
Halotolerant-Koccuria rhizophila (14asp)-Induced Amendment of Salt Stress in Pea Plants by Limiting Na+ Uptake and Elevating Production of Antioxidants

 , , ,
, , ,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Salt Stress Analysis
2.2. Bacteria Antibiotic Resistance
2.3. Bacterial Growth Conditions and Pea Seed Inoculations
2.4. Experimental Setup
2.5. Plant Growth Parameters
2.6. Leaf Relative Water Content (RWC)
2.7. Estimation of Leaf Chlorophyll Content
2.8. Antioxidant Assays in Peas
2.9. Proline Assay in Peas
2.10. Mineral Analysis in Pea Shoots
2.11. Recovery of Inoculated Bacteria
2.12. Statistics
3. Results
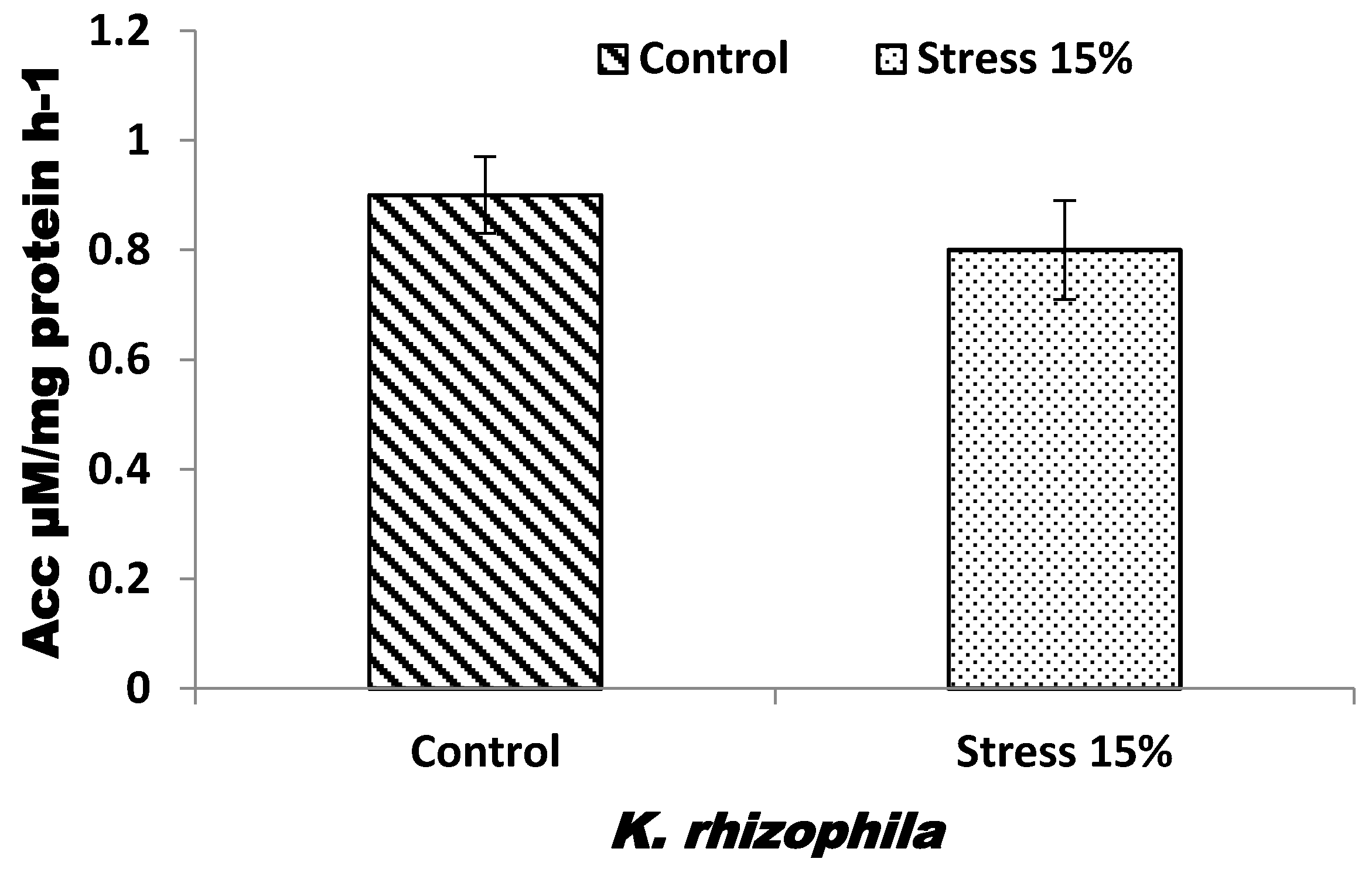
3.1. Bacteria Salt Tolerance
3.2. Plant Growth Parameters
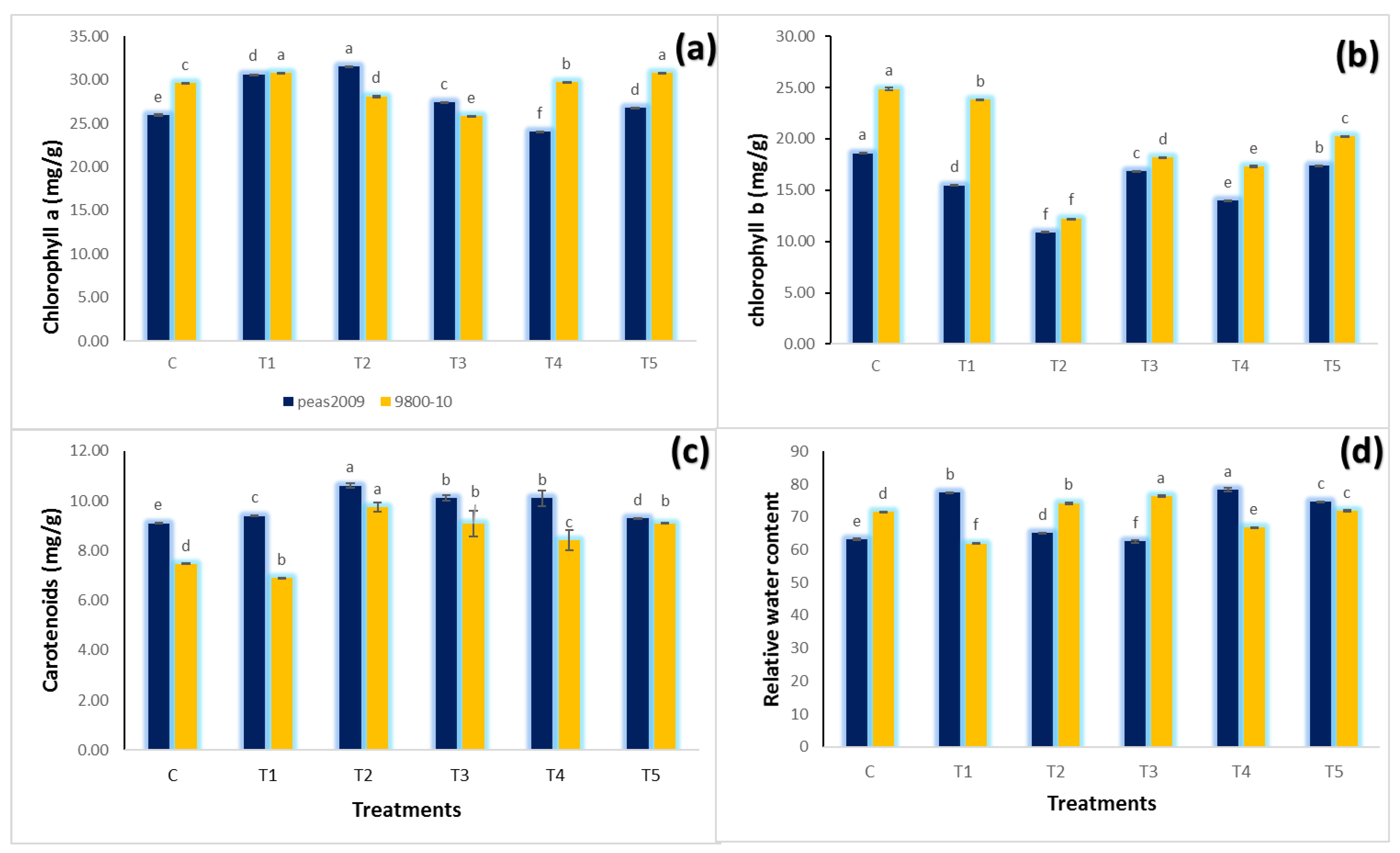
3.3. Leaf Chlorophyll Content
3.4. Leaf Relative Water Contents (RWC)
3.5. Pea Leaf Antioxidants
3.6. Proline
3.7. Inorganic Osmolytes
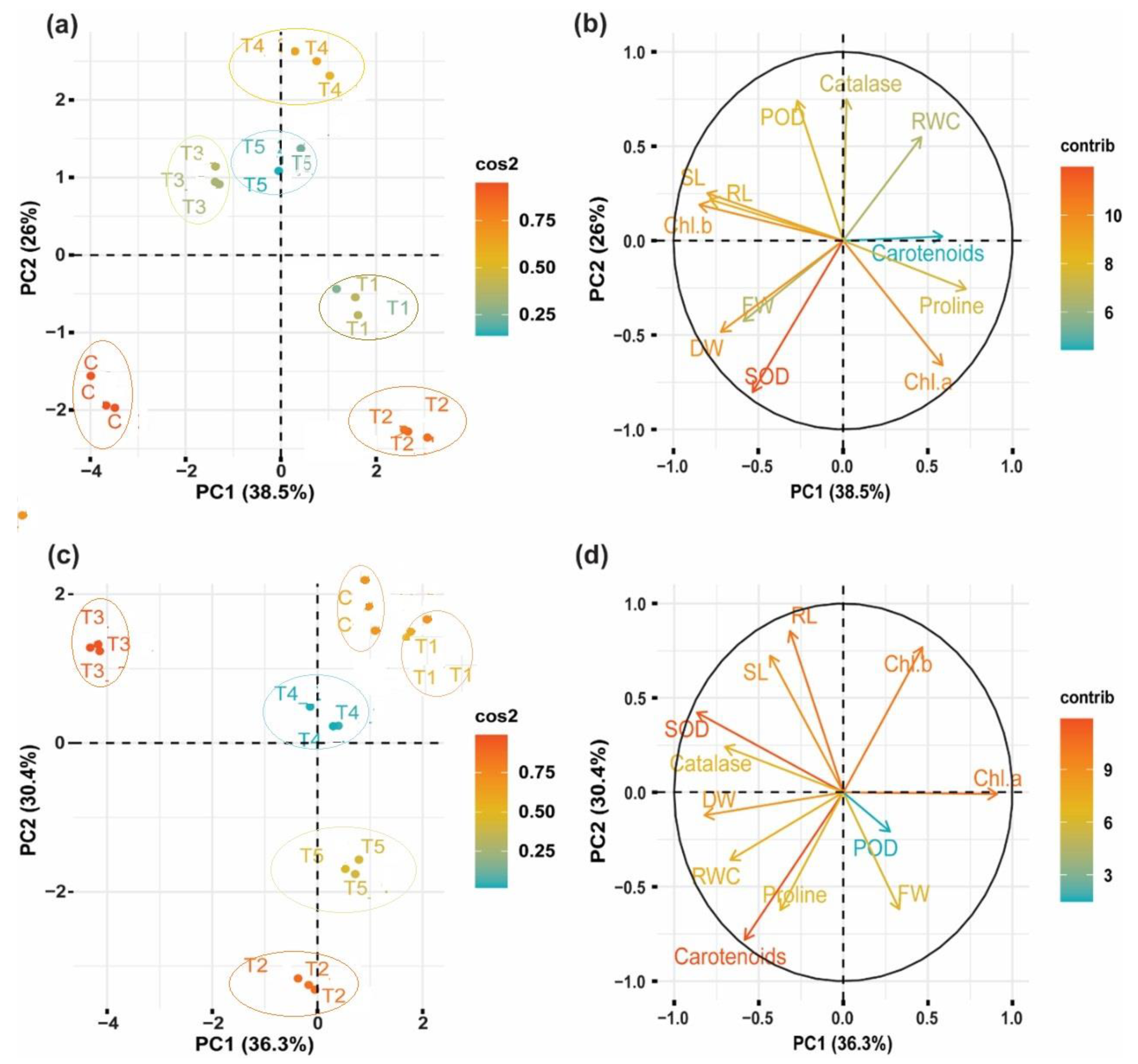
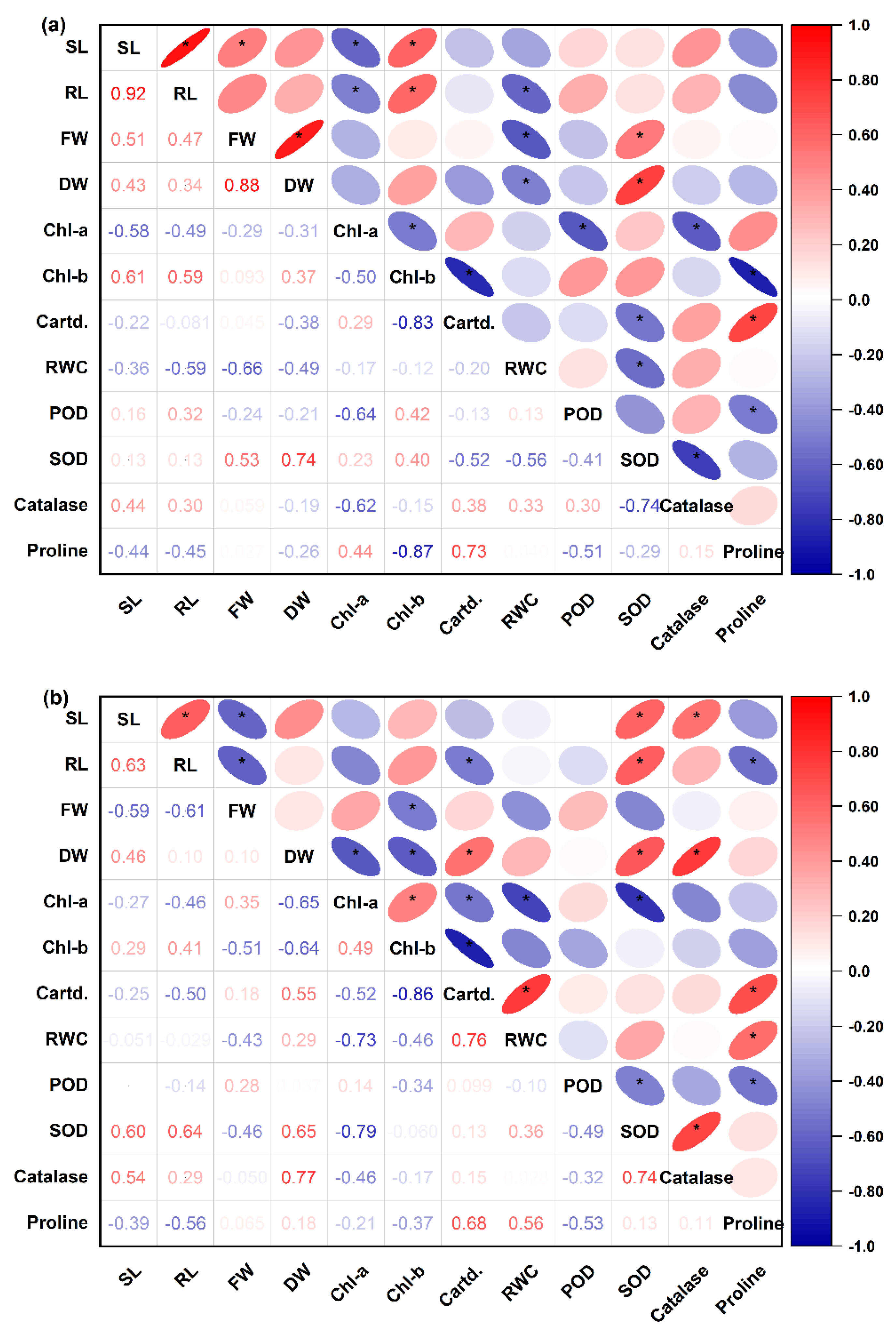
3.8. Principal Component Analysis and Pearson’s Correlation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shaukat, M.; Wu, J.; Fan, M.; Hussain, S.; Yao, J.; Serafim, M.E. Acclimation improves salinity tolerance capacity of pea by modulating potassium ions sequestration. Sci. Hortic. 2019, 254, 193–198. [Google Scholar] [CrossRef]
- Pandolfi, C.; Mancuso, S.; Shabala, S. Physiology of acclimation to salinity stress in pea (Pisum sativum). Environ. Exp. Bot. 2012, 84, 44–51. [Google Scholar] [CrossRef]
- Shannon, M.C. In quest of rapid screening techniques for plant salt tolerance. HortScience 1979, 14, 587–589. [Google Scholar]
- Qados, A.M.S.A. Effect of salt stress on plant growth and metabolism of bean plant Vicia faba (L.). J. Saudi Soc. Agric. Sci. 2011, 10, 7–15. [Google Scholar]
- Ghassemi, F.; Jakeman, A.J.; Nix, H.A. Salinisation of Land and Water Resources: Human Causes, Extent, Management and Case Studies; CAB International: Wallingford, UK, 1995. [Google Scholar]
- Hernández, J.A.; Río, L.A.D.; Sevilla, F. Salt stress-induced changes in superoxide dismutase isozymes in leaves and mesophyll protoplasts from Vigna unguiculata (L.) Walp. New Phytol. 2006, 126, 37–44. [Google Scholar] [CrossRef]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef]
- Hampson, C.R.; Simpson, G.M. Effects of temperature, salt, and osmotic potential on early growth of wheat (Triticum aestivum). I. Germination. Can. J. Bot. 1990, 68, 524–528. [Google Scholar] [CrossRef]
- Cordovilla, M.P.; Ocana, A.; Ligero, F.; Lluch, C. Salinity effects on growth analysis and nutrient composition in four grain legumes-rhizobium symbiosis. J. Plant Nutr. 1995, 18, 1595–1609. [Google Scholar] [CrossRef]
