coimbra, M.A. Headspace-spme applied to varietal volatile components evolution during Vitis vinifera L. Cv. ’Baga’ ripening. Anal. Chim. Acta 2006, 563, 204–214. [Google Scholar] [CrossRef]
- Zhang, P.; Howell, K.; Krstic, M.; Herderich, M.; Barlow, E.W.R.; Fuentes, S. Environmental factors and seasonality affect the concentration of rotundone in Vitis vinifera L. Cv. Shiraz wine. PLoS ONE 2015, 10, e0133137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, E.; Rocha, S.M.; Barros, A.S.; Delgadillo, I.; Coimbra, M.A. Screening of variety- and pre-fermentation-related volatile compounds during ripening of white grapes to define their evolution profile. Anal. Chim. Acta 2007, 597, 257–264. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.F.; Nieddu, G. Vine growth and physiological performance of two red grape cultivars under natural and reduced uv solar radiation. Aust. J. Grape Wine Res. 2016, 22, 105–114. [Google Scholar] [CrossRef]
- Gil, M.; Pontin, M.; Berli, F.; Bottini, R.; Piccoli, P. Metabolism of terpenes in the response of grape (Vitis vinifera L.) leaf tissues to uv-b radiation. Phytochemistry 2012, 77, 89–98. [Google Scholar] [CrossRef]
- Song, J.; Smart, R.; Wang, H.; Dambergs, B.; Sparrow, A.; Qian, M.C. Effect of grape bunch sunlight exposure and uv radiation on phenolics and volatile composition of Vitis vinifera L. Cv. Pinot noir wine. Food Chem. 2015, 173, 424–431. [Google Scholar] [CrossRef]
- Gregan, S.; Wargent, J.; Liu, L.; Shinkle, J.; Hofmann, R.; Winefield, C.; Trought, M.; Jordan, B. Effects of solar ultraviolet radiation and canopy manipulation on the biochemical composition of sauvignon blanc grapes. Aust. J. Grape Wine Res. 2012, 18, 227–238. [Google Scholar] [CrossRef]
- Šuklje, K.; Antalick, G.; Coetzee, Z.; Schmidtke, L.M.; Baša Česnik, H.; Brandt, J.; du Toit, W.J.; Lisjak, K.; Deloire, A. Effect of leaf removal and ultraviolet radiation on the composition and sensory perception of Vitis vinifera L. Cv. Sauvignon blanc wine. Aust. J. Grape Wine Res. 2014, 20, 223–233. [Google Scholar] [CrossRef]
- Gil, M.; Bottini, R.; Berli, F.; Pontin, M.; Silva, M.F.; Piccoli, P. Volatile organic compounds characterized from grapevine (Vitis vinifera L. Cv. Malbec) berries increase at pre-harvest and in response to uv-b radiation. Phytochemistry 2013, 96, 148–157. [Google Scholar] [CrossRef]
- Sasaki, K.; Takase, H.; Matsuyama, S.; Kobayashi, H.; Matsuo, H.; Ikoma, G.; Takata, R. Effect of light exposure on linalool biosynthesis and accumulation in grape berries. Biosci. Biotechno. Biochem. 2016, 80, 2376–2382. [Google Scholar] [CrossRef] [Green Version]
- Jug, T.; Rusjan, D. Advantages and disadvantages of uv-b radiations on grapevine (vitis sp.). Emir. J. Food Agric. 2012, 24, 576. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Cao, X.; Liu, X.; Xin, R.; Wang, J.; Gao, J.; Wu, B.; Gao, L.; Xu, C.; Zhang, B. Uv-b irradiation differentially regulates terpene synthases and terpene content of peach. Plant Cell Environ. 2017, 40, 2261–2275. [Google Scholar] [CrossRef]
- Coombe, B.G. Growth stages of the grapevine: Adoption of a system for identifying grapevine growth stages. Aust. J. Grape Wine Res. 1995, 1, 104–110. [Google Scholar] [CrossRef]
- Iland, P.; Bruer, N.; Edwards, G.; Weeks, S.; Wilkes, E. Chemical Analysis of Grapes and Wine: Techniques and Concepts; Patrick Iland Wine Promotions Pty Ltd: Adelaide, Australia, 2004. [Google Scholar]
