Evaluation of Italian and Spanish Accessions of Brassica rapa L.: Effect of Flowering Earliness on Fresh Yield and Biological Value

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
2.3. Biomorphometric Traits
2.4. GLSs Identification and Quantification
2.5. Evaluation of Antioxidant Activity: ABTS Assay
2.6. Estimation of Phenolic Content
2.7. Statistical Analysis
3. Results
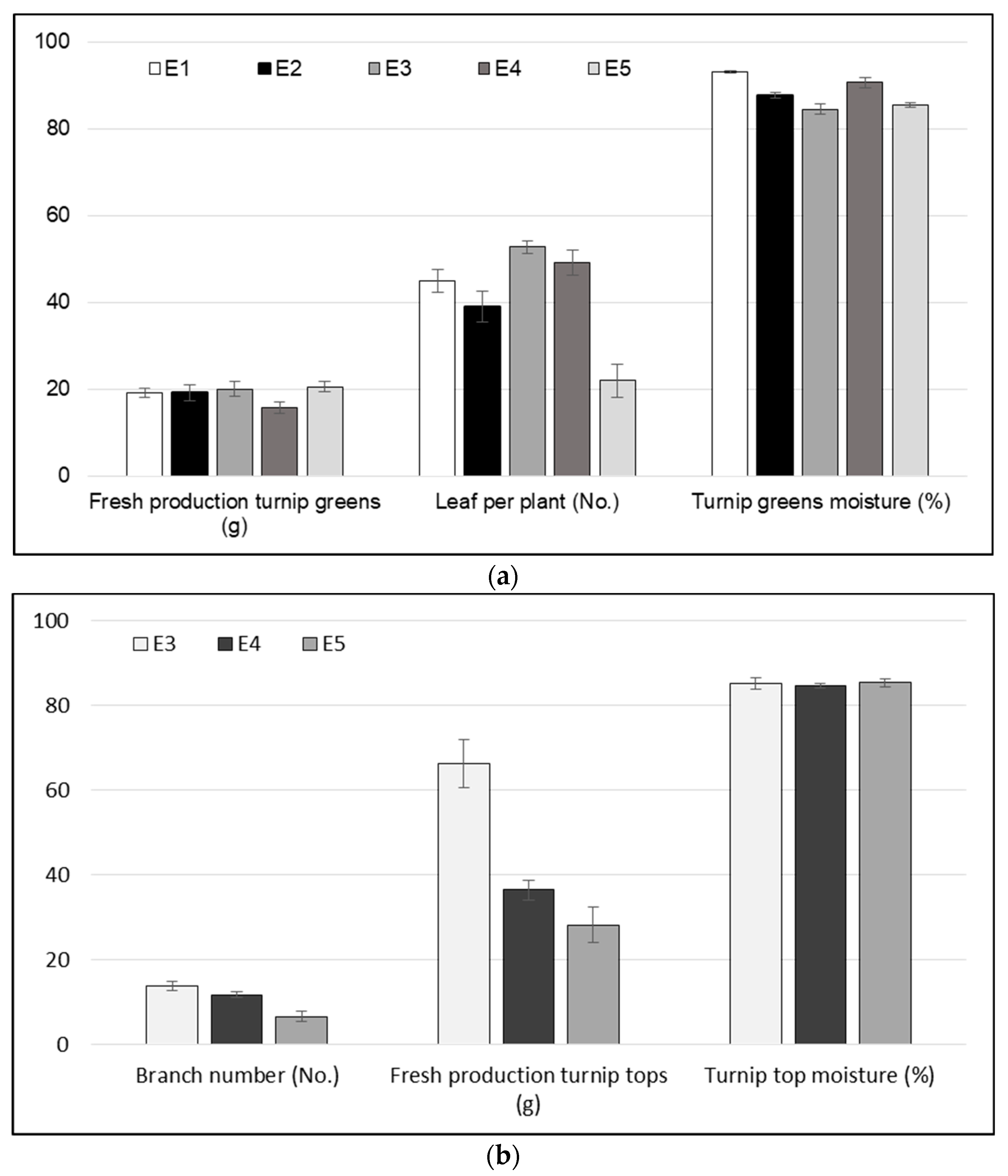
3.1. Agronomical Traits
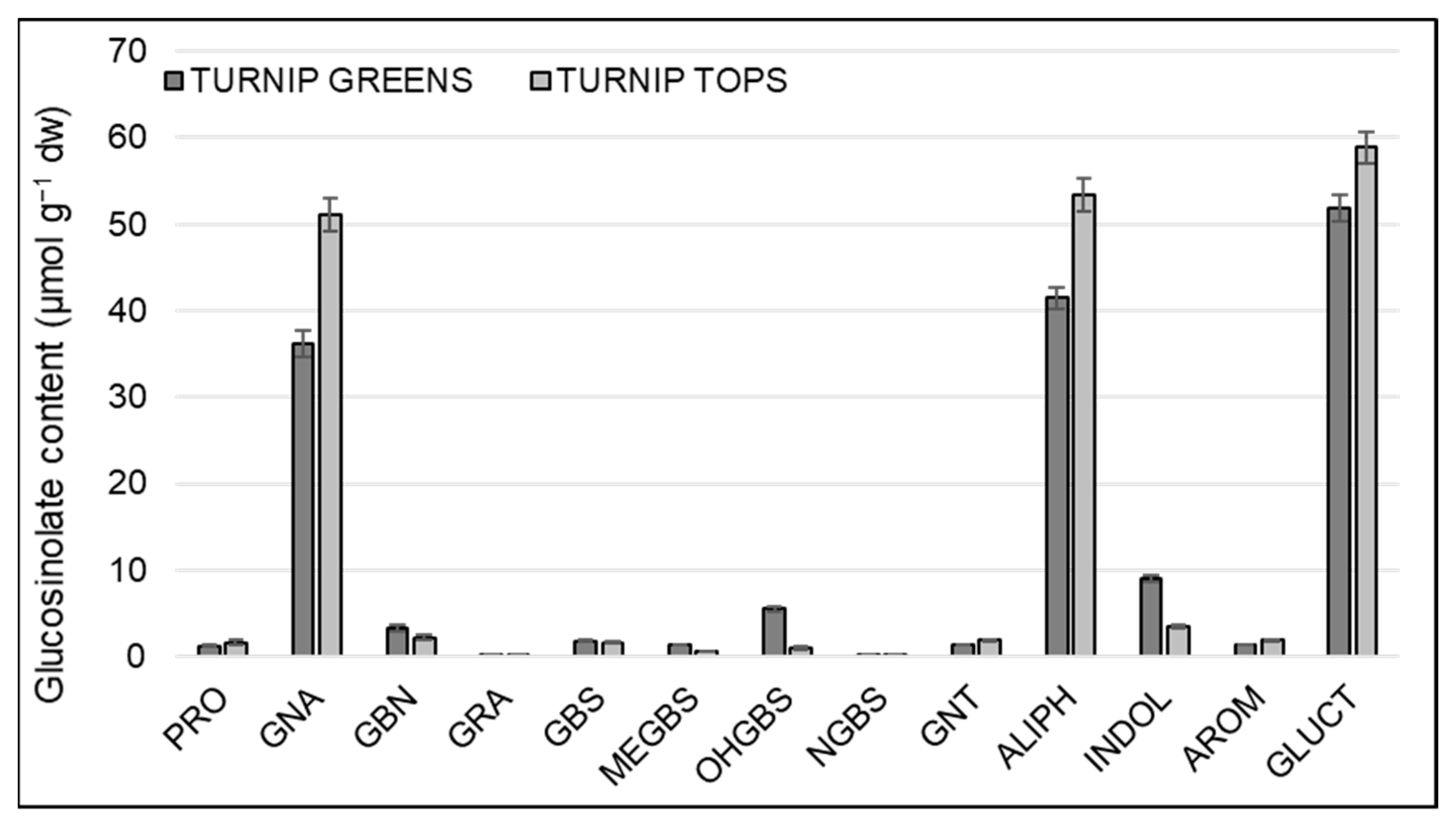
3.2. Variation of GSLs among Varieties, Locations, and Plant Organs
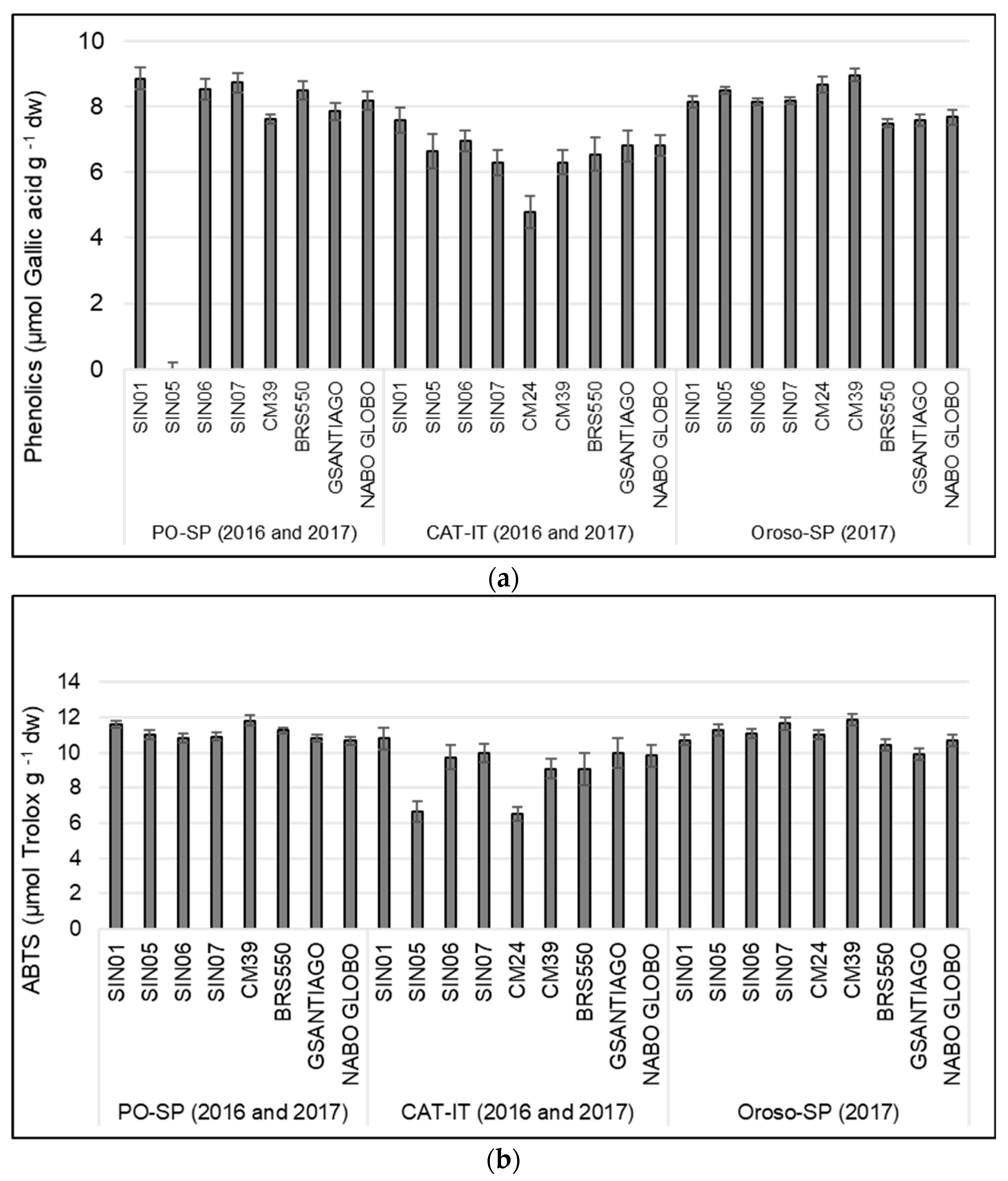
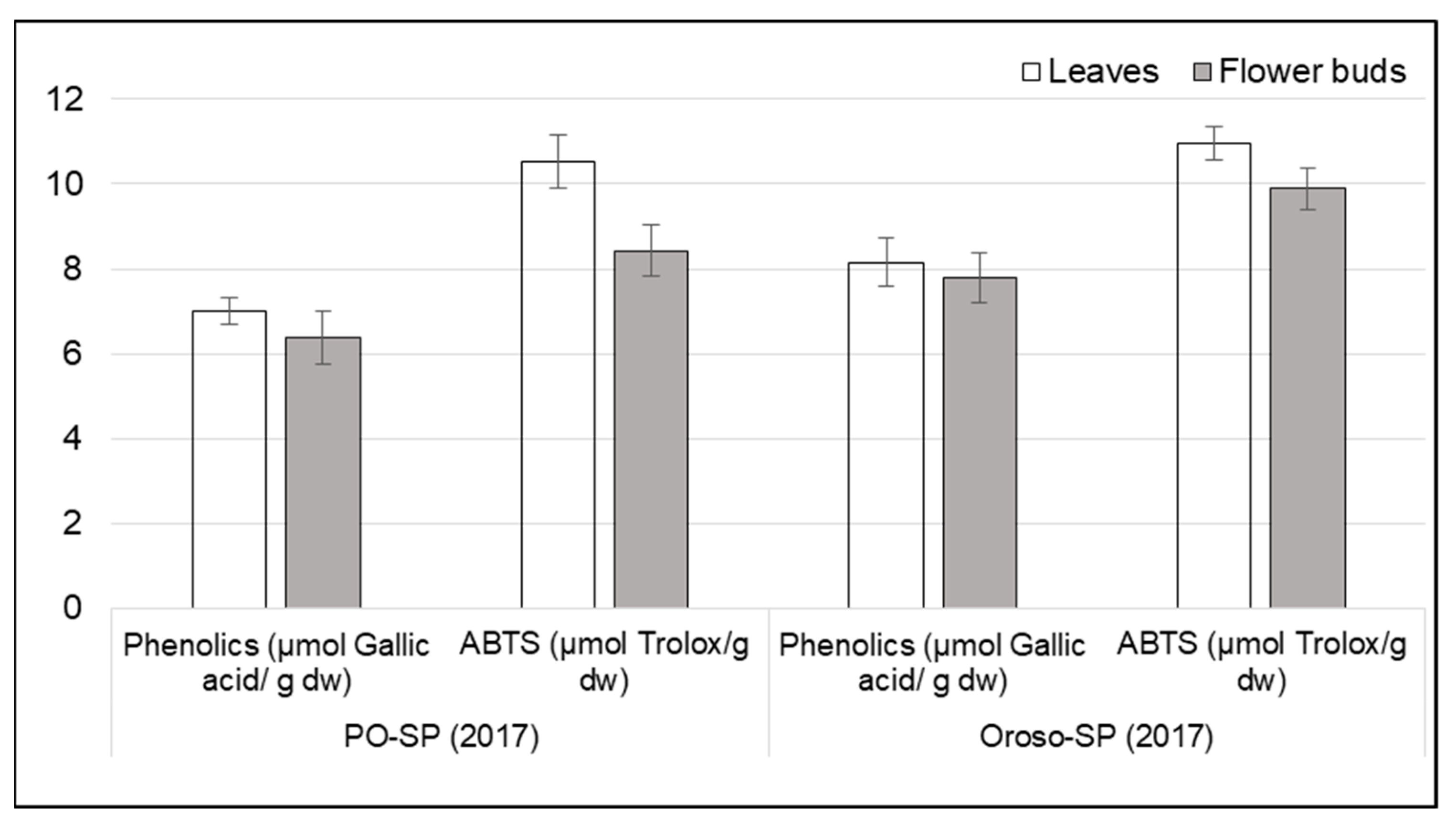
3.3. Variation of Phenolic Content and Antioxidant Activity among Varieties, Locations, and Plant Organs
4. Discussion
Bioactive Compounds
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dixon, G.R. Origins and diversity of Brassica and its relatives. In Vegetable Brassicas and Related Crucifers; Dixon, G.R., Ed.; CABI: Wallingford, UK, 2006; pp. 1–33. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Wang, X.; Deng, B.; Lou, P.; Wu, J.; Sun, R.; Xu, Z.; Vromans, J.; Koornneef, M. Genetic relationships within Brassica rapa as inferred from AFLP fingerprints. Theor. Appl. Genet. 2005, 110, 1301–1314. [Google Scholar] [CrossRef] [PubMed]
- Prakash, S.; Hinata, K. Taxonomy, cytogenetics and origin of crop Brassicas, a review. Opera. Bot. 1980, 55, 3–57. [Google Scholar]
- Monteiro, A.; Dias, J. Portuguese Brassica landraces: A perspective. Acta Hortic. 1996, 407, 87–94. [Google Scholar] [CrossRef]
- Bianco, V.V.; Damato, G.; Pomarici, R. Sowing and transplant dates in four cima di rapa (Brassica rapa L.) cultivars.—I. Sowing dates. Acta Hort. 1996, 407, 293–295. [Google Scholar] [CrossRef]
- Padilla, G.; Cartea, M.E.; Rodríguez, V.M.; Ordás, A. Genetic diversity in a germplasm collection of Brassica rapa subsp rapa L. from northwestern Spain. Euphytica 2005, 145, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Francisco, M.; Cartea, M.E.; Soengas, P.; Velasco, P. Effect of genotype and environmental conditions on health-promoting compounds in Brassica rapa. J. Agric. Food Chem. 2011, 59, 2421–2431. [Google Scholar] [CrossRef]
- Cartea, M.E.; De Haro, A.; Obregón, S.; Soengas, P.; Velasco, P. Glucosinolate variation in leaves of Brassica rapa crops. Plant Foods Hum. Nutr. 2012, 67, 283–288. [Google Scholar] [CrossRef]