- Nikolopoulou, D.; Grigorakis, K.; Stasini, M.; Alexis, M.N.; Iliadis, K. Differences in chemical composition of field pea (Pisum sativum) cultivars: Effects of cultivation area and year. Food Chem. 2007, 103, 847–852. [Google Scholar] [CrossRef]
- Rao, G.V.S.; Johansen, C.; Rao, J.V.D.K.K.; Jana, M.K. Salinity Tolerance in F1 Hybrids of Pigeonpea and a Tolerant Wild Relative. Crop Sci. 1990, 30, 785–788. [Google Scholar]
- Najafi, F.; Khavarinejad, R.A.; Rastgarjazii, F.; Sticklen, M. Growth and Some Physiological Attributes of Pea (Pisum sativum L.) As Affected by Salinity. Pak. J. Biol. Sci. 2007, 10, 2752–2755. [Google Scholar]
- Vinocur, B.; Altman, A. Recent advances in engineering plant tolerance to abiotic stress: Achievements and limitations. Curr. Opin. Biotechnol. 2005, 16, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Mateo, A. LESION SIMULATING DISEASE 1 Is Required for Acclimation to Conditions That Promote Excess Excitation Energy. Plant Physiol. 2004, 136, 2818–2830. [Google Scholar] [CrossRef]
- Termaat, A.; Munns, R. Use of Concentrated Macronutrient Solutions to Separate Osmotic from NaCl-specific Effects on Plant Growth. Funct. Plant Biol. 1986, 13, 509–522. [Google Scholar] [CrossRef]
- Silberbush, M.; Ben-Asher, J. Simulation study of nutrient uptake by plants from soilless cultures as affected by salinity buildup and transpiration. Plant Soil 2001, 233, 59–69. [Google Scholar] [CrossRef]
- Xu, G.; Magen, H.; Tarchitzky, J.; Kafkafi, U. Advances in Chloride Nutrition of Plants. Adv. Agron. 2000, 68, 97–110. [Google Scholar]
- Cuin, T.A.; Shabala, S. Compatible solutes reduce ROS-induced potassium efflux in Arabidopsis roots. Plant Cell Environ. 2007, 30, 875–885. [Google Scholar] [CrossRef]
- Chen, Z.; Pottosin, I.I.; Cuin, T.A.; Fuglsang, A.T.; Tester, M.; Jha, D.; Zepeda-Jazo, I.; Zhou, M.; Palmgren, M.G.; Newman, I.A.; et al. Root Plasma Membrane Transporters Controlling K+/Na+ Homeostasis in Salt-Stressed Barley. Plant Physiol. 2007, 145, 1714–1725. [Google Scholar] [CrossRef]
- Sairam, R.K.; Srivastava, G.C. Changes in antioxidant activity in sub-cellular fractions of tolerant and susceptible wheat genotypes in response to long term salt stress. Plant Sci. 2002, 162, 897–904. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Demidchik, V.; Shabala, S.N.; Davies, J.M. Spatial variation in H2O2 response of Arabidopsis thaliana root epidermal Ca2+ flux and plasma membrane Ca2+ channels. Plant J. Cell Mol. Biol. 2007, 49, 377–386. [Google Scholar] [CrossRef]
- Shabala, S. Salinity and programmed cell death: Unravelling mechanisms for ion specific signalling. J. Exp. Bot. 2009, 60, 709–712. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef]
