Improvement of Tillering and Grain Yield by Application of Cytokinin Derivatives in Wheat and Barley

 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
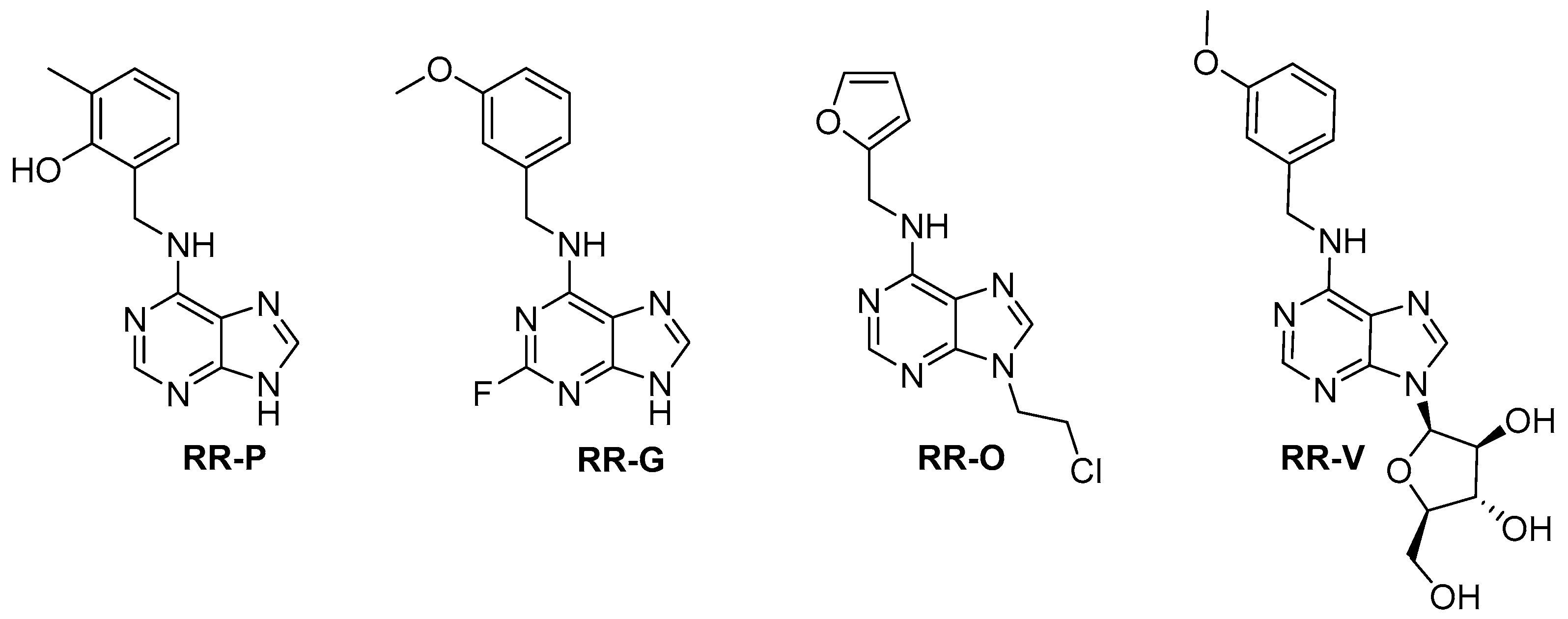
2.1. Compounds Structures
2.2. Analysis of Endogenous Cytokinins
2.3. CK Activity Bioassays
2.4. Field Plot Experiments
2.5. Statistical Analysis
3. Results
