
Difficulties in Potato Pest Control: The Case of Pyrethroids on Colorado Potato Beetle

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. A Brief Historical Overview of Insecticide Use
3. Types of Insecticides Affecting Different Membrane Channels
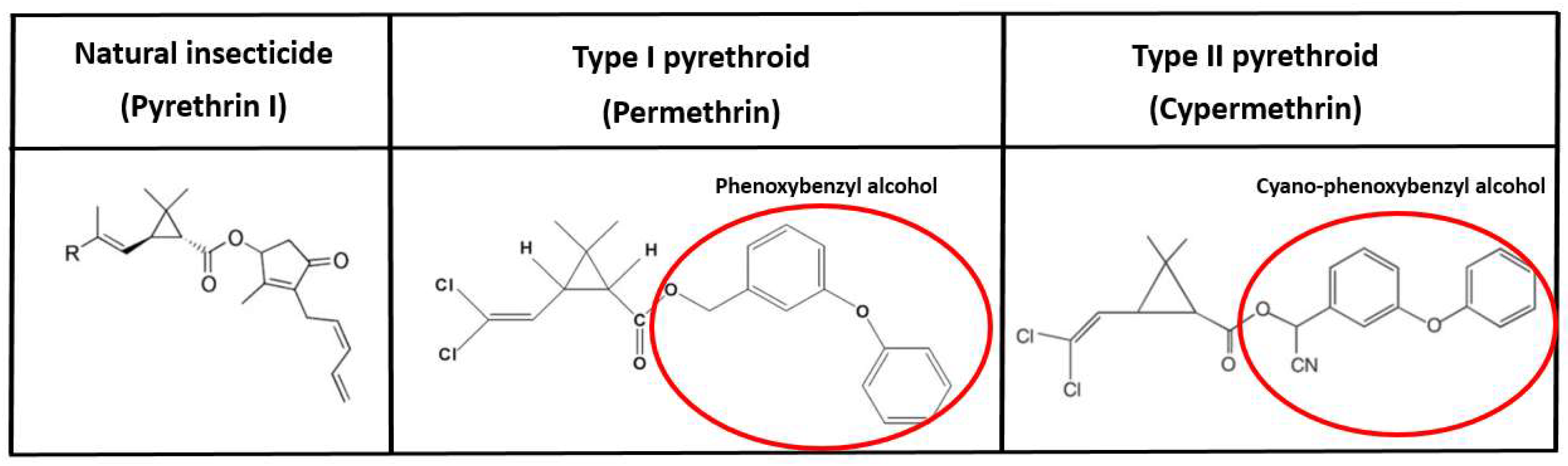
3.1. Pyrethroids
3.2. Toxication Symptoms of Pyrethroids
4. Mechanism of Action of Pyrethroids
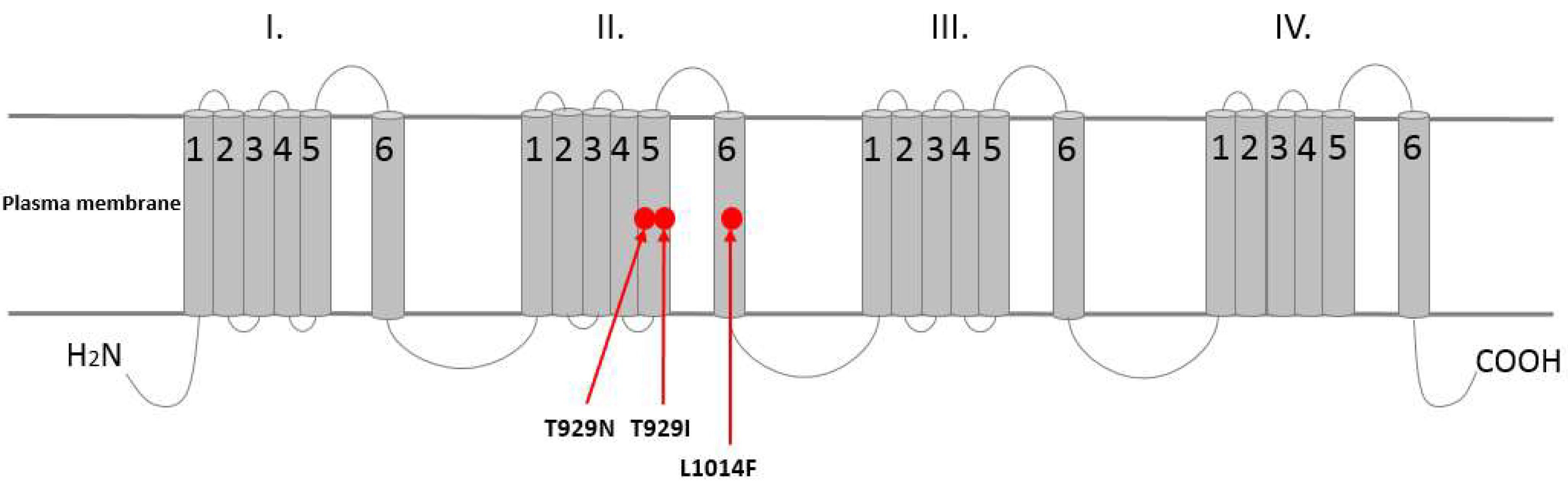
4.1. Pyrethroids Effects on Voltage-Gated Sodium Channels (VGSCs)
4.2. Alternative Targets of Pyrethroids
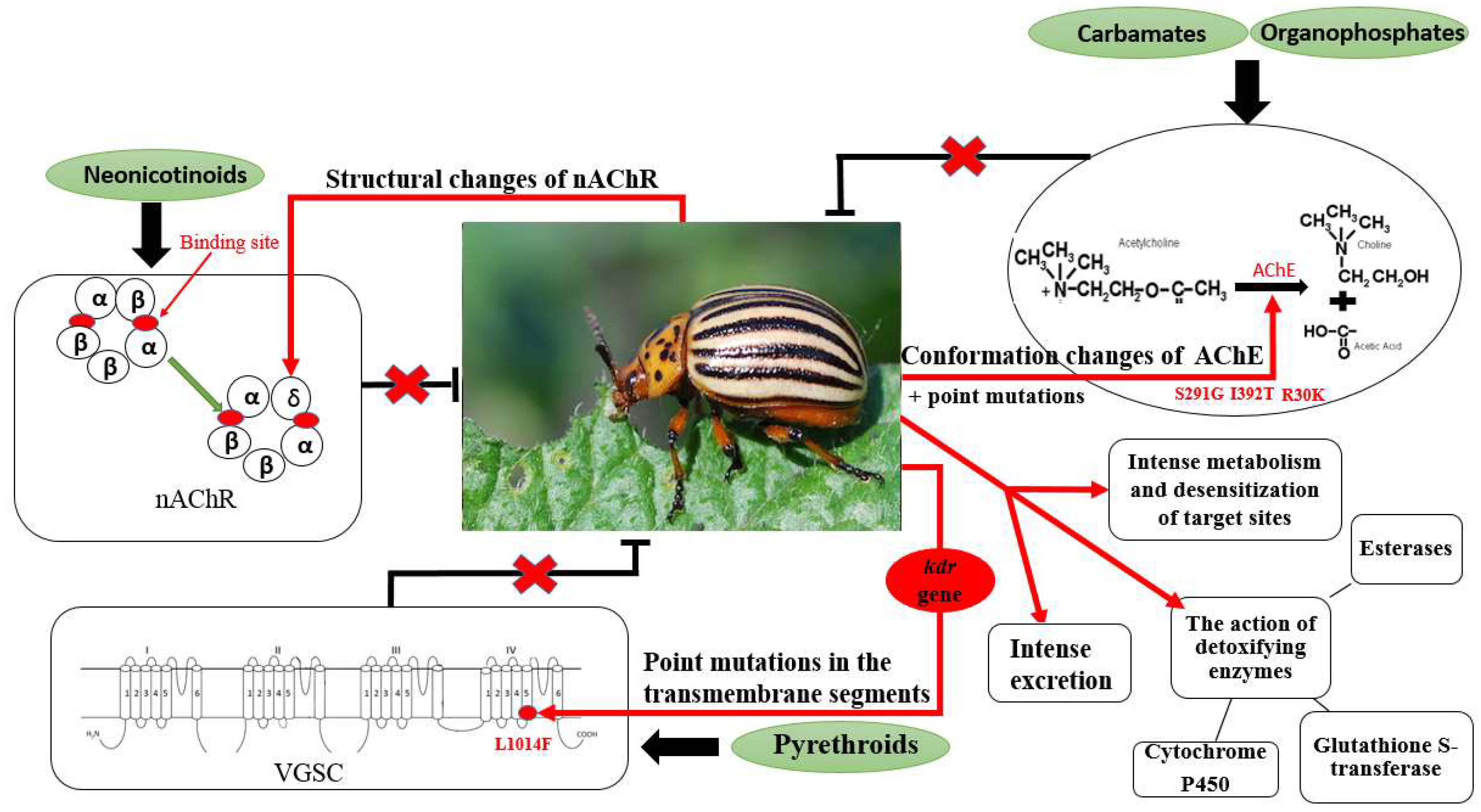
5. Insecticide Resistance—The Case of Colorado Potato Beetle
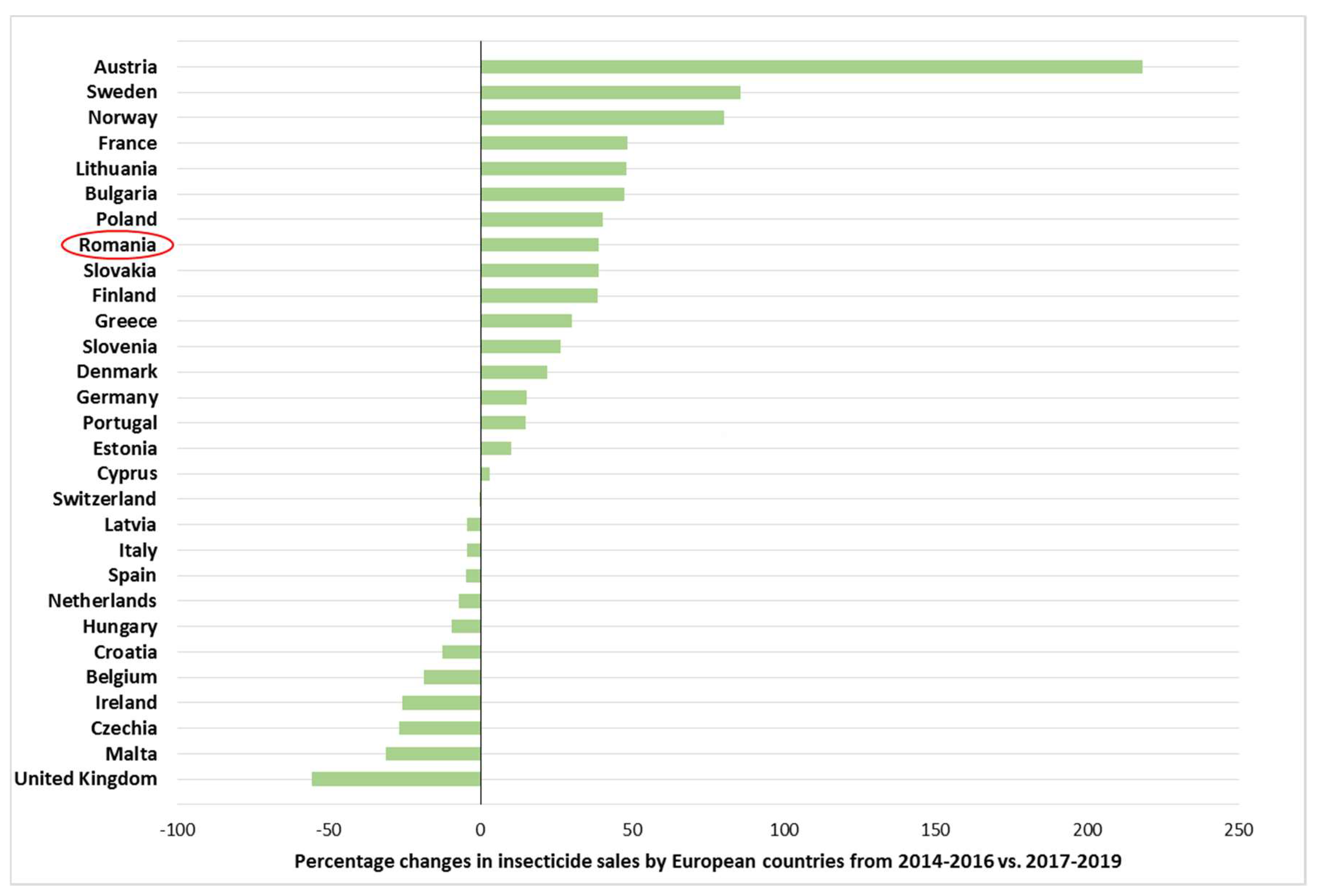
6. The Numbers behind Potato Cultivation and Insecticide Use
7. Perspectives to Reduce Insecticide Use