- Soengas, P.; Cartea, M.E.; Francisco, M.; Velasco, P. Genetic structure and diversity of a collection of Brassica rapa subsp. rapa L. revealed by simple sequence repeat markers. J. Agric. Sci. 2011, 149, 617–624. [Google Scholar] [CrossRef]
- Cartea, M.E.; Velasco, P. Glucosinolates in Brassica foods: Bioavailability in food and significance for human health. Phytochem. Rev. 2008, 7, 213–229. [Google Scholar] [CrossRef]
- Hong, E.Y.; Kim, G.H. Anticancer and antimicrobial activities of β-phenylethyl isothiocyanate in Brassica rapa L. Food Sci. Technol. Res. 2008, 14, 377–382. [Google Scholar] [CrossRef] [Green Version]
- Traka, M.; Mithen, R. Glucosinolates, isothiocyanates and human health. Phytochem. Rev. 2009, 8, 269–282. [Google Scholar] [CrossRef]
- Paul, S.; Geng, C.A.; Yang, T.H.; Yang, Y.P.; Chen, J.J. Phytochemical and health-beneficial progress of turnip (Brassica rapa). J. Food Sci. 2019, 84, 19–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzo, M.N.; Pezzino, F.; Amodeo, L.; Catalano, D.; Viola, M.; Tribulato, A.; Travali, S.; Branca, F. Evaluation of a Sicilian black broccoli extract on in vitro cell models. Acta Hortic. 2018, 1202, 135–142. [Google Scholar] [CrossRef]
- Terzo, M.N.; Russo, A.; Ficili, B.; Pezzino, F.M.; Tribulato, A.; Russello, D.; Branca, F.; Travali, S. Neglected Sicilian landraces of black broccoli (Brassica oleracea var. italica Plenck) and health benefits: An in vivo study. Acta Hortic. 2020, 1267, 91–96. [Google Scholar] [CrossRef]
- Fernandes, F.; Valentao, P.; Sousa, C.; Pereira, J.A.; Seabra, R.M.; Andrade, P.B. Chemical and antioxidative assessment of dietary turnip (Brassica rapa var. rapa L.). Food Chem. 2007, 105, 1003–1010. [Google Scholar] [CrossRef]
- Shawon, R.A.; Kang, B.S.; Lee, S.G.; Kim, S.K.; Lee, H.J.; Katrich, E.; Gorinstein, S.; Ku, Y.G. Influence of drought stress on bioactive compounds, antioxidant enzymes and glucosinolate contents of Chinese cabbage (Brassica rapa). Food Chem. 2020, 308, 125657. [Google Scholar] [CrossRef] [PubMed]
- Cámara-Martos, F.; Obregón-Cano, S.; Mesa-Plata, O.; Cartea-González, M.E.; de Haro–Bailón, A. Quantification and in vitro bioaccessibility of glucosinolates and trace elements in Brassicaceae leafy vegetables. Food Chem. 2021, 339, 127860. [Google Scholar] [CrossRef]
