Assessment of the Capacity of Beneficial Bacterial Inoculants to Enhance Canola (Brassica napus L.) Growth under Low Water Activity

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation Method
2.2. Assessment of Bacterial Isolates for Catalase Activity
2.3. Iron Solubilisation Assays
2.4. Phosphate Solubilisation Assays
2.5. Quantification of Indole-3-Acetic Acid (IAA) Production by Bacterial Isolates
2.6. Quantification of 1-Aminocyclopropane-1-Carboxylate (ACC) Deaminase Activity by Bacterial Isolates
2.7. Bacterial Identification Using 16S rRNA Gene Sequences
2.8. Screening of Bacterial Isolates and Canola Germination on Medium Containing Polyethylene Glycol (PEG) to Reduce Water Activity
2.9. Effect of Water Activity on Growth of Bacterial Isolates
2.10. Screening of Bacterial Isolates on Canola Plants in a Sterilised Sand-Based Medium
2.11. Screening of Bacterial Isolates on Canola Plants in Sterilised Clay Loam-Based Medium
2.12. Field-Based Assessment of PGPB Strains on Canola Growth
2.13. Statistical Analyses
3. Results
3.1. Biochemical Screening of Isolates for PGPB Traits
3.2. Strain Identification
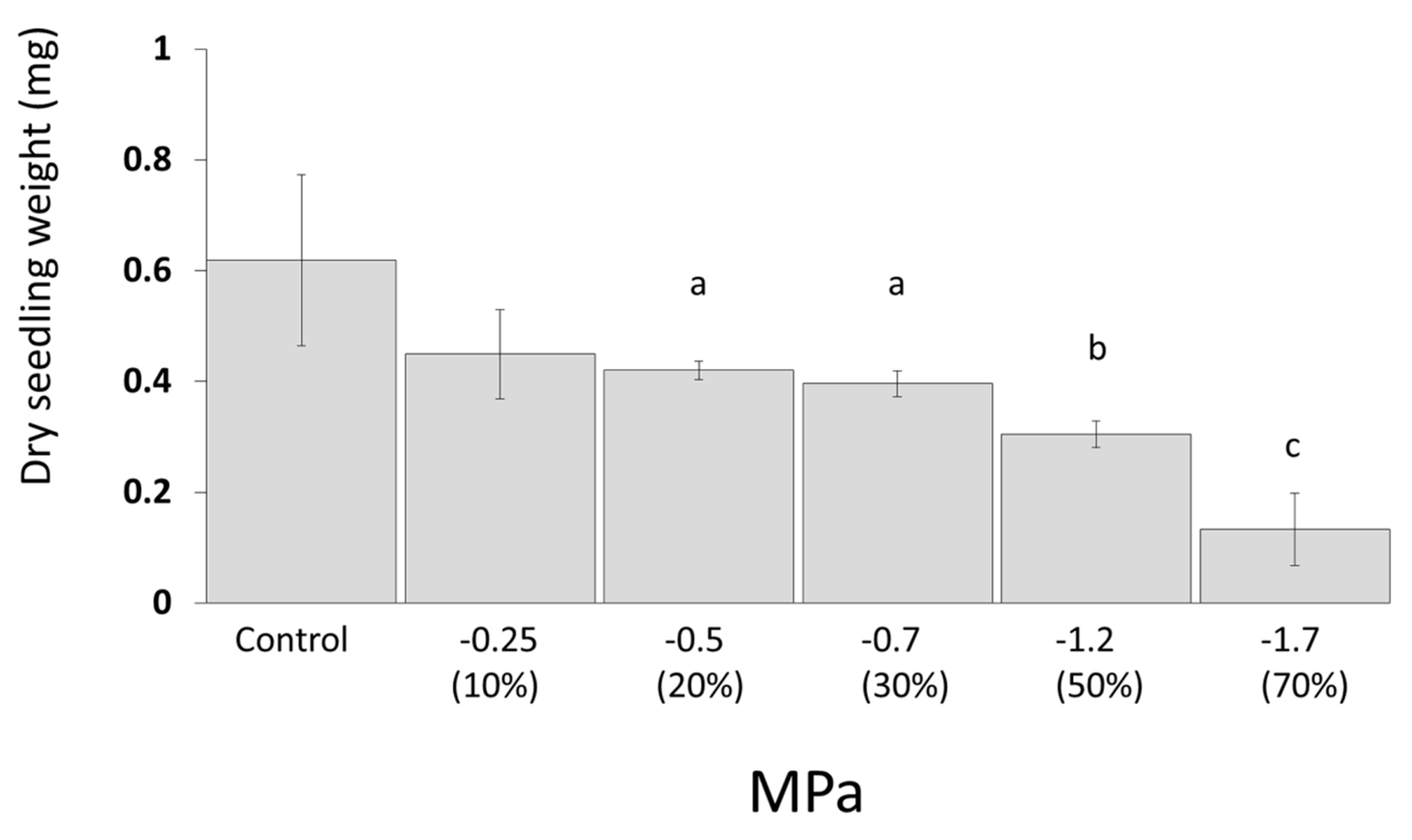
3.3. Desiccation Tolerance of Canola and Bacterial Strains
3.4. Growth Assessment in Sand-Based Medium
3.5. Effect of Bacterial Isolates on Canola Growth after 91 Days of Growth in a Sterile Clay Loam-Based Medium with Normal (8%) and Low (4%) Water Activity Levels
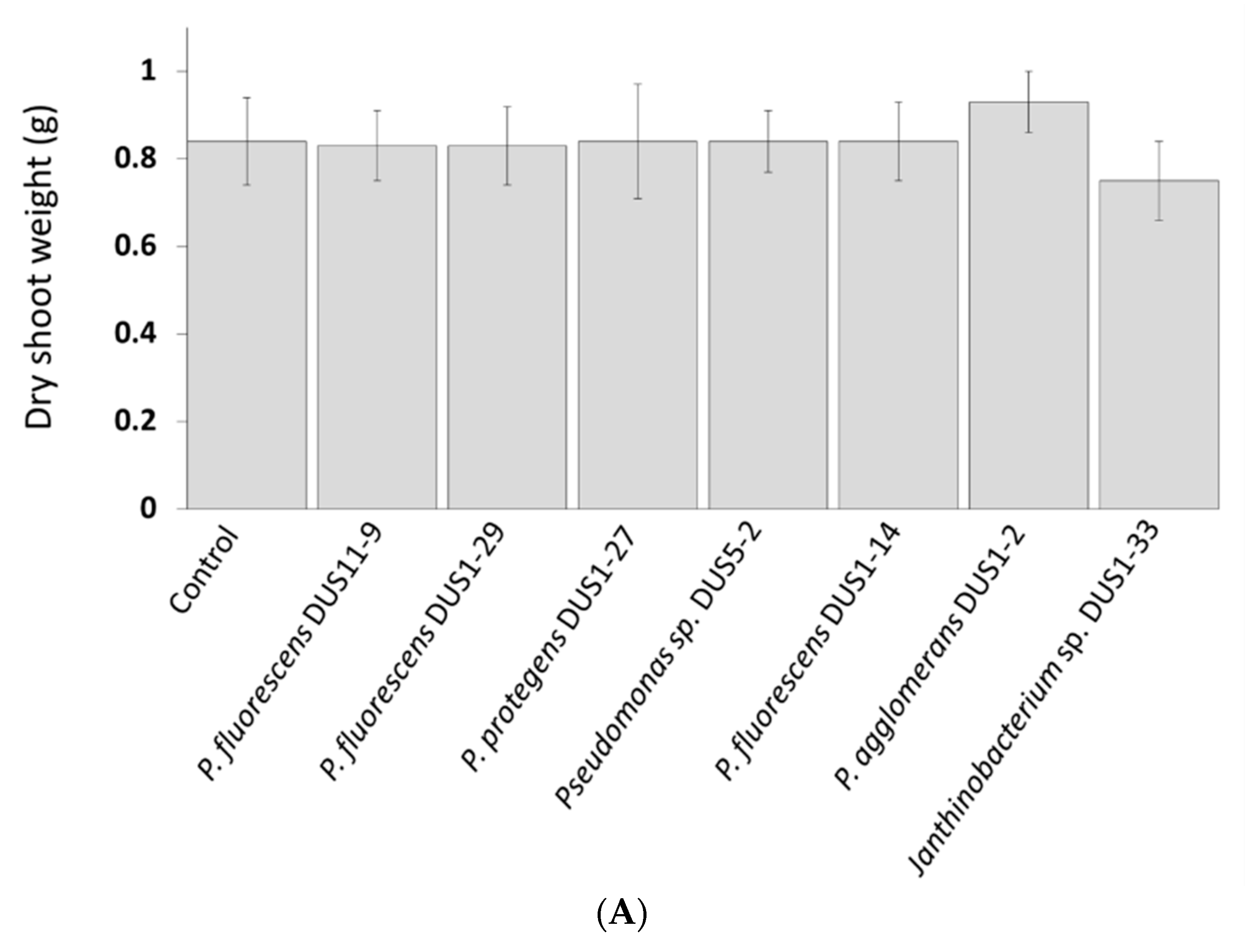
3.6. Assessment of Strain Performance on Enhancing B. napus L. Total Dry Plant Biomass in Sand or Soil Based Pot Systems with Either “Optimum” (3% Water Activity or Sand, or 8% Water Activity for Soil) or “Low” (1.5% Water Activity for Sand or 4% Water Activity for Soil) Water Activity
3.7. Field Assessment
3.8. Assessment of Strain Performance on Enhancing B. napus L. Total Dry Plant Biomass across All Plant Growth Systems Including Field Conditions
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Van de Wouw, A.; Idnurm, A.; Davidson, J.; Sprague, S.; Khangura, R.; Ware, A.; Lindbeck, K.; Marcroft, S. Fungal diseases of canola in Australia: Identification of trends, threats and potential therapies. Australas. Plant Pathol. 2016, 45, 415–423. [Google Scholar] [CrossRef]
- GRDC. GRDC Grownotes-Canola; Grain Research and Development Corporation Australia: Barton, Australia, 2018; pp. 1–8. [Google Scholar]
- Eady, S. Greenhouse Gas Emissions from the Cultivation of Canola Oilseed in Australia; CSIRO: Clayton, Australia, 2017. [Google Scholar]
- AOF. The Australian Oilseed Industry; Australian Oilseeds Federation: Sydney, Australia, 2010. [Google Scholar]
- Westwood, T. Oilseeds; The Australian Bureau of Agricultural and Resource Economics: Canberra, Australia, 2020; pp. 1–6. [Google Scholar]
- Faralli, M.; Grove, I.G.; Hare, M.C.; Kettlewell, P.S. In-field film antitranspirant application shows potential yield protection from flowering-stage drought periods in winter canola (Brassica napus). Crop. Pasture Sci. 2017, 68, 243–253. [Google Scholar] [CrossRef]