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobrzanski, Z.; Buszewski, B.; Opalinski, S.; Kolacz, R.; Koziel, J.A. Xenobiotics, toxic compounds, mutagens and carcinogens substances. Classification and regulatory challenges. Chem. Ind. 2017, 96, 76–84. [Google Scholar]
- Tennekes, H.A. The importance of dose-time-response relationships for hazard identification and limitation of animal experiments. Open Acc. J. Toxicol. 2017, 1, 555572. [Google Scholar] [CrossRef]
- European Environment—State and Outlook 2015: Assessment of Global Megatrends, European Environment Agency. Available online: https://www.eea.europa.eu/soer/2015/global/action-download-pdf (accessed on 1 August 2021).
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Balaško, K.M.; Mikac, K.M.; Bažok, R.; Lemic, D. Modern Techniques in Colorado Potato Beetle (Leptinotarsa decemlineata Say) Control and Resistance Management: History Review and Future Perspectives. Insects 2020, 11, 581. [Google Scholar] [CrossRef]
- Oberemok, V.V.; Laikova, K.V.; Gninenko, Y.I.; Zaitsev, A.S.; Nyadar, P.M.; Adeyemi, T.A. A short history of insecticides. J. Plant Prot. Res. 2015, 55, 221–226. [Google Scholar] [CrossRef] [Green Version]
- Casida, J.E.; Durkin, K.A. Anticholinesterase insecticide retrospective. Chem.-Biol. Interact. 2013, 203, 221–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.C. Carbamate Pesticides. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Elsevier Science Publishers: Amsterdam, The Netherlands, 2014; pp. 661–664. [Google Scholar]
- Le Goff, G.; Giraudo, M. Effects of pesticides on the environment and insecticide resistance. In Olfactory Concepts of Insect Control-Alternative to Insecticides; Picimbon, J.F., Ed.; Springer: Cham, Switzerland, 2019; pp. 51–78. [Google Scholar]
- Kaneko, H. Pyrethroid chemistry and metabolism. In Hayes’ Handbook of Pesticide Toxicology, 3rd ed.; Krieger, R., Ed.; Academic Press: Amsterdam, The Netherlands, 2010; pp. 1635–1663. [Google Scholar]
- Soderlund, D.M. Toxicology and mode of action of pyrethroid insecticides. In Hayes’ Handbook of Pesticide Toxicology, 3rd ed.; Krieger, R., Ed.; Academic Press: Amsterdam, The Netherlands, 2010; pp. 1665–1686. [Google Scholar]
- Insecticide Resistance Committee (IRAC). Available online: https://irac-online.org/documents/moa-classification/ (accessed on 15 July 2021).