- Singh, A. Glucosinolates and plant defense. In Glucosinolates; Mérillon, J.M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–26. [Google Scholar] [CrossRef]
- Núñez-Iglesias, M.J.; Novio, S.; García, C.; Soengas, P.; Cartea, M.E.; Velasco, P.; Freire-Garabal, M. Glucosinolate-degradation products as coadjuvant therapy on prostate cancer in vitro. Int. J. Mol. Sci. 2019, 20, 4977. [Google Scholar] [CrossRef] [Green Version]
- Llorach, R.; Espian, J.C.; S-Barberan, F.A.T.; Ferreres, F. Valorization of cauliflower (Brassica oleracea L. var. botrytis) by-products as a source of antioxidant phenolics. J. Agric. Food Chem. 2003, 51, 2181–2187. [Google Scholar] [CrossRef]
- Branca, F.; Chiarenza, G.L.; Cavallaro, C.; Gu, H.; Zhao, Z.; Tribulato, A. Diversity of Sicilian broccoli (Brassica oleracea var. italica) and cauliflower (Brassica oleracea var. botrytis) landraces and their distinctive bio-morphological, antioxidant, and genetic traits. Genet. Resour. Crop. Ev. 2018, 6, 485–502. [Google Scholar] [CrossRef]
- Di Bella, M.C.; Niklas, A.; Toscano, S.; Picchi, V.; Romano, D.; Lo Scalzo, R.; Branca, F. Morphometric characteristics, polyphenols and ascorbic acid variation in Brassica oleracea L. Novel Foods: Sprouts, Microgreens and Baby Leaves. Agronomy 2020, 10, 782. [Google Scholar] [CrossRef]
- IBPGR. Descriptors for Brassica and Raphanus; International Board for Plant Genetic Resources: Rome, Italy, 1990. [Google Scholar]
- Kliebenstein, D.J.; Lambrix, V.; Reichelt, M.; Gershenzon, J.; Mitchell-Olds, T. Gene duplication and the diversification of secondary metabolism: Sidechain modification of glucosinolates in Arabidopsis thaliana. Plant Cell 2001, 13, 681–693. [Google Scholar] [CrossRef] [Green Version]
- Samarth, R.; Panwar, M.; Kumar, M.; Soni, A. Evaluation of antioxidant and radical-scavenging activities of certain radioprotective plant extracts. Food Chem. 2008, 106, 868–873. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radical Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Adom, K.; Liu, R. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Steel, R.D.G.; Torrie, J.H.; Dickey, D.A. Principles and Procedures in Statistics: A Biometrical Approach; McGraw Hill: New York, NY, USA, 1997. [Google Scholar]