- Jimenez, A.; Hernandez, J.A.; Rio, L.A.D.; Sevilla, F. Evidence for the Presence of the Ascorbate-Glutathione Cycle in Mitochondria and Peroxisomes of Pea Leaves. Plant Physiol. 1997, 114, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Kerepesi, I.; Galiba, G. Osmotic and Salt Stress-Induced Alteration in Soluble Carbohydrate Content in Wheat Seedlings. Crop Sci. 2000, 40, 482. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Ghoulam, C.; Foursy, A.; Fares, K. Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environ. Exp. Bot. 2002, 47, 39–50. [Google Scholar] [CrossRef]
- Benlloch-González, M.; Fournier, J.M.; Ramos, J.; Benlloch, M. Strategies underlying salt tolerance in halophytes are present in Cynara cardunculus. Plant Sci. 2005, 168, 653–659. [Google Scholar] [CrossRef]
- Adriano, S.; Antonio, S.; Maria, N.; Antonella, V. Ascorbate Peroxidase and Catalase Activities and Their Genetic Regulation in Plants Subjected to Drought and Salinity Stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar]
- ALKahtani, M.D.; Fouda, A.; Attia, K.A.; Al-Otaibi, F.; Eid, A.M.; Ewais, E.E.-D.; Hijri, M.; St-Arnaud, M.; Hassan, S.E.-D.; Khan, N. Isolation and characterization of plant growth promoting endophytic bacteria from desert plants and their application as bioinoculants for sustainable agriculture. Agronomy 2020, 10, 1325. [Google Scholar] [CrossRef]
- Stone, J.K.; Bacon, C.W.; White, J. An overview of endophytic microbes: Endophytism defined. Microb. Endophytes 2000, 3, 29–33. [Google Scholar]
- Alfonzo, A.; Piccolo, S.L.; Conigliaro, G.; Ventorino, V.; Burruano, S.; Moschetti, G. Antifungal peptides produced byBacillus amyloliquefaciensAG1 active against grapevine fungal pathogens. Ann. Microbiol. 2012, 62, 1593–1599. [Google Scholar] [CrossRef]
- Li, H.-Y.; Shen, M.; Zhou, Z.-P.; Li, T.; Wei, Y.; Lin, L. Diversity and cold adaptation of endophytic fungi from five dominant plant species collected from the Baima Snow Mountain, Southwest China. Fungal Divers. 2012, 54, 79–86. [Google Scholar] [CrossRef]
- Khan, A.; Sirajuddin; Xue, Q.Z.; Javed, M.T.; Masood, S. Bacillus pumilus enhances tolerance in rice (Oryza sativa L.) to combined stresses of NaCl and high boron due to limited uptake of Na+. Environ. Exp. Bot. 2016, 124, 120–129. [Google Scholar] [CrossRef]
- Afridi, M.S.; Amna; Sumaira; Mahmood, T.; Salam, A.; Mukhtar, T.; Mehmood, S.; Ali, J.; Khatoon, Z.; Bibi, M.; et al. Induction of tolerance to salinity in wheat genotypes by plant growth promoting endophytes: Involvement of ACC deaminase and antioxidant enzymes. Plant Physiol. Biochem. 2019, 3, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Charles, T.C.; Glick, B.R. Amelioration of high salinity stress damage by plant growth-promoting bacterial endophytes that contain ACC deaminase. Plant Physiol. Biochem. 2014, 80, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res. Microbiol. 2017, 169, 20–32. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Ahmad, N.; Hussain, J.; Kang, S.M.; Kim, Y.H.; Adnan, M.; Tang, D.S.; Waqas, M.; Radhakrishnan, R. Salinity stress resistance offered by endophytic fungal interaction between Penicillium minioluteum LHL09 and Glycine max. L. J. Microbiol. Biotechnol. 2011, 21, 893–902. [Google Scholar] [CrossRef]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.-P. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- Arshad, M.; Shaharoona, B.; Mahmood, T. Inoculation with Pseudomonas spp. Containing ACC-Deaminase Partially Eliminates the Effects of Drought Stress on Growth, Yield, and Ripening of Pea (Pisum sativum L.). Pedosphere 2008, 18, 611–620. [Google Scholar] [CrossRef]
- Marulanda, A.; Azcón, R.; Chaumont, F.; Ruiz-Lozano, J.M.; Aroca, R. Regulation of plasma membrane aquaporins by inoculation with a Bacillus megaterium strain in maize (Zea mays L.) plants under unstressed and salt-stressed conditions. Planta 2010, 232, 533–543. [Google Scholar] [CrossRef]
- Egamberdiyeva, D.; Hflich, G. Influence of growth-promoting bacteria on the growth of wheat in different soils and temperatures. Soil Biol. Biochem. 2003, 35, 973–978. [Google Scholar] [CrossRef]
- Ait Barka, E.; Nowak, J.; Clement, C. Enhancement of Chilling Resistance of Inoculated Grapevine Plantlets with a Plant Growth-Promoting Rhizobacterium, Burkholderia phytofirmans Strain PsJN. Appl. Environ. Microbiol. 2006, 72, 7246–7252. [Google Scholar] [CrossRef]
- Figueiredo, M.D.V.B.; Seldin, L.; Araujo, F.F.D.; Mariano, R.D.L.R. Plant Growth Promoting Rhizobacteria: Fundamentals and Applications; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Mufti, R.; Amna; Rafique, M.; Haq, F.; Chaudhury, H.J. Genetic diversity and metal resistance assessment of endophytes isolated from Oxalis corniculata. Soil Environ. 2015, 34, 89–99. [Google Scholar]
- Haq, F.; Butt, M.; Ali, H.; Chaudhary, H.J. Biosorption of cadmium and chromium from water by endophytic Kocuria rhizophila: Equilibrium and kinetic studies. Desalination Water Treat. 2016, 57, 19946–19958. [Google Scholar] [CrossRef]
- Hussain, A.; Amna; Kamran, M.A.; Javed, M.T.; Hayat, K.; Farooq, M.A.; Ali, N.; Ali, M.; Manghwar, H.; Jan, F. Individual and combinatorial application of Kocuria rhizophila and citric acid on phytoextraction of multi-metal contaminated soils by Glycine max L. Environ. Exp. Bot. 2019, 159, 23–33. [Google Scholar] [CrossRef]
- Din, B.U.; Sarfraz, S.; Xia, Y.; Kamran, M.A.; Javed, M.T.; Sultan, T.; Munis, M.F.H.; Chaudhary, H.J. Mechanistic elucidation of germination potential and growth of wheat inoculated with exopolysaccharide and ACC-deaminase producing Bacillus strains under induced salinity stress. Ecotoxicol. Environ. Saf. 2019, 183, 109466. [Google Scholar]