- Satelle, D.B.; Cordova, D.; Cheek, T.R. Insect ryanodine receptors: Molecular targets for novel pest control chemicals. Rev. Invert. Neurosci. 2008, 8, 107–119. [Google Scholar] [CrossRef]
- Ebbinghaus-Kintscher, U.; Luemmen, P.; Lobitz, N.; Schulte, T.; Funke, C.; Fischer, R.; Masaki, T.; Yasokawa, N.; Tohnishi, M. Phthalic acid diamides activate ryanodine-sensitive Ca2+ release channels in insects. Cell Calcium 2006, 39, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Kadala, A.; Charreton, M.; Collet, C. Flubendiamide, the first phthalic acid diamide insecticide, impairs neuronal calcium signalling in the honey bee’s antennae. J. Insect Physiol. 2020, 125, 104086–104093. [Google Scholar] [CrossRef] [PubMed]
- Buszewski, B.; Bukowska, M.; Ligor, M.; Staneczko-Baranowska, I. A holistic study of neonicotinoids neuroactive insecticides-properties, applications, occurrence, and analysis. Environ. Sci. Pollut. Res. Int. 2019, 26, 34723–34740. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.Y.; Daugherty, L.; Mitchell, A. Bee afraid, bee very afraid—Neonicotinoids and the nAChRs family. InterPro Protein Focus 2013, 25, 1–8. [Google Scholar]
- Bonmatin, J.M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A.D.; et al. Environmental fate and exposure; neonicotinoids and fipronil. Environ. Sci. Pollut. Res. 2015, 22, 35–67. [Google Scholar] [CrossRef]
- European Commision Website for Food Safety. Available online: https://ec.europa.eu/food/plants/pesticides/approval-active-substances/renewal-approval/neonicotinoids_en (accessed on 21 July 2021).
- Peter, J.V.; Sudarsan, T.I.; Moran, J.L. Clinical features of organophosphate poisoning: A review of different classification systems and approaches. Indian J. Crit. Care Med. 2014, 18, 735–745. [Google Scholar] [CrossRef] [Green Version]
- Jenson, L.; Anderson, T.D.; Bloomquist, J.R. Insect Voltage-gated chloride channels as a possible insecticide target site. In Discovery and Synthesis of Crop Protection Products; Maienfisch, P., Stevenson, T.M., Eds.; American Chemical Society: Washington DC, USA, 2015; pp. 447–454. [Google Scholar]
- Bloomquist, J.R. GABA and glutamate receptors as biochemical sites for insecticide action. In Biochemical Sites of Insecticide Action and Resistance; Ishaaya, I., Ed.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 17–41. [Google Scholar]
- Dong, K. Insect sodium channels and insecticide resistance. Invert. Neurosci. 2007, 7, 17–30. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Pesticide Evaluation Scheme, Vector Ecology and Management. Available online: http://apps.who.int/iris/bitstream/handle/10665/44305/9789241599276_eng.pdf;jsessionid=5BEC372017196B11B48CED6AF3B1289E?sequence=1 (accessed on 14 July 2021).
- Elliott, M. Chemicals in insect control. In Pyrethrum Flowers: Production, Chemistry, Toxicology, and Uses; Casida, J.E., Quistad, G.B., Eds.; Oxford University Press: New York, NY, USA, 1995; pp. 3–31. [Google Scholar]
- Nasuti, C.; Cantalamessa, F.; Falcioni, G.; Gabbianelli, R. Different effects of Type I and Type II pyrethroids on erythrocyte plasma membrane properties and enzymatic activity in rats. Toxicology 2003, 191, 233–244. [Google Scholar] [CrossRef]
- Kurihara, N.; Miyamot, J.; Paulson, G.D.; Zeeh, B.; Skidmore, M.W.; Hollingworth, R.M.; Kuiper, H.A. Chirality in synthetic agrochemicals: Bioactivity and safety consideration. Pure Appl. Chem. 1997, 69, 2007–2025. [Google Scholar] [CrossRef]
- Bradberry, S.M.; Cage, S.A.; Proudfoot, A.T.; Vale, J.A. Poisoning due to pyrethroids. Toxicol. Rev. 2005, 24, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Chrustek, A.; Hołyńska-Iwan, I.; Dziembowska, I.; Bogusiewicz, J.; Wróblewski, M.; Cwynar, A.; Olszewska-Słonina, D. Current research on the safety of pyrethroids used as insecticides. Medicina 2018, 54, 61. [Google Scholar] [CrossRef] [Green Version]
- Cárcamo, J.G.; Aguilar, M.N.; Carreño, C.F.; Vera, T.; Arias-Darraz, L.; Figueroa, J.E.; Romero, A.P.; Alvarez, M.; Yañez, A.J. Consecutive emamectin benzoate and deltamethrin treatments affect the expressions and activities of detoxification enzymes in the rainbow trout (Oncorhynchus mykiss) Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2017, 191, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Burridge, L.E.; Haya, K. Lethality of pyrethrins to larvae and postlarvae of the American lobster (Homarus americanus). Ecotoxicol. Environ. Saf. 1997, 38, 150–154. [Google Scholar] [CrossRef]