- Lou, P.; Zhao, J.; Kim, J.S.; Shen, S.; Del Carpio, D.P.; Song, X.; Jin, M.; Vreugdenhil, D.; Wang, X.; Koornneef, M.; et al. Quantitative trait loci for flowering time and morphological traits in multiple populations of Brassica rapa. J. Experim. Bot. 2007, 58, 4005–4016. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Quiros, C. Survey of glucosinolate variation in leaves of Brassica rapa crops. Gen. Res. Crop Evol. 2010, 57, 1079–1089. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Kawaguchi, S.; Watanabe, Y. Glucosinolates in vegetative tissues and seeds of twelve cultivars of vegetable turnip rape (Brassica rapa L.). Soil Sci. Plant Nutr. 2003, 49, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Charron, C.S.; Saxton, A.M.; Sams, C.E. Relationship of climate ad genotype to seasonal variation in the glucosinolate-mirosinase system I. Glucosinolate content in ten cultivars of Brassica oleracea grown in fall and spring seasons. J. Sci. Food Agric. 2005, 85, 671–681. [Google Scholar] [CrossRef]
- Ciska, E.; Martyniak-Przybyszewska, B.; Kozlowska, H. Content of glucosinolates in cruciferous vegetables grown at the same site for two years under different climatic conditions. J. Agric. Food Chem. 2000, 48, 2862–2867. [Google Scholar] [CrossRef]
- Velasco, P.; Cartea, M.E.; González, C.; Vilar, M.; Ordás, A. Factors affecting the glucosinolate content of kale (Brassica oleracea acephala group). J. Agric. Food Chem. 2007, 55, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Bonnema, G.; Lee, J.G.; Shuhang, W.; Lagarrigue, D.; Bucher, J.; Wehrens, R.; de Vos, R.; Beekwilder, J. Glucosinolate variability between turnip organs during development. PLoS ONE 2019, 14, e0217862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, E.A.S.; Heaney, R.K.; Fenwick, G.R.; Portas, C.A.M. Glucosinolates in crop plants. Hortic. Rev. 1997, 19, 99–215. [Google Scholar]
- Francisco, M.; Velasco, P.; Romero, A.; Vazquez, L.; Cartea, M.E. Sensory quality of turnip greens and turnip tops grown in northwestern Spain. Eur. Food Res. Technol. 2009, 230, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, S.R.; Jo, J.S.; Lee, J.G. Comparison of glucosinolate profiles in different tissues of nine Brassica crops. Molecules 2015, 20, 15827–15841. [Google Scholar] [CrossRef]