- Ahmad, I.; Akhtar, M.J.; Asghar, H.N.; Ghafoor, U.; Shahid, M. Differential Effects of Plant Growth-Promoting Rhizobacteria on Maize Growth and Cadmium Uptake. J. Plant Growth Regul. 2015, 35, 303–315. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Obini, M.; Ugoji, E.O. Comparison of plant growth-promotion with Pseudomonas aeruginosa and Bacillus subtilis in three vegetables. Braz. J. Microbiol. 2008, 39, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Bano, A. Role of plant growth promoting rhizobacteria and Ag-nano particle in the bioremediation of heavy metals and maize growth under municipal wastewater irrigation. Int. J. Phytoremediation 2016, 18, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Sheng, X.; He, L.; Huang, Z.; Zhang, W. Effects of root inoculation with bacteria on the growth, Cd uptake and bacterial communities associated with rape grown in Cd-contaminated soil. J. Hazard. Mater. 2013, 244, 709–717. [Google Scholar] [CrossRef]
- Noreen, S.; Ashraf, M. Alleviation of adverse effects of salt stress on sunflower (Helianthus annuus L.) by exogenous application of salicylic acid: Growth and photosynthesis. Pak. J. Bot 2008, 40, 1657–1663. [Google Scholar]
- Balestri, M.; Bottega, S.; Spanò, C. Response of Pteris vittata to different cadmium treatments. Acta Physiol. Plant. 2014, 36, 767–775. [Google Scholar] [CrossRef]
- Shankar, V.; Kumar, D.; Agrawal, V. Assessment of antioxidant enzyme activity and mineral nutrients in response to NaCl stress and its amelioration through glutathione in chickpea. Appl. Biochem. Biotechnol. 2016, 178, 267–284. [Google Scholar] [CrossRef]
- Lum, M.; Hanafi, M.; Rafii, Y.; Akmar, A. Effect of drought stress on growth, proline and antioxidant enzyme activities of upland rice. J. Anim. Plant Sci. 2014, 24, 1487–1493. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ansari, M.; Shekari, F.; Mohammadi, M.H.; Juhos, K.; Végvári, G.; Biró, B. Salt-tolerant plant growth-promoting bacteria enhanced salinity tolerance of salt-tolerant alfalfa (Medicago sativa L.) cultivars at high salinity. Acta Physiol. Plant. 2019, 41, 195. [Google Scholar] [CrossRef]
- Halo, B.A.; Khan, A.L.; Waqas, M.; Al-Harrasi, A.; Hussain, J.; Ali, L.; Adnan, M.; Lee, I.-J. Endophytic bacteria (Sphingomonas sp. LK11) and gibberellin can improve Solanum lycopersicum growth and oxidative stress under salinity. J. Plant Interact. 2015, 10, 117–125. [Google Scholar] [CrossRef]
- Abd_Allah, E.F.; Alqarawi, A.A.; Hashem, A.; Radhakrishnan, R.; Al-Huqail, A.A.; Al-Otibi, F.O.N.; Malik, J.A.; Alharbi, R.I.; Egamberdieva, D. Endophytic bacterium Bacillus subtilis (BERA 71) improves salt tolerance in chickpea plants by regulating the plant defense mechanisms. J. Plant Interact. 2018, 13, 37–44. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Wirth, S.; Egamberdieva, D. The Interaction between Arbuscular Mycorrhizal Fungi and Endophytic Bacteria Enhances Plant Growth of Acacia gerrardii under Salt Stress. Front. Microbiol. 2016, 7, 1089. [Google Scholar] [CrossRef] [PubMed]