- Lidova, J.; Stara, A.; Kouba, A.; Velisek, J. The effects of cypermethrin on oxidative stress and antioxidant biomarkers in marbled crayfish (Procambarus fallax f. virginalis). Neuro Endocrinol. Lett. 2016, 37 (Suppl. 1), 53–59. [Google Scholar] [PubMed]
- Lutnicka, H.; Kozińska, A. Pyrethroids as a predisposing factor in fish diseases. Ochr. Środ. Zasobów Nat. 2009, 41, 285–292. [Google Scholar]
- Wang, Y.; Lv, L.; Yu, Y.; Yang, G.; Xu, Z.; Wang, Q.; Cai, L. Single and joint toxic effects of five selected pesticides on the early life stages of zebrafish (Denio renio). Chemosphere 2017, 170, 61–67. [Google Scholar] [CrossRef]
- Miyamoto, J.; Kaneko, H.; Tsuji, R.; Okuno, Y. Pyrethroids, nerve poisons: How their risks to human health should be assessed. Toxicol. Lett. 1995, 82–83, 933–940. [Google Scholar] [CrossRef]
- Scollon, E.J.; Starr, J.M.; Godin, S.J.; DeVito, M.J.; Hughes, M.F. In vitro metabolism of pyrethroid pesticides by rat and human hepatic microsomes and cytochrome P450 isoforms. Drug Metab. Dispos. 2009, 37, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Damalas, C.A.; Eleftherohotinos, I.G. Pesticide Exposure, Safety Issues, and Risk Assessment Indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef]
- Glorennec, P.; Serrano, T.; Fravallo, M.; Warembourg, C.; Monfort, C.; Cordier, S.; Viel, J.; Le Gléau, F.; Le Bot, B.; Chevrier, C. Determinants of children’s exposure to pyrethroid insecticides in western France. Environ. Int. 2017, 104, 76–82. [Google Scholar] [CrossRef]
- Del Prado-Lu, J.L. Insecticide residues in soil, water, and eggplant fruits and farmers’ health effects due to exposure to pesticides. Environ. Health Prev. Med. 2015, 20, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Shafer, T.J.; Meyer, D.A.; Crofton, K.M. Developmental neurotoxicity of pyrethroid insecticides: Critical review and future research needs. Environ. Health Perspect. 2005, 113, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Nasuti, C.; Carloni, M.; Fedeli, D.; Gabbianelli, R.; Di Stefano, A.; Serafina, C.; Silva, I.; Dominques, V.; Ciccocioppo, R. Effects of early life permethrin exposure on spatial working memory and on monoamine levels in different brain areas of pre-senescent rats. Toxicology 2013, 303, 162–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasuti, C.; Fattoretti, P.; Carloni, M.; Fedeli, D.; Ubaldi, M.; Ciccocioppo, R.; Gabbianelli, R. Neonatal exposure to permethrin pesticide causes lifelong fear and spatial learning deficits and alters hippocampal morphology of synapses. J. Neurodev. Dis. 2014, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Outhlote, Y.; Bouchard, M. Urinary metabolities of organophosphate and pyrethroid pesticides and behavioral problems in Canadian children. Environ. Health Perspect. 2013, 121, 1378–1384. [Google Scholar]
- Viel, J.F.; Warembourg, C.; Le Mauer-Idrissi, G.; Lacroix, A.; Limon, G.; Rouget, F.; Monfort, C.; Durand, G.; Cordier, S.; Cherier, C. Pyrethroid insecticide exposure and cognitive developmental disabilities in children: The PELAGIE mother-child cohort. Environ. Int. 2015, 82, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Skolarczyk, J.; Pekar, J.; Nieradko-Iwanicka, B. Immune disorders induced by exposure to pyrethroid insecticides. Postepy Hig. Med. Dośw. 2017, 71, 446–453. [Google Scholar] [CrossRef]
- Corcellas, C.; Feo, M.L.; Torres, J.P.; Malm, O.; Ocampo-Duque, W.; Eljarrat, E.; Barceló, D. Pyrethroids in human breast milk: Occurrence and nursing daily intake estimation. Environ. Int. 2014, 47, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Mauro, R.; Li, Z. Unique insights into the actions of CNS agents: Lessons from studies of Chlorpyrifos and other common pesticides. Cent. Nerv. Syst. Agents Med. Chem. 2007, 7, 183–199. [Google Scholar] [CrossRef]
- Wylie, B.J.; Hauptman, M.; Woolf, A.D.; Goldman, R.H. Insect Repellants during Pregnancy in the Era of the Zika Virus. Obstet. Gynecol. 2016, 128, 1111–1115. [Google Scholar] [CrossRef] [PubMed]
- Central Disease Control. CDC Zika Interim Response Plan. Available online: https://www.cdc.gov/zika/pdfs/zika-draft-interim-conus-plan.pdf (accessed on 2 July 2021).