- Elferjani, R.; Soolanayakanahally, R. Canola responses to drought, heat, and combined stress: Shared and specific effects on carbon assimilation, seed yield, and oil composition. Front. Plant Sci. 2018, 9, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Channaoui, S.; El Idrissi, I.S.; Mazouz, H.; Nabloussi, A. Reaction of some rapeseed (Brassica napus L.) genotypes to different drought stress levels during germination and seedling growth stages. OCL 2019, 26, 23. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, N.H.; Møller, B.L.; Neale, A.D.; Hamill, J.D.; Blomstedt, C.K.; Gleadow, R.M. Effects of PEG-induced osmotic stress on growth and dhurrin levels of forage sorghum. Plant Physiol. Biochem. 2013, 73, 83–92. [Google Scholar] [CrossRef]
- Santos, M.S.; Nogueira, M.A.; Hungria, M. Microbial inoculants: Reviewing the past, discussing the present and previewing an outstanding future for the use of beneficial bacteria in agriculture. AMB Express 2019, 9, 205. [Google Scholar] [CrossRef]
- Alori, E.T.; Babalola, O.O. Microbial inoculants for improving crop quality and human health in Africa. Front. Microbiol. 2018, 9, 2213. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Cohen, A.C.; Bottini, R.; Pontin, M.; Berli, F.J.; Moreno, D.; Boccanlandro, H.; Travaglia, C.N.; Piccoli, P.N. Azospirillum brasilense ameliorates the response of Arabidopsis thaliana to drought mainly via enhancement of ABA levels. Physiol. Plant. 2015, 153, 79–90. [Google Scholar] [CrossRef]
- Wang, C.-J.; Yang, W.; Wang, C.; Gu, C.; Niu, D.-D.; Liu, H.-X.; Wang, Y.-P.; Guo, J.-H. Induction of drought tolerance in cucumber plants by a consortium of three plant growth-promoting rhizobacterium strains. PLoS ONE 2012, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.-H.; Kim, S.-D. Induction of drought stress resistance by multi-functional PGPR Bacillus licheniformis K11 in pepper. Plant Pathol. J. 2013, 29, 201. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Vardharajula, S.; Zulfikar Ali, S.; Grover, M.; Reddy, G.; Bandi, V. Drought-tolerant plant growth promoting Bacillus spp.: Effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Interact. 2011, 6, 1–14. [Google Scholar] [CrossRef]
- Kaushal, M.; Wani, S.P. Rhizobacterial-plant interactions: Strategies ensuring plant growth promotion under drought and salinity stress. Agric. Ecosyst. Environ. 2016, 231, 68–78. [Google Scholar] [CrossRef]
- Lobo, C.B.; Tomás, M.S.J.; Viruel, E.; Ferrero, M.A.; Lucca, M.E. Development of low-cost formulations of plant growth-promoting bacteria to be used as inoculants in beneficial agricultural technologies. Microbiol. Res. 2019, 219, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.Z.; Sandhya, V.; Grover, M.; Linga, V.R.; Bandi, V. Effect of inoculation with a thermotolerant plant growth promoting Pseudomonas putida strain AKMP7 on growth of wheat (Triticum spp.) under heat stress. J. Plant Interact. 2011, 6, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Xiong, L. Genetic engineering and breeding of drought-resistant crops. Annu. Rev. Plant Biol. 2014, 65, 715–741. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Clark, J.S.; Campbell, J.H.; Grizzle, H.; Acosta-Martìnez, V.; Zak, J.C. Soil microbial community response to drought and precipitation variability in the Chihuahuan Desert. Microb. Ecol. 2009, 57, 248–260. [Google Scholar] [CrossRef]