- Kim, J.K.; Chu, S.M.; Kim, S.J.; Lee, D.J.; Lee, S.Y.; Lim, S.H.; Ha, S.H.; Kweon, S.J.; Cho, H.S. Variation of glucosinolates in vegetable crops of Brassica rapa L. ssp. pekinensis. Food Chem. 2010, 119, 423–428. [Google Scholar] [CrossRef]
- Samec, D.; Piljac-Zagarac, J.; Bogovic, M.; Habjanic, K.; Gruz, J. Antioxidant potency of white (Brassica oleracea L. var. capitata) and Chinese (Brassica rapa L. var. pekinensis (Lour.)) cabbage: The influence of development stage, cultivar choice and seed selection. Sci. Hortic. 2011, 128, 78–83. [Google Scholar] [CrossRef]
- Soengas, P.; Cartea, M.E.; Francisco, M.; Sotelo, T.; Velasco, P. New insights into antioxidant activity of Brassica crops. Food Chem. 2012, 134, 725–733. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety Name | Description | Origin | Source 1 | Flowering Earliness |
|---|---|---|---|---|
| SIN05 C3 | Synthetic | Spain | MBG | Early |
| SIN06 C3 | Synthetic | Spain | MBG | Medium |
| SIN07 C3 | Synthetic | Spain | MBG | Late |
| SIN01 C4 | Synthetic | Spain | MBG | Medium |
| BRS0550 | Landrace | Spain | MBG | Extra-late |
| Nabo globo de Lugo | Commercial | Spain | Rocalba | Medium |
| Grelos de Santiago | Commercial | Spain | Rocalba | Late |
| Broccoletto di Rapa sessantino Riccio San Marzano (“CM24”) | Landrace | Italy | UNICT 817 | Extra-early |
| Nabo sessantino (“CM39”) | Commercial | Italy | UNICT 3272 | Extra-early |
| Agronomic Traits | Description |
|---|---|
| Time to flowering (d) | Days from transplanting until 50% of plants have the first flower |
| Turnip greens fresh production (g) | Average fresh weight of 30 leaves per plot |
| Turnip greens moisture (%) | Leaf water content |
| Branch number (n) | Average number of secondary stems per plant at first flower opening of five plants per plot |
| Leaf per plant (n) | Average number of leaves per plant at first flower opening of five plants per plot |
| Turnip tops fresh production (g) | Average fresh weight of the turnip top of five plants per plot |
| Turnip tops moisture (%) | Turnip top water content |
| Country | Varieties | Time to Flowering (d) | Turnip Greens Fresh Production(g) | Turnip Greens Moisture (%) | Branch Number1 (n) | Leaf per Plant (n) | Turnip Tops Fresh Production 1 (g) | Turnip Top Moisture 1 (%) |
|---|---|---|---|---|---|---|---|---|
| Spain | BRS550 | 205.67 ± 2.36 | 27.73 ± 1.25 | 89.95 ± 0.62 | 15.72 ± 1.26 | 65.21 ± 10.2 | 65.27 ± 6.95 | 91.43 ± 2.56 |