- Raghavan, M.; Fee, D.; Barkhaus, P.E. Generation and propagation of the action potential. Handb. Clin. Neurol. 2019, 160, 3–22. [Google Scholar]
- Rinkevich, F.D.; Du, Y.; Dong, K. Diversity and convergence of sodium channel mutations involved in resistance to pyrethroids. Pestic. Biochem. Physiol. 2013, 106, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Lipkind, G.M.; Fozzard, H.A. KcsA crystal structure as framework for a molecular model of the Na+ channel pore. Biochemistry 2000, 39, 8161–8170. [Google Scholar] [CrossRef]
- Wang, J.; Ou, S.W.; Wang, Y.J. Distribution and function of voltage-gated sodium channels in the nervous system. Channels 2017, 11, 534–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, L.G. The neurotoxicity of organochlorine and pyrethroid pesticides. Handb. Clin. Neurol 2015, 131, 135–148. [Google Scholar] [PubMed]
- Taylor-Wells, J.; Brooke, B.D.; Bermudez, I.; Jones, A.K. The neonicotinoid imidacloprid, and the pyrethroid deltamethrin, are antagonists of the insect Rdl GABA receptor. J. Neurochem. 2015, 135, 705–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenson, L.J. Characterization of Neuronal Structure and Insecticide Target Site Expression in Spodoptera frugiperda (Sf21) and Anopheles gambiae (Sua1B) Insect Cells. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2013. [Google Scholar]
- Quintavalle, A. Voltage-gated calcium channels in honey bees: Physiological roles and potential targets for insecticides. BioSci. Master Rev. 2013, 1–11. Available online: http://biologie.ens-lyon.fr/ressources/bibliographies/m1-11-12-biosci-reviews-quintavalle-a-1c-m.xml (accessed on 2 July 2021).