- Xue, R.; Shen, Y.; Marschner, P. Soil water content during and after plant growth influence nutrient availability and microbial biomass. J. Soil Sci. Plant Nutr. 2017, 17, 702–715. [Google Scholar] [CrossRef] [Green Version]
- Guntinas, M.; Gil-Sotres, F.; Leiros, M.; Trasar-Cepeda, C. Sensitivity of soil respiration to moisture and temperature. J. Soil Sci. Plant Nutr. 2013, 13, 445–461. [Google Scholar] [CrossRef] [Green Version]
- Papendick, R.; Campbell, G.S. Theory and Measurement of Water Potential; Soil Science Society of America: Madison, WI, USA, 1981; Volume 9, pp. 1–22. [Google Scholar]
- Harris, R. Effect of Water Potential on Microbial Growth and Activity; Soil Science Society of America: Madison, WI, USA, 1981; Volume 9, pp. 23–95. [Google Scholar]
- Filgueiras, L.; Silva, R.; Almeida, I.; Vidal, M.; Baldani, J.I.; Meneses, C.H.S.G. Gluconacetobacter diazotrophicus mitigates drought stress in Oryza sativa L. Plant Soil 2019, 451, 1–17. [Google Scholar] [CrossRef]
- Marasco, R.; Mosqueira, M.J.; Fusi, M.; Ramond, J.-B.; Merlino, G.; Booth, J.M.; Maggs-Kölling, G.; Cowan, D.A.; Daffonchio, D. Rhizosheath microbial community assembly of sympatric desert speargrasses is independent of the plant host. Microbiome 2018, 6, 215–225. [Google Scholar] [CrossRef]
- Jilani, G.; Akram, A.; Ali, R.M.; Hafeez, F.Y.; Shamsi, I.H.; Chaudhry, A.N.; Chaudhry, A.G. Enhancing crop growth, nutrients availability, economics and beneficial rhizosphere microflora through organic and biofertilizers. Ann. Microbiol. 2007, 57, 177–184. [Google Scholar] [CrossRef]
- Andrews, M.; James, E.; Cummings, S.; Zavalin, A.; Vinogradova, L.; McKenzie, B. Use of nitrogen fixing bacteria inoculants as a substitute for nitrogen fertiliser for dryland graminaceous crops: Progress made, mechanisms of action and future potential. Symbiosis 2003, 35, 209–229. [Google Scholar]
- Ren, X.; Guo, S.; Tian, W.; Chen, Y.; Han, H.; Chen, E.; Li, B.; Li, Y.; Chen, Z. Effects of plant growth-promoting bacteria (PGPB) inoculation on the growth, antioxidant activity, Cu uptake, and bacterial community structure of rape (Brassica napus L.) grown in Cu-contaminated agricultural soil. Front. Microbiol. 2019, 10, 1455. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Kamilova, F.; Validov, S.; Gafurova, L.; Kucharova, Z.; Lugtenberg, B. High incidence of plant growth stimulating bacteria associated with the rhizosphere of wheat grown on salinated soil in Uzbekistan. Environ. Microbiol. 2008, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; van der Voort, M.; Schneider, J.H.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.M.; de Leij, F. Rhizosphere; John Wiley & Sons, Ltd.: Chichester, UK, 2001. [Google Scholar] [CrossRef]
- Silva, R.; Filgueiras, L.; Santos, B.; Coelho, M.; Silva, M.; Estrada-Bonilla, G.; Vidal, M.; Baldani, J.I.; Meneses, C. Gluconacetobacter diazotrophicus changes the molecular mechanisms of root development in Oryza sativa L. growing under water stress. Int. J. Mol. Sci. 2020, 21, 333. [Google Scholar] [CrossRef] [Green Version]
- Duke, P.; Jarvis, J. The catalase test-a cautionary tale. J. Med. Lab. Technol. 1972, 29, 203–204. [Google Scholar]