- Zhang, P.; Fuentes, S.; Siebert, T.; Krstic, M.; Herderich, M.; Barlow, E.W.R.; Howell, K. Terpene evolution during the development of Vitis vinifera L. Cv. Shiraz grapes. Food Chem. 2016, 204, 463–474. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Brotchie, J.; Pang, M.; Marriott, P.J.; Howell, K.; Zhang, P. Dataset of concentrations of free terpenes at different phenological stages in Vitis vinifera L. Shiraz, cabernet sauvignon, riesling, chardonnay and pinot gris. Data Brief 2019, 27, 104595. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Brotchie, J.; Pang, M.; Marriott, P.J.; Howell, K.; Zhang, P. Free terpene evolution during the berry maturation of five Vitis vinifera L. Cultivars. Food Chem. 2019, 299, 125101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Fan, P.; Liu, C.; Wu, B.; Li, S.; Liang, Z. Sunlight exclusion from muscat grape alters volatile profiles during berry development. Food Chem. 2014, 164, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.; Chai, F.; Zhang, H.; Li, S.; Liang, Z.; Fan, P. Effects of sunlight exclusion on the profiles of monoterpene biosynthesis and accumulation in grape exocarp and mesocarp. Food Chem. 2017, 237, 379–389. [Google Scholar] [CrossRef]
- Berli, F.; Fanzone, M.; Piccoli, P.; Bottini, R. Solar uv-b and aba are involved in phenol metabolism of Vitis vinifera L. Increasing biosynthesis of berry skin polyphenols. J. Agric. Food Chem. 2011, 59, 4874–4884. [Google Scholar] [CrossRef]
- Lücker, J.; Bowen, P.; Bohlmann, J. Vitis vinifera terpenoid cyclases: Functional identification of two sesquiterpene synthase cdnas encoding (+)-valencene synthase and (−)-germacrene d synthase and expression of mono- and sesquiterpene synthases in grapevine flowers and berries. Phytochemistry 2004, 65, 2649–2659. [Google Scholar] [CrossRef]
- Bönisch, F.; Frotscher, J.; Stanitzek, S.; Rühl, E.; Bitz, O.; Schwab, W.; Wüst, M. Enantioselectivities of uridine diphosphate-glucose:Monoterpenol glucosyltransferases from grapevine vitis vinifera l.). In Importance of Chirality to Flavor Compounds; American Chemical Society: Washington, DC, USA, 2015; Volume 1212, pp. 77–83. [Google Scholar]
- Liu, D.; Gao, Y.; Li, X.X.; Li, Z.; Pan, Q.H. Attenuated uv radiation alters volatile profile in cabernet sauvignon grapes under field conditions. Molecules 2015, 20, 16946–16969. [Google Scholar] [CrossRef] [Green Version]
- Lafontaine, M.; Schultz, H.R.; Lopes, C.; Bálo, B.; Varadi, G. Leaf and fruit responses of riesling grapevines to uv-radiation in the field. In Proceedings of the Seventh International Symposium on Grapevine Physiology and Biotechnology, Davis, CA, USA, 21–25 June 2004. [Google Scholar]
- Bureau, S.M.; Baumes, R.L.; Razungles, A.J. Effects of vine or bunch shading on the glycosylated flavor precursors in grapes of Vitis vinifera L. Cv. Syrah. J. Agric. Food Chem. 2000, 48, 1290–1297. [Google Scholar] [CrossRef] [PubMed]
- Kwasniewski, M.T.; Vanden Heuvel, J.E.; Pan, B.S.; Sacks, G.L. Timing of cluster light environment manipulation during grape development affects C13 norisoprenoid and carotenoid concentrations in riesling. J. Agric. Food Chem. 2010, 58, 6841–6849. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, S.; Terrier, N.; Procureur, J.; Bigey, F.; Günata, Z. A carotenoid cleavage dioxygenase from Vitis vinifera L.: Functional characterization and expression during grape berry development in relation to C13-norisoprenoid accumulation. J. Exp. Bot. 2005, 56, 2721–2731. [Google Scholar] [CrossRef]