| CM24 | 70.33 ± 1.33 | 8.30 ± 1.57 | 91.27 ± 0.30 | 7.21 ± 0.84 | 18.10 ± 2.9 | 24.66 ± 1.75 | 91.71 ± 1.65 | |
| CM39 | 84.32 ± 3.11 | 7.23 ± 0.87 | 82.43 ± 3.41 | 6.53 ± 1.54 | 24.13 ± 7.5 | 20.52 ± 2.10 | 85.81 ± 0.78 | |
| SIN01C4 | 145.11 ± 4.16 | 21.15 ± 1.68 | 90.34 ± 0.90 | 12.77 ± 0.79 | 63.84 ± 10.2 | 54.16 ± 6.65 | 85.05 ± 1.05 | |
| SIN05 C3 | 117.00 ± 3.34 | 11.21 ± 1.29 | 89.65 ± 3.19 | 8.54 ± 1.84 | 42.51 ± 5.3 | 29.69 ± 4.73 | 79.38 ± 0.88 | |
| SIN06 C3 | 140.00 ± 4.69 | 17.92 ± 1.63 | 90.08 ± 1.18 | 11.39 ± 1.41 | 55.14 ± 8.9 | 47.98 ± 5.69 | 83.85 ± 1.22 | |
| SIN07 C3 | 152.11 ± 2.19 | 21.02 ± 1.93 | 90.55 ± 0.78 | 14.74 ± 0.59 | 72.23 ± 11.5 | 65.67 ± 12.71 | 84.06 ± 2.02 | |
| Grelos de Santiago | 161.33 ± 3.26 | 18.28 ± 1.38 | 89.84 ± 1.11 | 13.26 ± 0.89 | 63.61 ± 6.3 | 65.36 ± 13.45 | 85.92 ± 0.92 | |
| Nabo Globo Lugo | 156.44 ± 2.79 | 18.07 ± 1.36 | 89.77 ± 0.99 | 13.32 ± 1.21 | 69.32 ± 8.9 | 61.42 ± 8.94 | 84.72 ± 1.25 | |
| LSD (5%) | 3.84 | 3.52 | 1.08 | 2.95 | 26.31 | 11.73 | 3.68 | |
| Italy | BRS550 | -2 | 16.50 ± 4.23 | 87.41 ± 1.05 | 6.33 ± 0.88 | 26.78 ± 5.9 | 74.00 ± 26.00 | 87.25 ± 2.19 |
| CM24 | 58.50 ± 6.50 | 9.81 ± 2.06 | 84.94 ± 3.00 | 6.13 ± 0.81 | 23.10 ± 5.6 | 20.00 ± 0.99 | 82.14 ± 1.12 | |
| CM39 | 64.60 ± 0.40 | 26.52 ± 4.02 | 85.61 ± 0.53 | 7.83 ± 0.46 | 16.78 ± 2.8 | 60.40 ± 7.60 | 83.15 ± 0.43 | |
| SIN01C4 | 82.67 ± 5.20 | 23.03 ± 2.42 | 87.09 ± 0.89 | 4.89 ± 0.61 | 46.89 ± 18.9 | 20.42 ± 4.08 | 86.29 ± 0.99 | |
| SIN05 C3 | 78.83 ± 4.03 | 15.22 ± 2.32 | 87.96 ± 1.11 | 6.78 ± 0.83 | 26.89 ± 7.5 | 18.43 ± 2.00 | 83.16 ± 0.91 | |
| SIN06 C3 | 84.25 ± 6.36 | 14.80 ± 2.33 | 88.43 ± 1.50 | 4.56 ± 0.38 | 45.8 ± 10.2 | 17.67 ± 3.84 | 86.52 ± 1.20 | |
| SIN07 C3 | 92.50 ± 3.48 | 28.38 ± 4.10 | 86.62 ± 0.53 | 7.22 ± 0.89 | 41.71 ± 8.9 | 20.88 ± 8.08 | 83.62 ± 0.73 | |
| Grelos de Santiago | 111.17 ± 0.65 | 15.81 ± 1.95 | 85.55 ± 0.82 | 6.44 ± 0.71 | 53.22 ± 11.9 | 22.72 ± 5.90 | 81.55 ± 0.92 | |
| Nabo Globo Lugo | 130.00 ± 1.15 | 20.90 ± 3.66 | 86.61 ± 0.95 | 8.44 ± 1.71 | 55.50 ± 25.6 | 31.60 ± 10.28 | 84.61 ± 0.85 | |
| LSD (5%) | 4.52 | 8.35 | 3.62 | 4.18 | 23.59 | 30.61 | 4.25 |