- Forbes, B.A.; Sahm, D.F.; Weissfeld, A. Bailey and Scott’s Diagnostic Microbiology, 14th ed.; Elsevier-Health Sciences Division: St. Louis, MO, USA, 2007; pp. 779–800. [Google Scholar]
- MacFaddin, J.F. Biochemical Tests for Identification of Medical Bacteria, 3rd ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1976. [Google Scholar]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of blue agar CAS assay for siderophore detection. J. Microbiol. Biol. Educ. 2011, 12, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilising microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Mohite, B. Isolation and characterisation of indole acetic acid (IAA) producing bacteria from rhizospheric soil and its effect on plant growth. J. Soil Sci. Plant Nutr. 2013, 13, 638–649. [Google Scholar]
- Li, Z.; Chang, S.; Lin, L.; Li, Y.; An, Q. A colorimetric assay of 1-aminocyclopropane-1-carboxylate (ACC) based on ninhydrin reaction for rapid screening of bacteria containing ACC deaminase. Lett. Appl. Microbiol. 2011, 53, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Vílchez, J.I.; García-Fontana, C.; Román-Naranjo, D.; González-López, J.; Manzanera, M. Plant drought tolerance enhancement by trehalose production of desiccation-tolerant microorganisms. Front. Microbiol. 2016, 7, 1577. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil, 2nd ed.; College of Agriculture, University of California: Berkeley, CA, USA, 1950; Volume 347. [Google Scholar]
- Ding, W.; Shah, N.P. Effect of various encapsulating materials on the stability of probiotic bacteria. J. Food Sci. 2009, 74, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, 566–570. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloepper, J.W.; Lifshitz, R.; Zablotowicz, R.M. Free-living bacterial inocula for enhancing crop productivity. Trends Biotechnol. 1989, 7, 39–43. [Google Scholar] [CrossRef]
- Arruda, L.; Beneduzi, A.; Martins, A.; Lisboa, B.; Lopes, C.; Bertolo, F.; Passaglia, L.M.P.; Vargas, L.K. Screening of rhizobacteria isolated from maize (Zea mays L.) in Rio Grande do Sul State (South Brazil) and analysis of their potential to improve plant growth. Appl. Soil Ecol. 2013, 63, 15–22. [Google Scholar] [CrossRef]
- Trifi, H.; Salem, I.B.; Benzina, N.K.; Fourati, A.; Costa, M.C.; Achouak, W.; Sghaier, H.; Saidi, M. Effectiveness of the plant growth-promoting rhizobacterium Pantoea sp. BRM17 in enhancing Brassica napus growth in phosphogypsum-amended soil. Pedosphere 2017, 30, 570–576. [Google Scholar] [CrossRef]
- El-Howeity, M.; Asfour, M. Response of some varieties of canola plant (Brassica napus L.) cultivated in a newly reclaimed desert to plant growth promoting rhizobacteria and mineral nitrogen fertilizer. Ann. Agric. Sci. 2012, 57, 129–136. [Google Scholar] [CrossRef]
- Kirkham, M. Field Capacity, Wilting Point, Available Water, and the Non-Limiting Water Range, 2nd ed.; Acedemic Press: Cambridge, MA, USA, 2005. [Google Scholar] [CrossRef]
- Amellal, N.; Burtin, G.; Bartoli, F.; Heulin, T. Colonization of wheat roots by an exopolysaccharide-producing Pantoea agglomerans strain and its effect on rhizosphere soil aggregation. Appl. Environ. Microbiol. 1998, 64, 3740–3747. [Google Scholar] [CrossRef] [Green Version]