3.1. CK Activity Bioassays
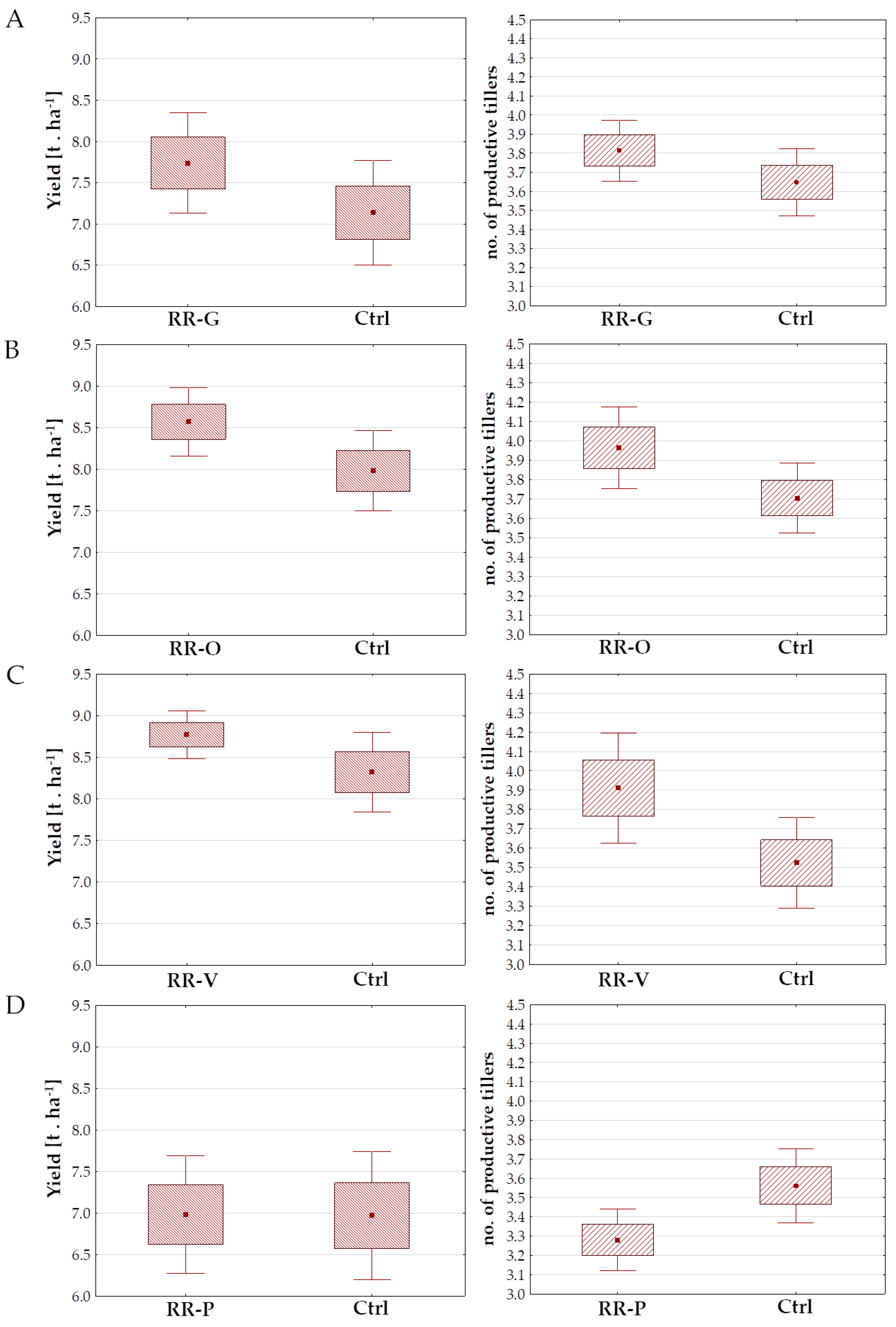
3.2. Effect of CK Derivatives and CK Antagonist on Tillering and Grain Yield
3.3. Analysis of Endogenous Cytokinin Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture organization of the United Nations. 2008. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 17 October 2020).