- Ali, Z.; Ullah, N.; Naseem, S.; Haq, M.I.U.; Jacobsen, H.J. Soil bacteria conferred a positive relationship and improved salt stress tolerance in transgenic pea (Pisum sativum L.) harboring Na+/H+ antiporter. Turk. J. Bot. 2015, 39, 962–972. [Google Scholar] [CrossRef]
- Bertrand, A.; Dhont, C.; Bipfubusa, M.; Chalifour, F.-P.; Drouin, P.; Beauchamp, C.J. Improving salt stress responses of the symbiosis in alfalfa using salt-tolerant cultivar and rhizobial strain. Appl. Soil Ecol. 2015, 87, 108–117. [Google Scholar] [CrossRef]
- Bano, A.; Fatima, M. Salt tolerance in Zea mays (L). following inoculation with Rhizobium and Pseudomonas. Biol. Fertil. Soils 2009, 45, 405–413. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Sankar, B.; Sridharan, R.; Panneerselvam, R. Soil salinity alters growth, chlorophyll content, and secondary metabolite accumulation in Catharanthus roseus. Turk. J. Biol. 2008, 32, 79–83. [Google Scholar]
- Anower, M.R.; Mott, I.W.; Peel, M.D.; Wu, Y. Characterization of physiological responses of two alfalfa half-sib families with improved salt tolerance. Plant Physiol. Biochem. 2013, 71, 103–111. [Google Scholar] [CrossRef]
- Heidari, M.; Golpayegani, A. Effects of water stress and inoculation with plant growth promoting rhizobacteria (PGPR) on antioxidant status and photosynthetic pigments in basil (Ocimum basilicum L.). J. Saudi Soc. Agric. Sci. 2012, 11, 57–61. [Google Scholar] [CrossRef]
- Han, H.; Lee, K. Plant growth promoting rhizobacteria effect on antioxidant status, photosynthesis, mineral uptake and growth of lettuce under soil salinity. Res. J. Agric. Biol. Sci. 2005, 1, 210–215. [Google Scholar]
- Hosseinzadah, F.; Satei, A.; Ramezanpour, M.R. Effects of mycorhiza and plant growth promoting rhizobacteria on growth, nutrients uptake and physiological characteristics in Calendula officinalis L. Middle East J. Sci. Res. 2011, 8, 947–953. [Google Scholar]
- Habib, S.H.; Kausar, H.; Saud, H.M. Plant growth-promoting rhizobacteria enhance salinity stress tolerance in okra through ROS-scavenging enzymes. BioMed Res. Int. 2016, 2016, 6284547. [Google Scholar] [CrossRef]
- Xu, H.-C.; Tie, C.; Wang, Z.-L.; HE, M.-R. Physiological basis for the differences of productive capacity among tillers in winter wheat. J. Integr. Agric. 2015, 14, 1958–1970. [Google Scholar] [CrossRef]
- Akram, M.S.; Shahid, M.; Tariq, M.; Azeem, M.; Javed, M.T.; Saleem, S.; Riaz, S. Deciphering Staphylococcus sciuri SAT-17 mediated anti-oxidative defense mechanisms and growth modulations in salt stressed maize (Zea mays L.). Front. Microbiol. 2016, 7, 867. [Google Scholar] [CrossRef]