- Pera, A.; Vallini, G.; Sireno, I.; Bianchin, M.L.; De Bertoldi, M. Effect of organic matter on rhizosphere microorganisms and root development of sorghum plants in two different soils. Plant Soil 1983, 74, 3–18. [Google Scholar] [CrossRef]
- Glick, B.R.; Liu, C.; Ghosh, S.; Dumbroff, E.B. Early development of canola seedlings in the presence of the plant growth-promoting rhizobacterium Pseudomonas putida GR12-2. Soil Biol. Biochem. 1997, 29, 1233–1239. [Google Scholar] [CrossRef]
- Bertrand, H.; Nalin, R.; Bally, R.; Cleyet-Marel, J.-C. Isolation and identification of the most efficient plant growth-promoting bacteria associated with canola (Brassica napus). Biol. Fertil. Soils 2001, 33, 152–156. [Google Scholar] [CrossRef]
- Kloepper, J.; Hume, D.; Scher, F.; Singleton, C.; Tipping, B.; Laliberte, M.; Frauley, K.; Kutchaw, T.; Simonson, C.; Lifshitz, R. Plant growth-promoting rhizobacteria on canola (rapeseed). Plant Dis. 1988, 72, 42–46. [Google Scholar] [CrossRef]
- Ahmadi, K.; Zarebanadkouki, M.; Ahmed, M.A.; Ferrarini, A.; Kuzyakov, Y.; Kostka, S.J.; Carminati, A. Rhizosphere engineering: Innovative improvement of root environment. Rhizosphere 2017, 3, 176–184. [Google Scholar] [CrossRef]
- Santaella, C.; Schue, M.; Berge, O.; Heulin, T.; Achouak, W. The exopolysaccharide of Rhizobium sp. YAS34 is not necessary for biofilm formation on Arabidopsis thaliana and Brassica napus roots but contributes to root colonization. Environ. Microbiol. 2008, 10, 2150–2163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carminati, A.; Zarebanadkouki, M.; Kroener, E.; Ahmed, M.A.; Holz, M. Biophysical rhizosphere processes affecting root water uptake. Ann. Bot. 2016, 118, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Chenu, C. Extracellular Polysaccharides: An Interface between Microorganisms and Soil Constituents; CRC Lewis Publishers: Boca Raton, FL, USA, 1995; pp. 217–233. [Google Scholar]
- Pantanella, F.; Berlutti, F.; Passariello, C.; Sarli, S.; Morea, C.; Schippa, S. Violacein and biofilm production in Janthinobacterium lividum. J. Appl. Microbiol 2007, 102, 992–999. [Google Scholar] [CrossRef]
- Yu, X.; Jiang, J.; Liang, C.; Zhang, X.; Wang, J.; Shen, D.; Feng, Y. Indole affects the formation of multicellular aggregate structures in Pantoea agglomerans YS19. J. Gen. Appl. Microbiol. 2016, 62, 31–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, R.; Pal, K.; Bhatt, D.; Chauhan, S. Growth promotion and yield enhancement of peanut (Arachis hypogaea L.) by application of plant growth-promoting rhizobacteria. Microbiol. Res. 2004, 159, 371–394. [Google Scholar] [CrossRef]
- Hernlem, B.J.; Vane, L.M.; Sayles, G.D. The application of siderophores for metal recovery and waste remediation: Examination of correlations for prediction of metal affinities. Water Res. 1999, 33, 951–960. [Google Scholar] [CrossRef]
- Schalk, I.J.; Hannauer, M.; Braud, A. New roles for bacterial siderophores in metal transport and tolerance. Environ. Microbiol. 2011, 13, 2844–2854. [Google Scholar] [CrossRef]