- Darwinkel, A. Ear development and formation of grain yield in winter wheat. Neth. J. Agric. Sci. 1980, 28, 156–163. [Google Scholar] [CrossRef]
- Kirby, E.J.M.; Jones, H.G. The relationhips between the main shoot and tillers in barley plants. J. Agric. Sci. 1977, 78, 281–288. [Google Scholar] [CrossRef]
- Yang, M.-D.; Leghari, S.J.; Guan, X.-K.; Ma, S.-C.; Ding, C.-M.; Mei, F.-J.; Wei, L.; Wang, T.-C. Deficit Subsurface Drip Irrigation Improves Water Use Efficiency and Stabilizes Yield by Enhancing Subsoil Water Extraction in Winter Wheat. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Kirby, E.J.M.; Riggs, T.J. Developmental consequences of two-row and six-row ear type in spring barley: 2. Shoot apex, leaf and tiller development. J. Agric. Sci. 1978, 91, 207–216. [Google Scholar] [CrossRef]
- Kulshrestha, V.P.; Chowdhury, S. A new selection criterion for yield in wheat. Theor. Appl. Genet. 1987, 74, 275–279. [Google Scholar] [CrossRef]
- Simmons, S.R.; Rasmusson, D.C.; Wiersma, J.V. Tillering in Barley: Genotype, Row Spacing, and Seeding Rate Effects 1. Crop. Sci. 1982, 22, 801–805. [Google Scholar] [CrossRef]
- Thorne, G.N. Survival of Tillers and Distribution of Dry Matter between Ear and Shoot of Barley Varieties. Ann. Bot. 1962, 26, 37–54. [Google Scholar] [CrossRef]
- Del Moral, M.; Del Moral, L. Tiller production and survival in relation to grain yield in winter and spring barley. Field Crop. Res. 1995, 44, 85–93. [Google Scholar] [CrossRef]
- McSteen, P. Hormonal Regulation of Branching in Grasses. Plant Physiol. 2009, 149, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.O.; Skoog, F.; Von Saltza, M.H.; Strong, F.M.; Kinetin, A. Cell Division Factor from Deoxyribonucleic Acid1. J. Am. Chem. Soc. 2005, 77, 1392. [Google Scholar] [CrossRef]
- Letham, D.S. Zeatin, a Factor Inducing Cell Division from Zea mays. Life Sci. 1963, 2, 569–573. [Google Scholar] [CrossRef]
- Letham, D. Regulators of cell division in plant tissues. N. Z. J. Bot. 2012, 1, 336–350. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, H.; Kasahara, H.; Ueda, N.; Kojima, M.; Takei, K.; Hishiyama, S.; Asami, T.; Okada, K.; Kamiya, Y.; Yamaya, T.; et al. Agrobacterium tumefaciens increases cytokinin production in plastids by modifying the biosynthetic pathway in the host plant. Proc. Natl. Acad. Sci. USA 2005, 102, 9972–9977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takei, K.; Sakakibara, H.; Sugiyama, T. Identification of Genes Encoding Adenylate Isopentenyltransferase, a Cytokinin Biosynthesis Enzyme, inArabidopsis thaliana. J. Biol. Chem. 2001, 276, 26405–26410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakimoto, T. Identification of plant cytokinin biosynthetic enzymes as dimethylallyl diphosphate: ATP/ADP isopentenyl-transferases. Plant Cell Physiol. 2001, 42, 677–685. [Google Scholar] [CrossRef]
- Schmülling, T.; Werner, T.; Riefler, M.; Krupková, E.; Manns, I.B. Structure and function of cytokinin oxi-dase/dehydrogenase genes of maize, rice, Arabidopsis and other species. J. Plant Res. 2003, 116, 241–252. [Google Scholar] [CrossRef]
- Mok, M.C. Cytokinins and plant development-An overview. In Cytokinins: Chemistry, Activity and Function; Mok, D.W.S., Mok, M.C., Eds.; CRC: Boca Raton, FL, USA, 1994; pp. 155–166. [Google Scholar]