- Han, Q.-Q.; Lü, X.-P.; Bai, J.-P.; Qiao, Y.; Paré, P.W.; Wang, S.-M.; Zhang, J.-L.; Wu, Y.-N.; Pang, X.-P.; Xu, W.-B.; et al. Beneficial soil bacterium Bacillus subtilis (GB03) augments salt tolerance of white clover. Front. Plant Sci. 2014, 5, 525. [Google Scholar] [CrossRef] [PubMed]
- Sziderics, A.; Rasche, F.; Trognitz, F.; Sessitsch, A.; Wilhelm, E. Bacterial endophytes contribute to abiotic stress adaptation in pepper plants (Capsicum annuum L.). Can. J. Microbiol. 2007, 53, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Younesi, O.; Moradi, A. Effects of plant growth-promoting rhizobacterium (PGPR) and arbuscular mycorrhizal fungus (AMF) on antioxidant enzyme activities in salt-stressed bean (Phaseolus vulgaris L.). Agriculture (Pol’nohospodárstvo) 2014, 60, 10–21. [Google Scholar] [CrossRef]
- Volkov, V. Salinity tolerance in plants. Quantitative approach to ion transport starting from halophytes and stepping to genetic and protein engineering for manipulating ion fluxes. Front. Plant Sci. 2015, 6, 873. [Google Scholar] [CrossRef] [PubMed]
- Baset, M.; Shamsuddin, Z.; Wahab, Z.; Marziah, M. Effect of plant growth promoting rhizobacterial (PGPR) inoculation on growth and nitrogen incorporation of tissue-cultured’musa’plantlets under nitrogen-free hydroponics condition. Aust. J. Crop. Sci. 2010, 4, 85. [Google Scholar]
- Dodd, I.C.; Pérez-Alfocea, F. Microbial amelioration of crop salinity stress. J. Exp. Bot. 2012, 63, 3415–3428. [Google Scholar] [CrossRef]
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci. Rep. 2016, 6, 34768. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Zou, Y.-N.; He, X.-H. Contributions of arbuscular mycorrhizal fungi to growth, photosynthesis, root morphology and ionic balance of citrus seedlings under salt stress. Acta Physiol. Plant. 2010, 32, 297–304. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | NaCl Concentration | Inoculation |
|---|---|---|
| Control | ✗ | ✗ |
| T1 | 75 mM | ✗ |
| T2 | 150 mM | ✗ |
| T3 | ✗ | ✓ |
| T4 | 75 mM | ✓ |
| T5 | 150 mM | ✓ |
| Treatments | SL (cm) | RL (cm) | FW (g) | DW (g) | ||||
|---|---|---|---|---|---|---|---|---|
| peas2009 | 9800-10 | peas2009 | 9800-10 | peas2009 | 9800-10 | peas2009 | 9800-10 | |
| Control | 18.67 ± 0.16 b | 14.5 ± 0.76 c | 17.5 ± 0.28 b | 23.33 ± 0.66 b | 3.29 ± 0.07 a | 2.54 ± 0.17 d | 0.34 ± 0.02 a | 0.36 ± 0.01 e |
| T1 | 14.33 ± 0.33 d | 14.05 ± 0.5 d | 11.67 ± 0.66 e | 21.16 ± 0.6 ac | 2.5 ± 0.08 e | 2.4 ± 0.14 e | 0.15 ± 0.01 d | 0.29 ± 0.02 d |
| T2 | 11.18 ± 0.28 f | 12.33 ± 0.33 f | 11 ± 0.57 f | 11.4 ± 0.8 e | 1.69 ± 0.19 b | 2.18 ± 0.16 f | 0.17 ± 0.02 b | 0.22 ± 0.03 c |
| T3 | 19.46 ± 0.33 a | 15.83 ± 0.16 a | 20.67 ± 0.66 a | 24.67 ± 0.08 a | 4.31 ± 0.10 d | 2.58 ± 0.18 c | 0.41 ± 0.08 cd | 0.42 ± 0.03 a |
| T4 | 16.55 ± 0.28 c | 15.67 ± 0.66 b | 14.33 ± 0.21 c | 22.33 ± 0.33 d | 2.3 ± 0.25 c | 3.21 ± 0.33 a | 0.22 ± 0.03 bc | 0.39 ± 0.01 b |