- Rajkumar, M.; Ae, N.; Prasad, M.N.V.; Freitas, H. Potential of siderophore-producing bacteria for improving heavy metal phytoextraction. Trends Biotechnol. 2010, 28, 142–149. [Google Scholar] [CrossRef]
- Trapet, P.; Avoscan, L.; Klinguer, A.; Pateyron, S.; Citerne, S.; Chervin, C.; Mazurier, S.; Lemanceau, P.; Wendehenne, D.; Besson-Bard, A. The Pseudomonas fluorescens siderophore pyoverdine weakens Arabidopsis thaliana defense in favour of growth in iron-deficient conditions. Plant Physiol. 2016, 675–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganeshan, G.; Manoj Kumar, A. Pseudomonas fluorescens, a potential bacterial antagonist to control plant diseases. J. Plant Interact. 2005, 1, 123–134. [Google Scholar] [CrossRef]
- Aznar, A.; Dellagi, A. New insights into the role of siderophores as triggers of plant immunity: What can we learn from animals? J. Exp. Bot. 2015, 66, 3001–3010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiharn, M.; Chunhaleuchanon, S.; Lumyong, S. Screening siderophore producing bacteria as potential biological control agent for fungal rice pathogens in Thailand. World J. Microbiol. Biotechnol. 2009, 25, 1919–1928. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiharn, M.; Theantana, T.; Pathom-aree, W. Evaluation of Biocontrol Activities of Streptomyces spp. against Rice Blast Disease Fungi. Pathogens 2020, 9, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekaran, M.; Chun, S.C. Induction of defence-related enzymes in tomato (Solanum lycopersicum) plants treated with Bacillus subtilis CBR05 against Xanthomonas campestris pv. vesicatoria. Biocontrol Sci. Technol. 2016, 26, 1366–1378. [Google Scholar] [CrossRef]
- Shoaib, A.; Awan, Z.A.; Khan, K.A. Intervention of antagonistic bacteria as a potential inducer of disease resistance in tomato to mitigate early blight. Sci. Hortic. 2019, 252, 20–28. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Patten, C.L.; Glick, B.R. Bacterial biosynthesis of indole-3-acetic acid. Can. J. Microbiol. 1996, 42, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Prinsen, E.; Costacurta, A.; Michiels, K.; Vanderleyden, J.; Van Onckelen, H. Azospirillum brasilense indole-3-acetic acid biosynthesis: Evidence for a non-tryptophan dependent pathway. Mol. Plant Microbe Interact. 1993, 6, 609–615. [Google Scholar] [CrossRef]
- Jasim, B.; Joseph, A.A.; John, C.J.; Mathew, J.; Radhakrishnan, E. Isolation and characterization of plant growth promoting endophytic bacteria from the rhizome of Zingiber officinale. 3 Biotech 2014, 4, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Francis, C.A.; Obraztsova, A.Y.; Tebo, B.M. Dissimilatory metal reduction by the facultative anaerobe Pantoea agglomerans SP1. Appl. Environ. Microbiol. 2000, 66, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Sachdev, D.P.; Chaudhari, H.G.; Kasture, V.M.; Dhavale, D.D.; Chopade, B.A. Isolation and characterization of indole acetic acid (IAA) producing Klebsiella pneumoniae strains from rhizosphere of wheat (Triticum aestivum) and their effect on plant growth. Indian J. Exp. Biol. 2009, 47, 993–1000. [Google Scholar] [PubMed]
- Glick, B.R.; Karaturovíc, D.M.; Newell, P.C. A novel procedure for rapid isolation of plant growth promoting pseudomonads. Can. J. Microbiol. 1995, 41, 533–536. [Google Scholar] [CrossRef]