- Sharif, R.; Dale, J.E. Growth-Regulating Substances and the Growth of Tiller Buds in Barley; Effects of IAA and GA3. J. Exp. Bot. 1980, 31, 1191–1197. [Google Scholar] [CrossRef]
- Ha, S.; Vankova, R.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.-S.P. Cytokinins: Metabolism and function in plant adaptation to environmental stresses. Trends Plant Sci. 2012, 17, 172–179. [Google Scholar] [CrossRef]
- Harrison, M.A.; Kaufman, P.B. Hormonal regulation of lateral bud (tiller) release in oats (Avena saliva L.). Plant Physiol. 1980, 66, 1123–1127. [Google Scholar] [CrossRef] [Green Version]
- Yeh, S.-Y.; Chen, H.-W.; Ng, C.-Y.; Lin, C.-Y.; Tseng, T.-H.; Li, W.-H.; Ku, M.S.B. Down-Regulation of Cytokinin Oxidase 2 Expression Increases Tiller Number and Improves Rice Yield. Rice 2015, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Holubová, K.; Hensel, G.; Vojta, P.; Tarkowski, P.; Bergougnoux, V.; Galuszka, P. Modification of Barley Plant Productivity Through Regulation of Cytokinin Content by Reverse-Genetics Approaches. Front. Plant Sci. 2018, 9, 1676. [Google Scholar] [CrossRef] [Green Version]
- Morris, R.D.; Blevins, D.G.; Dietrich, J.T.; Durley, R.C.; Gelvin, S.B.; Gray, J.; Hommes, N.G.; Kaminek, M.; Mathews, L.J.; Meilan, R. Cytokinins in plant pathogenic bacteria and developing cere-al grains. Aust. J. Plant Physiol. 1993, 20, 621–637. [Google Scholar]
- Harrison, J.C. Development and genetics in the evolution of land plant body plans. Philos. Trans. R. Soc. B 2017, 372, 20150490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugière, N.; Jiao, S.; Hantke, S.; Zinselmeier, C.; Roessler, J.A.; Niu, X.; Jones, R.J.; Habben, J.E. Cytokinin oxidase gene expression in maize is localized to the vasculature, and is induced by cytokinins, abscisic acid, and abiotic stress. Plant Physiol. 2003, 132, 1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, J.T.; Kaminek, M.; Blevins, D.G.; Reinbott, T.M.; Morris, R.O. Changes in cytokinins and cytokinin oxidase activ-ity in developing maize kernels and the effects of endogenous cytokinins on kernel development. Plant Physiol. Biochem. 1995, 33, 327–336. [Google Scholar]
- Williams, R.H.; Cartwright, P.M. The Effect of Applications of a Synthetic Cytokinin on Shoot Dominance and Grain Yield in Spring Barley. Ann. Bot. 1980, 46, 445–452. [Google Scholar] [CrossRef]
- Jameson, P.E.; Song, J. Cytokinin: A key driver of seed yield. J. Exp. Bot. 2016, 67, 593–606. [Google Scholar] [CrossRef] [Green Version]
- Koprna, R.; De Diego, N.; Dundálková, L.; Spíchal, L. Use of cytokinins as agrochemicals. Bioorg. Med. Chem. 2016, 24, 484–492. [Google Scholar] [CrossRef]
- Mik, V.; Szüčová, L.; Šmehilová, M.; Zatloukal, M.; Doležal, K.; Nisler, J.; Gruz, J.; Galuszka, P.; Strnad, M.; Spíchal, L. N9-substituted derivatives of kinetin: Effective anti-senescence agents. Phytochemistry 2011, 72, 821–831. [Google Scholar] [CrossRef]
- Podlešáková, K.; Zalabák, D.; Čudejková, M.; Plíhal, O.; Szüčová, L.; Doležal, K.; Spíchal, L.; Strnad, M.; Galuszka, P. Novel Cytokinin Derivatives Do Not Show Negative Effects on Root Growth and Proliferation in Submicromolar Range. PLoS ONE 2012, 7, e39293. [Google Scholar] [CrossRef]