- Zhang, P.; Fuentes, S.; Siebert, T.; Krstic, M.; Herderich, M.; Barlow, E.W.R.; Howell, K. Comparison data of common and abundant terpenes at different grape development stages in shiraz wine grapes. Data Brief 2016, 8, 1127–1136. [Google Scholar] [CrossRef]
- Bönisch, F.; Frotscher, J.; Stanitzek, S.; Rühl, E.; Wüst, M.; Bitz, O.; Schwab, W. Activity-based profiling of a physiologic aglycone library reveals sugar acceptor promiscuity of family 1 udp-glucosyltransferases from grape. Plant Physiol. 2014, 166, 23–39. [Google Scholar] [CrossRef] [Green Version]
- Gil, M.; Bottini, R.; Pontin, M.; Berli, F.J.; Salomon, M.V.; Piccoli, P. Solar uv-b radiation modifies the proportion of volatile organic compounds in flowers of field-grown grapevine (Vitis vinifera L.) cv. Malbec. Plant Growth Regul. 2014, 74, 193–197. [Google Scholar] [CrossRef]
- May, B.; Lange, B.M.; Wust, M. Biosynthesis of sesquiterpenes in grape berry exocarp of Vitis vinifera L.: Evidence for a transport of farnesyl diphosphate precursors from plastids to the cytosol. Phytochemistry 2013, 95, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P. The Influence of Climate Factors and Plant Physiological Responses on the Accumulation of Rotundone in Vitis Vinifera cv. Shiraz Grapevines. Ph.D. Thesis, The University of Melbourne, Melbourne, Victoria, Australia, 2015. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Total Soluble Solid (°Brix) | pH | Titratable Acid (g/L) | Total Phenolic (Absorbance Units Per Gram) |
|---|---|---|---|---|
| Control | 21.7 ± 0.7 abc | 3.93 ± 0.11 ab | 4.7 ± 0.3 cd | 1.20 ± 0.09 a |
| TH PC | 21.4 ± 1.1 ab | 4.03 ± 0.18 ab | 3.7 ± 0.8 a | 1.31 ± 0.16 abc |
| TH PETG | 22.7 ± 1.0 bc | 4.03 ± 0.11 ab | 3.7 ± 0.3 a | 1.32 ± 0.11 abc |
| TI PC | 21.9 ± 0.8 abc | 4.03 ± 0.09 ab | 4.0 ± 0.4 abc | 1.32 ± 0.16 abc |
| TI PETG | 21.6 ± 0.3 abc | 4.00 ± 0.18 ab | 4.5 ± 0.8 bcd | 1.48 ± 0.07 c |
| TV PC | 22.0 ± 1.7 abc | 3.97 ± 0.11 ab | 4.6 ± 0.6 bcd | 1.40 ± 0.16 bc |
| TV PETG | 20.9 ± 1.1 a | 3.88 ± 0.13 a | 4.9 ± 0.4 d | 1.32 ± 0.11 abc |
| TW PC | 22.9 ± 1.0 c | 4.08 ± 0.09 b | 4.2 ± 0.5 abcd | 1.29 ± 0.09 ab |
| TW PETG | 22.2 ± 0.7 abc | 3.99 ± 0.13 ab | 3.9 ± 0.4 ab | 1.31 ± 0.09 abc |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, W.; Luo, J.; Liu, J.; Howell, K.; Zhang, P. The Influence of UV on the Production of Free Terpenes in Vitis vinifera cv. Shiraz. Agronomy 2020, 10, 1431. https://doi.org/10.3390/agronomy10091431
Miao W, Luo J, Liu J, Howell K, Zhang P. The Influence of UV on the Production of Free Terpenes in Vitis vinifera cv. Shiraz. Agronomy. 2020; 10(9):1431. https://doi.org/10.3390/agronomy10091431
Chicago/Turabian StyleMiao, Wen, Jiaqiang Luo, Junda Liu, Kate Howell, and Pangzhen Zhang. 2020. "The Influence of UV on the Production of Free Terpenes in Vitis vinifera cv. Shiraz" Agronomy 10, no. 9: 1431. https://doi.org/10.3390/agronomy10091431
APA StyleMiao, W., Luo, J., Liu, J., Howell, K., & Zhang, P. (2020). The Influence of UV on the Production of Free Terpenes in Vitis vinifera cv. Shiraz. Agronomy, 10(9), 1431. https://doi.org/10.3390/agronomy10091431