- Belimov, A.A.; Safronova, V.I.; Sergeyeva, T.A.; Egorova, T.N.; Matveyeva, V.A.; Tsyganov, V.E.; Borisov, A.Y.; Tikhonovich, I.A.; Kluge, C.; Preisfeld, A. Characterization of plant growth promoting rhizobacteria isolated from polluted soils and containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2001, 47, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Indiragandhi, P.; Anandham, R.; Madhaiyan, M.; Sa, T. Characterization of plant growth–promoting traits of bacteria isolated from larval guts of diamondback moth Plutella xylostella (Lepidoptera: Plutellidae). Curr. Microbiol. 2008, 56, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Siddikee, M.A.; Chauhan, P.; Anandham, R.; Han, G.-H.; Sa, T. Isolation, characterization, and use for plant growth promotion under salt stress, of ACC deaminase-producing halotolerant bacteria derived from coastal soil. J. Microbiol. Biotechnol. 2010, 20, 1577–1584. [Google Scholar] [CrossRef] [PubMed]
- Lay, C.-Y.; Bell, T.H.; Hamel, C.; Harker, K.N.; Mohr, R.; Greer, C.W.; Yergeau, É.; St-Arnaud, M. Canola root–associated microbiomes in the Canadian Prairies. Front. Microbiol. 2018, 9, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Poly, F.; Monrozier, L.J.; Bally, R. Improvement in RFLP procedure to study the community of nitrogen fixers in soil through the diversity of nifH gene. Res. Microbiol. 2001, 152, 95–103. [Google Scholar] [CrossRef]
- Gray, E.; Smith, D. Intracellular and extracellular PGPR: Commonalities and distinctions in the plant–bacterium signaling processes. Soil Biol. Biochem. 2005, 37, 395–412. [Google Scholar] [CrossRef]
- Farina, R.; Beneduzi, A.; Ambrosini, A.; de Campos, S.B.; Lisboa, B.B.; Wendisch, V.; Vargas, L.K.; Passaglia, L.M. Diversity of plant growth-promoting rhizobacteria communities associated with the stages of canola growth. Appl. Soil Ecol. 2012, 55, 44–52. [Google Scholar] [CrossRef]
- de Campos, S.B.; Youn, J.-W.; Farina, R.; Jaenicke, S.; Jünemann, S.; Szczepanowski, R.; Beneduzi, A.; Vargas, L.K.; Goesmann, A.; Wendisch, V.F. Changes in root bacterial communities associated to two different development stages of canola (Brassica napus L. var oleifera) evaluated through next-generation sequencing technology. Microbiol. Res. 2013, 65, 593–601. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate ID | Organism Name | Fe2+/3+ Solubilisation | PO43− Solubilisation | IAA μg/mg Protein | ACC Deaminase Activity (µM) Per mg of Protein |
|---|---|---|---|---|---|
| DUS11-9 | Pseudomonas fluorescens | + | + | 0.24 | 0.94 |
| DUS1-29 | Pseudomonas fluorescens | + | + | 0.25 | 1.12 |
| DUS1-27 | Pseudomonas protegens | + | + | 0.07 | 0.79 |
| DUS5-2 | Pseudomonas sp. | + | + | 0.29 | 0.45 |
| DUS1-14 | Pseudomonas fluorescens | + | + | 0.07 | 1.59 |
| DUS1-2 | Pantoea agglomerans | + | + | 0.84 | 1.33 |
| DUS1-33 | Janthinobacterium sp. | + | + | 2.41 | 0.84 |
| Strain Name | CFU × 104 Inoculated Strain | CFU × 102 Other Bacteria |
|---|---|---|
| Control (uninoculated) | N/A | 4 ± 1.0 |
| P. fluorescens DUS11-9 | 9.0 ± 1.0 | 3 ± 1.0 |
| P. fluorescens DUS1-29 | 9.0 ± 2.0 | 4 ± 1.0 |
| P. protegens DUS1-27 | 7.0 ± 1.0 | 4 ± 1.0 |