- Bryksová, M.; Dabravolski, S.; Kučerová, Z.; Zavadil Kokáš, F.; Špundová, M.; Plíhalová, L.; Takáč, T.; Grúz, J.; Hudeček, M.; Hloušková, V.; et al. Aromatic Cytokinin Arabinosides Promote PAMP-like Responses and Positively Regulate Leaf Longevity. ACS Chem. Biol. 2020, 15, 1949–1963. [Google Scholar] [CrossRef] [PubMed]
- Nisler, J.; Zatloukal, M.; Popa, I.; Doležal, K.; Strnad, M.; Spíchal, L. Cytokinin receptor antagonists derived from 6-benzylaminopurine. Phytochemistry 2010, 71, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Spíchal, L.; Werner, T.; Popa, I.; Riefler, M.; Schmülling, T.; Strnad, M. The purine derivative PI-55 blocks cytokinin action via receptor inhibition. FEBS J. 2009, 276, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Spíchal, L.; Kryštof, V.; Paprskářová, M.; Lenobel, R.; Stýskala, J.; Binarová, P.; Cenklová, V.; De Veylder, L.; Inze, D.; Kontopidis, G.; et al. Classical Anticytokinins Do Not Interact with Cytokinin Recep-tors but Inhibit Cyclin-dependent Kinases. Biol. Chem. 2007, 282, 14356. [Google Scholar] [CrossRef] [Green Version]
- Popa, I.; Holub, J.; Lenobel, R.; Werbrouck, S.; Doležal, K.; Strnad, M.; Zatloukal, M.; Massino, F.J. Heterocyclic Compounds Based on N6-Substituted Adenine, Methods of Their Preparation, Their Use for Preparation of Drugs, Cosmetic Preparation and Growth Regulators, Pharmaceutical Preparations, Cosmetic Preparations and Growth Regulators Containing These Compounds. U.S. Patent 20080014227A1, 9 October 2008. [Google Scholar]
- Dolezal, K.; Popa, I.; Holub, J.; Lenobel, R.; Werbrouck, S.; Strnad, M.; Zatloukal, M. Heterocyclic Compounds Based on N6-Substituted Adenine, Methods of Their Preparation, Their Use for Preparation of Drugs, Cosmetic Preparation and Growth Regulators, Pharmaceutical Preparations, Cosmetic Preparations and Growth Regulators Containing These Compounds. U.S. Patent 2005.0043328A1, 24 February 2005. [Google Scholar]
- Svačinová, J.; Novák, O.; Plačková, L.; Lenobel, R.; Holík, J.; Strnad, M.; Doležal, K. A new approach for cytokinin isola-tion from Arabidopsis tissues using miniaturized purification: Pipette tip solid-phase extraction. Plant Methods 2012, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Antoniadi, I.; Plačková, L.; Simonovik, B.; Doležal, K.; Turnbull, C.; Ljung, K.; Novák, O. Cell-type specific cytokinin dis-tribution within the Arabidopsis primary root apex. Plant Cell 2015, 27, 1955–1967. [Google Scholar] [CrossRef] [Green Version]
- Rittenberg, D.; Foster, L. A new procedure for quantitative analysis by isotope dilution, with application to the determina-tion of amino acids and fatty acids. J. Biol. Chem. 1940, 133, 727–744. [Google Scholar]
- Holub, J.; Hanuš, J.; Hanke, D.E.; Strnad, M. Biological activity of cytokinins derived from Ortho- and Meta-Hydroxybenzyladenine. Plant Growth Regul. 1998, 26, 109–115. [Google Scholar] [CrossRef]
- Mathews, R.A.J. Beyond significance: Principles and Practise of the Analysis of Credibility. R. Soc. Open Sci. 2018, 5, 171047. [Google Scholar] [CrossRef] [Green Version]
- Goodman, S.N. How sure are you of your result? Put a number on it. Nat. Cell Biol. 2018, 564, 7. [Google Scholar] [CrossRef]