| T5 | 12.63 ± 0.16 e | 13.83 ± 0.16 e | 12.73 ± 0.8 d | 12.67 ± 0.32 f | 1.8 ± 0.15 e | 3.07 ± 0.14 b | 0.19 ± 0.05 d | 0.31 ± 0.08 d |
| Treatments | Na+ (mg g-1Dw) | K+ (mg g-1Dw) | Ca+ (mg g-1Dw) | Mg+ (mg g-1Dw) | ||||
|---|---|---|---|---|---|---|---|---|
| peas2009 | 9800-10 | peas2009 | 9800-10 | peas2009 | 9800-10 | peas2009 | 9800-10 | |
| Control | 15.84 ± 0.06 c | 16.97 ± 0.03 f | 17.8 ± 0.16 e | 20.68 ± 0.02 d | 18.58 ± 0.9 c | 18.97 ± 0.01 a | 18.47 ± 0.02 b | 28.88 ± 0.01 d |
| T1 | 18.54 ± 0.01 b | 19.68 ± 0.03 e | 17.18 ± 0.15 f | 20.78 ± 0.08 c | 18.91 ± 0.5 b | 17.32 ± 0.1 e | 10.98 ± 0.12 e | 37.12 ± 0.02 c |
| T2 | 23.94 ± 0.02 a | 24.26 ± 0.06 a | 17.83 ± 0.13 d | 19.36 ± 0.06 f | 18.95 ± 0.9 a | 17.85 ± 0.08 d | 10.09 ± 0.16 f | 40.01 ± 0.02 b |
| T3 | 14.05 ± 0.01 e | 20.26 ± 0.09 d | 19.47 ± 0.07 a | 21.39 ± 0.26 b | 18.07 ± 0.13 d | 18.37 ± 0.02 c | 19.35 ± 0.38 a | 45.64 ± 0.03 a |
| T4 | 12.83 ± 0.08 f | 20.32 ± 0.05 c | 20.85 ± 0.05 a | 21.91 ± 0.04 a | 17.49 ± 0.04 f | 18.94 ± 0.08 a | 17.48 ± 0.24 d | 27.86 ± 0.1 e |
| T5 | 15.31 ± 0.01 cd | 22.91 ± 0.02 b | 20.33 ± 0.02 b | 20.51 ± 0.01 e | 17.87 ± 0.37 e | 18.61 ± 0.2 b | 12.08 ± 0.79 c | 27.66 ± 0.02 f |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.A.; Wang, T.; Hussain, T.; Amna; Ali, F.; Shi, F.; Latef, A.A.H.A.; Ali, O.M.; Hayat, K.; Mehmood, S.; et al. Halotolerant-Koccuria rhizophila (14asp)-Induced Amendment of Salt Stress in Pea Plants by Limiting Na+ Uptake and Elevating Production of Antioxidants. Agronomy 2021, 11, 1907. https://doi.org/10.3390/agronomy11101907
Khan AA, Wang T, Hussain T, Amna, Ali F, Shi F, Latef AAHA, Ali OM, Hayat K, Mehmood S, et al. Halotolerant-Koccuria rhizophila (14asp)-Induced Amendment of Salt Stress in Pea Plants by Limiting Na+ Uptake and Elevating Production of Antioxidants. Agronomy. 2021; 11(10):1907. https://doi.org/10.3390/agronomy11101907
Chicago/Turabian StyleKhan, Amir Abdullah, Tongtong Wang, Tayyaba Hussain, Amna, Fayaz Ali, Fuchen Shi, Arafat Abdel Hamed Abdel Latef, Omar M. Ali, Kashif Hayat, Shehzad Mehmood, and et al. 2021. "Halotolerant-Koccuria rhizophila (14asp)-Induced Amendment of Salt Stress in Pea Plants by Limiting Na+ Uptake and Elevating Production of Antioxidants" Agronomy 11, no. 10: 1907. https://doi.org/10.3390/agronomy11101907
APA StyleKhan, A. A., Wang, T., Hussain, T., Amna, Ali, F., Shi, F., Latef, A. A. H. A., Ali, O. M., Hayat, K., Mehmood, S., Zainab, N., Muneer, M. A., Munis, M. F. H., Soliman, M. H., & Chaudhary, H. J. (2021). Halotolerant-Koccuria rhizophila (14asp)-Induced Amendment of Salt Stress in Pea Plants by Limiting Na+ Uptake and Elevating Production of Antioxidants. Agronomy, 11(10), 1907. https://doi.org/10.3390/agronomy11101907