| Pseudomonas sp. DUS5-2 | 8.0 ± 1.0 | 4 ± 1.0 |
| P. fluorescens DUS1-14 | 9.0 ± 1.0 | 3 ± 1.0 |
| P. agglomerans DUS1-2 | 9.0 ± 1.0 | 3 ± 1.0 |
| Janthinobacterium sp. DUS1-33 | 7.0 ± 1.0 | 4 ± 1.0 |
| Strain Name | CFU × 103 Inoculated Strain | CFU × 102 Other Bacteria |
|---|---|---|
| Control (uninoculated) | N/A | 3 ± 1.0 |
| P. fluorescens DUS11-9 | 6.0 ± 1.0 | 3 ± 1.0 |
| P. fluorescens DUS1-29 | 7.0 ± 1.0 | 2 ± 1.0 |
| P. protegens DUS1-27 | 6.0 ± 1.0 | 3 ± 1.0 |
| Pseudomonas sp. DUS5-2 | 6.0 ± 1.0 | 2 ± 1.0 |
| P. fluorescens DUS1-14 | 6.0 ± 1.0 | 3 ± 1.0 |
| P. agglomerans DUS1-2 | 7.0 ± 1.0 | 2 ± 1.0 |
| Janthinobacterium sp. DUS1-33 | 5.0 ± 1.0 | 3 ± 1.0 |
| Strain Name | CFU × 104 Inoculated Strain | CFU × 104 Other Bacteria | Germination % (15 Seeds) |
|---|---|---|---|
| Control (uninoculated) | N/A | 53.6 ± 21.3 | 100 ± 0 |
| P. fluorescens DUS11-9 | 15.4 ± 9.7 | 19.4 ± 8.3 a | 100 ± 0 |
| P. fluorescens DUS1-29 | 39.0 ± 11.3 | 14.4 ± 8.5 a | 100 ± 0 |
| P. protegens DUS1-27 | 20.8 ± 8.3 | 28.2 ± 7.5 | 100 ± 0 |
| Pseudomonas sp. DUS5-2 | 20.2 ± 12.1 | 31.4 ± 10.4 | 100 ± 0 |
| P. fluorescens DUS1-14 | 24.4 ± 10.3 | 21.2 ± 10.3 | 98 ± 2 |
| P. agglomerans DUS1-2 | 22.4 ± 5.7 | 27.4 ± 10.3 | 100 ± 0 |
| Janthinobacterium sp. DUS1-33 | 39.4 ± 13.6 | 22.2 ± 8.4 | 98 ± 2 |
| Strain Name | CFU × 104 Inoculated Strain | CFU × 104 Other Bacteria | Germination % (15 Seeds) |
|---|---|---|---|
| Control (uninoculated) | N/A | 47.6 ± 16.4 | 97 ± 3 |
| P. fluorescens DUS11-9 | 29.4 ± 8.3 | 17.6 ± 6.8 b | 98 ± 2 |
| P. fluorescens DUS1-29 | 21.4 ± 9.2 | 15.2 ± 6.2 b | 98 ± 2 |
| P. protegens DUS1-27 | 61.2 ± 22.4 a | 11.6 ± 7.1 b | 98 ± 2 |
| Pseudomonas sp. DUS5-2 | 23.2 ± 7.2 | 20.6 ± 11.3 | 97 ± 3 |
| P. fluorescens DUS1-14 | 19.8 ± 8.3 | 23.2 ± 10.5 | 97 ± 3 |
| P. agglomerans DUS1-2 | 23.4 ± 8.6 | 18.6 ± 6.6 b | 100 ± 0 |
| Janthinobacterium sp. DUS1-33 | 23.2 ± 8.5 | 36.2 ± 8.3 | 97 ± 3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Premachandra, D.; Hudek, L.; Enez, A.; Ballard, R.; Barnett, S.; Franco, C.M.M.; Brau, L. Assessment of the Capacity of Beneficial Bacterial Inoculants to Enhance Canola (Brassica napus L.) Growth under Low Water Activity. Agronomy 2020, 10, 1449. https://doi.org/10.3390/agronomy10091449
Premachandra D, Hudek L, Enez A, Ballard R, Barnett S, Franco CMM, Brau L. Assessment of the Capacity of Beneficial Bacterial Inoculants to Enhance Canola (Brassica napus L.) Growth under Low Water Activity. Agronomy. 2020; 10(9):1449. https://doi.org/10.3390/agronomy10091449
Chicago/Turabian StylePremachandra, Dasun, Lee Hudek, Aydin Enez, Ross Ballard, Steve Barnett, Christopher M.M. Franco, and Lambert Brau. 2020. "Assessment of the Capacity of Beneficial Bacterial Inoculants to Enhance Canola (Brassica napus L.) Growth under Low Water Activity" Agronomy 10, no. 9: 1449. https://doi.org/10.3390/agronomy10091449
APA StylePremachandra, D., Hudek, L., Enez, A., Ballard, R., Barnett, S., Franco, C. M. M., & Brau, L. (2020). Assessment of the Capacity of Beneficial Bacterial Inoculants to Enhance Canola (Brassica napus L.) Growth under Low Water Activity. Agronomy, 10(9), 1449. https://doi.org/10.3390/agronomy10091449