- Nisler, J.; Kopečný, D.; Pěkná, Z.; Končitíková, R.; Koprna, R.; Murvanidze, N.; Werbrouck, S.; Havlíček, L.; De Diego, N.; Kopečná, M.; et al. Diphenylurea-derived cytokinin oxidase/dehydrogenase inhibitors for biotechnology and agriculture. J. Exp. Bot. 2020. [Google Scholar] [CrossRef] [PubMed]
- Szczepanek, M.; Jaśkiewicz, B.; Kotwica, K. Response of barley on seaweed biostimulant application. Res. Rural Dev. 2018, 2. [Google Scholar] [CrossRef]
- Zahir, Z.; Asghar, H.; Arshad, M. Cytokinin and its precursors for improving growth and yield of rice. Soil Biol. Biochem. 2001, 33, 405–408. [Google Scholar] [CrossRef]
- Yang, D.; Li, Y.; Shi, Y.; Cui, Z.; Luo, Y.; Zheng, M.; Chen, J.; Li, Y.; Yin, Y.; Wang, Z. Exogenous Cytokinins Increase Grain Yield of Winter Wheat Cultivars by Improving Stay-Green Characteristics under Heat Stress. PLoS ONE 2016, 11, e0155437. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gu, D.; Ding, Y.; Wang, Q.; Li, G.; Wang, S. The relationship between nitrogen, auxin and cytokinin in the growth regulation of rice (Oryza satival.) Tiller buds. Aust. J. Crop Sci. 2011, 5, 1019. [Google Scholar]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef]
- Kumari, A.; Baskaran, P.; Plačková, L.; Omámiková, H.; Nisler, J.; Doležal, K.; Van Staden, J. Plant growth regulator in-teractions in physiological processes for controlling plant regeneration and in vitro development of Tulbaghia simmleri. J. Plant Physiol. 2018, 223, 65–71. [Google Scholar] [CrossRef]
- Sakakibara, H. Cytokinin biosynthesis and metabolism. In Plant Hormones; Springer: Dordrecht, the Netherlands, 2010; pp. 95–114. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| A | Samples | Total Cytokinins | |||||||
| CTRL | 121.79 | ± | 16.35 | ||||||
| RR-O | 105.94 | ± | 18.64 | ||||||
| RR-G | 90.47 | ± | 11.10 | * | |||||
| RR-V | 117.97 | ± | 17.75 | ||||||
| RR-P | 95.50 | ± | 8.68 | * | |||||
| CTRL | 114.86 | ± | 15.85 | 2.28 | ± | 0.31 | |||
| RR-O | 99.62 | ± | 18.19 | 2.55 | ± | 0.32 | |||
| RR-G | 82.71 | ± | 10.90 | * | 3.62 | ± | 0.30 | ** | |
| RR-V | 110.58 | ± | 17.63 | 2.46 | ± | 0.14 | |||
| RR-P | 82.33 | ± | 8.68 | * | 3.45 | ± | 0.14 | ** | |
| C | Samples | Total CK Bases | Total CK Ribosides | ||||||
| CTRL | 1.12 | ± | 0.18 | 3.05 | ± | 0.48 | |||
| RR-O | 1.28 | ± | 0.23 | 1.88 | ± | 0.37 | * | ||
| RR-G | 1.66 | ± | 0.32 | * | 1.88 | ± | 0.26 | ** | |
| RR-V | 1.45 | ± | 0.17 | 2.90 | ± | 0.30 | |||
| RR-P | 1.99 | ± | 0.13 | *** | 7.16 | ± | 1.14 | ** | |
| D | Samples | iP | iPR | ||||||
| CTRL | 0.58 | ± | 0.08 | 0.84 | ± | 0.21 | |||
| RR-O | 0.77 | ± | 0.16 | 0.50 | ± | 0.12 | * | ||
| RR-G | 1.10 | ± | 0.24 | * | 0.48 | ± | 0.12 | * | |
| RR-V | 0.88 | ± | 0.20 | 0.67 | ± | 0.15 | |||
| RR-P | 1.20 | ± | 0.12 | *** | 1.78 | ± | 0.40 | * | |
| E | Samples | tZ | tZR | ||||||
| CTRL | 0.34 | ± | 0.08 | 0.18 | ± | 0.02 | |||
| RR-O | 0.32 | ± | 0.08 | 0.16 | ± | 0.02 | |||
| RR-G | 0.37 | ± | 0.07 | 0.19 | ± | 0.04 | |||
| RR-V | 0.34 | ± | 0.07 | 0.18 | ± | 0.02 | |||
| RR-P | 0.41 | ± | 0.03 | 0.28 | ± | 0.03 | ** | ||
| F | Samples | cZ | cZR | ||||||
| CTRL | 0.20 | ± | 0.03 | 2.02 | ± | 0.38 | |||
| RR-O | 0.19 | ± | 0.02 | 1.22 | ± | 0.24 | * | ||
| RR-G | 0.19 | ± | 0.04 | 1.21 | ± | 0.14 | * | ||
| RR-V | 0.23 | ± | 0.03 | 2.06 | ± | 0.17 | |||
| RR-P | 0.38 | ± | 0.03 | *** | 5.10 | ± | 0.76 | *** | |
| Wheat | Barley | ||||||||||