| Variety | PRO | GRA | GNA | GBN | GBS | OHGBS | MGBS | NGBS | ALIPH | INDOL | AROM | Total GLSs |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SIN01C4 | 0.80 ± 0.13 | 0.03 ± 0.01 | 40.14 ± 2.63 | 1.95 ± 0.22 | 1.68 ± 0.18 | 5.60 ± 0.63 | 1.34 ± 0.13 | 0.41 ± 0.06 | 42.97 ± 2.69 | 9.02 ± 0.84 | 0.78 ± 0.15 | 52.77 ± 3.29 |
| SIN05C3 | 1.74 ± 0.22 | 0.07 ± 0.04 | 21.63 ± 2.43 | 1.23 ± 0.21 | 1.17 ± 0.14 | 3.23 ± 0.51 | 0.83 ± 0.11 | 0.20 ± 0.04 | 24.67 ± 2.64 | 5.42 ± 0.73 | 0.35 ± 0.10 | 30.45 ± 3.32 |
| SIN06C3 | 1.20 ± 0.16 | 0.15 ± 0.06 | 30.91 ± 2.81 | 2.10 ± 0.36 | 1.43 ± 0.16 | 6.32 ± 0.94 | 1.16 ± 0.11 | 0.34 ± 0.05 | 34.50 ± 2.87 | 9.25 ± 1.15 | 0.61 ± 0.11 | 44.36 ± 3.56 |
| SIN07C3 | 1.49 ± 0.20 | 0.02 ± 0.02 | 36.14 ± 2.08 | 3.61 ± 0.45 | 1.60 ± 0.14 | 6.81 ± 0.85 | 1.33 ± 0.13 | 0.33 ± 0.03 | 41.44 ± 2.16 | 10.07 ± 1.04 | 0.98 ± 0.14 | 52.50 ± 2.83 |
| BRS550 | 2.59 ± 0.19 | 0.23 ± 0.57 | 32.49 ± 2.50 | 2.57 ± 0.30 | 1.60 ± 0.10 | 9.18 ± 0.90 | 1.92 ± 0.12 | 0.19 ± 0.02 | 37.98 ± 2.63 | 12.85 ± 1.05 | 0.69 ± 0.14 | 51.52 ± 3.24 |
| CM24 | 0.09 ± 0.05 | 0 | 5.35 ± 0.87 | 6.33 ± 1.24 | 0.79 ± 0.19 | 1.01 ± 0.17 | 0.29 ± 0.02 | 0.21 ± 0.04 | 12.09 ± 2.17 | 2.30 ± 0.37 | 1.36 ± 0.29 | 15.74 ± 2.76 |
| CM39 | 1.58 ± 0.32 | 0 | 6.58 ± 0.51 | 8.73 ± 0.75 | 1.16 ± 0.18 | 2.00 ± 0.49 | 0.71 ± 0.09 | 0.39 ± 0.06 | 18.71 ± 1.38 | 4.25 ± 0.66 | 0.97 ± 0.15 | 23.94 ± 1.88 |
| Grelos de Santiago | 0.21 ± 0.10 | 0.32 ± 0.08 | 33.62 ± 2.22 | 1.56 ± 0.24 | 1.39 ± 0.12 | 5.37 ± 0.55 | 1.16 ± 0.13 | 0.40 ± 0.06 | 35.85 ± 2.31 | 8.32 ± 0.74 | 1.35 ± 0.25 | 45.52 ± 2.93 |
| Nabo G. Lugo | 0.44 ± 0.19 | 0.04 ± 0.02 | 33.97 ± 2.70 | 1.21 ± 0.17 | 1.51 ± 0.16 | 4.48 ± 0.35 | 1.20 ± 0.13 | 0.32 ± 0.04 | 35.84 ± 2.72 | 7.51 ± 0.57 | 0.85 ± 0.14 | 44.20 ± 3.24 |
| LSD (5%) | 0.46 | 0.12 | 3.89 | 1.01 | 0.30 | 1.45 | 0.20 | 0.12 | 4.04 | 1.74 | 0.29 | 5.13 |
| Variety | PRO | GRA | GNA | GBN | GBS | OHGBS | MGBS | NGBS | ALIPH | INDOL | AROM | Total GLSs |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SIN01C4 | 1.00 ± 0.55 | 0.01 ± 0.01 | 68.93 ± 4.27 | 2.04 ± 0.41 | 2.00 ± 0.16 | 0.12 ± 0.07 | 0.99 ± 0.08 | 0.34 ± 0.10 | 71.98 ± 3.98 | 3.46 ± 0.27 | 2.01 ± 0.14 | 77.44 ± 4.09 |
| SIN05C3 | 3.98 ± 0.42 | 0.45 ± 0.11 | 36.13 ± 3.32 | 0.85 ± 0.12 | 2.02 ± 0.24 | 3.30 ± 0.83 | 0.49 ± 0.12 | 0.13 ± 0.03 | 41.43 ± 3.55 | 5.95 ± 1.15 | 2.20 ± 0.19 | 49.57 ± 4.51 |