| 2020 | Ctrl | RR-O | RR-V | Ctrl | RR-O | RR-V | |||||
| Ctrl | 0.967 | 1.000 | Ctrl | 0.406 | 1.000 | ||||||
| RR-O | 0.967 | 1.000 | RR-O | 0.406 | 0.714 | ||||||
| RR-V | 1.000 | 1.000 | RR-V | 1.000 | 0.714 | ||||||
| 2019 | Ctrl | RR-O | RR-V | Ctrl | RR-V | ||||||
| Ctrl | 1.000 | 1.000 | Ctrl | 0.462 | |||||||
| RR-O | 1.000 | 1.000 | RR-V | 0.462 | |||||||
| RR-V | 1.000 | 1.000 | |||||||||
| 2018 | Ctrl | RR-V | Ctrl | RR-O | RR-V | ||||||
| Ctrl | 0.935 | Ctrl | 1.000 | 1.000 | |||||||
| RR-V | 0.935 | RR-O | 1.000 | 1.000 | |||||||
| RR-V | 1.000 | 1.000 | |||||||||
| 2017 | Ctrl | RR-P | RR-G | RR-O | RR-V | Ctrl | RR-O | RR-V | |||
| Ctrl | 1.000 | 1.000 | 1.000 | 1.000 | Ctrl | 0.143 | 1.000 | ||||
| RR-P | 1.000 | 1.000 | 1.000 | 1.000 | RR-O | 0.143 | 0.804 | ||||
| RR-G | 1.000 | 1.000 | 1.000 | 1.000 | RR-V | 1.000 | 0.804 | ||||
| RR-O | 1.000 | 1.000 | 1.000 | 1.000 | |||||||
| RR-V | 1.000 | 1.000 | 1.000 | 1.000 | |||||||
| 2016 | Ctrl | RR-P | RR-G | Ctrl | RR-O | RR-G | |||||
| Ctrl | 1.000 | 0.714 | Ctrl | 0.718 | 1.000 | ||||||
| RR-P | 1.000 | 1.000 | RR-O | 0.718 | 1.000 | ||||||
| RR-G | 0.714 | 1.000 | RR-G | 1.000 | 1.000 | ||||||
| 2015 | Ctrl | RR-P | RR-G | RR-O | Ctrl | RR-O | RR-P | RR-G | |||
| Ctrl | 0.088 | 0.041 | 1.000 | Ctrl | 0.600 | 1.000 | 1.000 | ||||
| RR-P | 0.088 | 1.000 | 0.831 | RR-O | 0.600 | 1.000 | 1.000 | ||||
| RR-G | 0.041 | 1.000 | 0.447 | RR-P | 1.000 | 1.000 | 1.000 | ||||
| RR-O | 1.000 | 0.831 | 0.447 | RR-G | 1.000 | 1.000 | 1.000 | ||||
| 2014 | Ctrl | RR-P | RR-G | Ctrl | RR-P | RR-G | |||||
| Ctrl | 1.000 | 1.000 | Ctrl | 1.000 | 0.967 | ||||||
| RR-P | 1.000 | 1.000 | RR-P | 1.000 | 1.000 | ||||||
| RR-G | 1.000 | 1.000 | RR-G | 0.967 | 1.000 | ||||||
| 2013 | Ctrl | RR-P | RR-G | ||||||||
| Ctrl | 1.000 | 1.000 | |||||||||
| RR-P | 1.000 | 1.000 | |||||||||
| RR-G | 1.000 | 1.000 | |||||||||
| 2012 | Ctrl | RR-P | Ctrl | RR-P | RR-G | ||||||
| Ctrl | 0.564 | Ctrl | 1.000 | 0.350 | |||||||
| RR-P | 0.564 | RR-P | 1.000 | 0.509 | |||||||
| RR-G | 0.350 | 0.509 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koprna, R.; Humplík, J.F.; Špíšek, Z.; Bryksová, M.; Zatloukal, M.; Mik, V.; Novák, O.; Nisler, J.; Doležal, K. Improvement of Tillering and Grain Yield by Application of Cytokinin Derivatives in Wheat and Barley. Agronomy 2021, 11, 67. https://doi.org/10.3390/agronomy11010067
Koprna R, Humplík JF, Špíšek Z, Bryksová M, Zatloukal M, Mik V, Novák O, Nisler J, Doležal K. Improvement of Tillering and Grain Yield by Application of Cytokinin Derivatives in Wheat and Barley. Agronomy. 2021; 11(1):67. https://doi.org/10.3390/agronomy11010067
Chicago/Turabian StyleKoprna, Radoslav, Jan F. Humplík, Zdeněk Špíšek, Magdaléna Bryksová, Marek Zatloukal, Václav Mik, Ondřej Novák, Jaroslav Nisler, and Karel Doležal. 2021. "Improvement of Tillering and Grain Yield by Application of Cytokinin Derivatives in Wheat and Barley" Agronomy 11, no. 1: 67. https://doi.org/10.3390/agronomy11010067
APA StyleKoprna, R., Humplík, J. F., Špíšek, Z., Bryksová, M., Zatloukal, M., Mik, V., Novák, O., Nisler, J., & Doležal, K. (2021). Improvement of Tillering and Grain Yield by Application of Cytokinin Derivatives in Wheat and Barley. Agronomy, 11(1), 67. https://doi.org/10.3390/agronomy11010067