| SIN06C3 | 2.10 ± 0.74 | 0.25 ± 1.71 | 46.91 ± 5.37 | 1.96 ± 0.46 | 2.16 ± 0.31 | 0.20 ± 0.07 | 0.66 ± 0.08 | 0.20 ± 0.03 | 51.23 ± 5.51 | 3.20 ± 0.35 | 1.51 ± 0.19 | 55.94 ± 5.73 |
| SIN07C3 | 1.77 ± 0.46 | 0.04 ± 0.02 | 59.91 ± 4.45 | 2.84 ± 0.63 | 1.89 ± 0.20 | 0.68 ± 0.15 | 0.59 ± 0.11 | 0.20 ± 0.05 | 64.62 ± 4.24 | 3.36 ± 0.30 | 2.15 ± 0.28 | 70.14 ± 4.34 |
| CM24 | 0 | 0 | 8.93 ± 0.93 | 9.49 ± 0.75 | 0.94 ± 0.14 | 2.80 ± 0.30 | 0.09 ± 0.05 | 0.29 ± 0.07 | 19.51 ± 1.71 | 4.13 ± 0.48 | 3.13 ± 0.34 | 26.77 ± 2.35 |
| BRS0550 | 5.64 ± 0.73 | 0 | 62.51 ± 3.34 | 1.71 ± 0.18 | 0.79 ± 0.04 | 1.48 ± 0.05 | 0.42 ± 0.05 | 0.19 ± 0.03 | 69.94 ± 3.29 | 2.88 ± 0.11 | 0.94 ± 0.03 | 73.76 ± 3.41 |
| Grelos de Santiago | 0.04 ± 0.02 | 0.44 ± 0.10 | 53.00 ± 2.35 | 1.33 ± 0.28 | 1.34 ± 0.08 | 0.30 ± 0.08 | 0.62 ± 0.10 | 0.18 ± 0.03 | 54.83 ± 2.38 | 2.44 ± 0.16 | 2.04 ± 0.25 | 59.29 ± 2.36 |
| Nabo Globo Lugo | 0 | 0.10 ± 0.04 | 57.47 ± 3.54 | 0.96 ± 0.25 | 1.30 ± 0.13 | 0.57 ± 0.10 | 0.50 ± 0.11 | 0.24 ± 0.07 | 58.68 ± 3.49 | 2.61 ± 0.20 | 1.42 ± 0.23 | 62.71 ± 3.49 |
| LSD (5%) | 1.32 | 0.26 | 11.06 | 1.21 | 0.56 | 0.43 | 0.26 | 0.18 | 10.7 | 0.87 | 0.57 | 11.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cartea, M.E.; Di Bella, M.C.; Velasco, P.; Soengas, P.; Toscano, S.; Branca, F. Evaluation of Italian and Spanish Accessions of Brassica rapa L.: Effect of Flowering Earliness on Fresh Yield and Biological Value. Agronomy 2021, 11, 29. https://doi.org/10.3390/agronomy11010029
Cartea ME, Di Bella MC, Velasco P, Soengas P, Toscano S, Branca F. Evaluation of Italian and Spanish Accessions of Brassica rapa L.: Effect of Flowering Earliness on Fresh Yield and Biological Value. Agronomy. 2021; 11(1):29. https://doi.org/10.3390/agronomy11010029
Chicago/Turabian StyleCartea, María Elena, Maria Concetta Di Bella, Pablo Velasco, Pilar Soengas, Stefania Toscano, and Ferdinando Branca. 2021. "Evaluation of Italian and Spanish Accessions of Brassica rapa L.: Effect of Flowering Earliness on Fresh Yield and Biological Value" Agronomy 11, no. 1: 29. https://doi.org/10.3390/agronomy11010029
APA StyleCartea, M. E., Di Bella, M. C., Velasco, P., Soengas, P., Toscano, S., & Branca, F. (2021). Evaluation of Italian and Spanish Accessions of Brassica rapa L.: Effect of Flowering Earliness on Fresh Yield and Biological Value. Agronomy, 11(1), 29. https://doi.org/10.3390/agronomy11010029