- Wakeling, E.N.; Neal, A.P.; Atchison, W.D. Pyrethroids and their effects on ion channels. In Pesticides-Advances in Chemical and Botanical Pesticides; Soundararajan, R.P., Ed.; IntechOpen: London, UK, 2012; pp. 39–66. [Google Scholar]
- Hildebrand, M.E.; McRory, J.E.; Snutch, T.P.; Stea, A. Mammalian voltage-gated calcium channels are potently blocked by the pyrethroid insecticide allethrin. J. Pharmacol. Exp. Ther. 2004, 308, 805–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neal, A.P.; Yuan, Y.; Atchison, W. Allethrin differentially modulates voltage-gated calcium channel subtypes in rat PC12 cells. Toxicol. Sci. 2010, 116, 604–613. [Google Scholar] [CrossRef]
- Soderlund, D.M.; Clark, J.M.; Sheets, L.P.; Mullin, L.S.; Piccirillo, V.J.; Sargent, D.; Stevens, J.T.; Weiner, M.L. Mechanisms of pyrethroid toxicity: Implications for cumulative risk assessment. Toxicology 2002, 171, 3–59. [Google Scholar] [CrossRef]
- Forshaw, P.J.; Lister, T.; Ray, D.E. The role of voltage-gated chloride channels in type II pyrethroid insecticide poisoning. Toxicol. Appl. Pharmacol. 2000, 163, 1–8. [Google Scholar] [CrossRef]
- Breckenridge, C.B.; Holden, L.; Sturgess, N.; Weiner, M.; Sheets, L.; Sargent, D.; Soderlund, D.M.; Choi, J.-S.; Symington, S.; Clark, J.M.; et al. Evidence for separate mechanism of toxicity for type I and type II pyrethroid insecticides. Neurotoxicology 2009, 30, S17–S31. [Google Scholar] [CrossRef]
- Deglise, P.; Grunewald, B.; Gauthier, M. The insecticide imidacloprid is a partial agonist of the nicotinic receptor of honeybee Kenyon cells. Neurosci. Lett. 2002, 321, 13–16. [Google Scholar] [CrossRef]
- Casagrande, R.A. The Colorado potato beetle: 125 years of mismanagement. Bull. Entomol. Soc. 1987, 33, 142–150. [Google Scholar] [CrossRef]
- Alyokhin, A.; Mota-Sanchez, D.; Baker, M.; Snyder, W.E.; Menasha, S.; Whalon, M.; Dively, G.; Moarsi, W.F. Red Queen on a potato field: IPM vs. chemical dependency in Colorado potato beetle control. Pest Manag. Sci. 2015, 71, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Alyokhin, A. Colorado potato beetle management on potatoes: Current challenges and future prospects. Fruit Veg. Cereal Sci. Biotechnol. 2009, 3, 10–19. [Google Scholar]
- Hare, J.D. Ecology and management of the Colorado potato beetle. Annu. Rev. Entomol. 1990, 35, 81–100. [Google Scholar] [CrossRef]
- Senanayake, D.; Ratcliffe, E.B.; Holliday, N.J. Oviposition and diapause behavior in Colorado potato beetle (Coleoptera: Chrysomelidae) populations from east central Minnesota and the valley of the Red River of the North. Environ. Entomol. 2000, 29, 1123–1132. [Google Scholar] [CrossRef] [Green Version]
- Radcliffe, E.B.; Lagnaoui, A. Insect pests in potato. In Potato Biology and Biotechnology: Advances and Perspectives; Elsevier Science BV: Amsterdam, The Netherlands, 2007; pp. 543–567. [Google Scholar]
- Alyokhin, A.; Choban, R. Maturity-dependent mortality of Colorado potato beetle eggs treated with novaluron. Am. J. Potato Res. 2010, 87, 557–560. [Google Scholar] [CrossRef]
- Hsiao, H.T.H. Host specificity, seasonality and bionomics of Leptinotarsa beetles. In Biology of Chrysomelidae; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1988; pp. 581–659. [Google Scholar]
- Udalov, M.B.; Benkovskaya, G.V. Population genetics of the Colorado potato beetle: From genotype to phenotype. Russ. J. Genet. Appl. Res. 2011, 1, 321–333. [Google Scholar] [CrossRef]
- Schoville, S.D.; Chen, Y.H.; Andersson, M.N.; Benoit, J.B.; Bhandari, A.; Bowsher, J.H.; Brevik, K.; Cappelle, K.; Chen, M.J.M.; Childers, A.K.; et al. A model species for agricultural pest genomics: The genome of the Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Sci. Rep. 2018, 8, 1931–1949. [Google Scholar] [CrossRef] [Green Version]
- Alyokhin, A.; Vincent, C.; Giordanengo, P. Insect Pests of Potato: Global Perspectives on Biology and Management, 1st ed.; Accademic Press: Oxford, UK, 2013; pp. 77–96. [Google Scholar]
- Shields, E.J.; Wyman, J.A. Effect of defoliation at specific growth stages on potato yields. J. Econ. Entomol. 1984, 77, 1194–1199. [Google Scholar] [CrossRef]
- Maharijaya, A.; Vosman, B. Managing the Colorado potato beetle; the need for resistance breeding. Euphytica 2015, 204, 487–501. [Google Scholar] [CrossRef] [Green Version]
- Margus, A.; Piiroinen, S.; Lehmann, P.; Tikka, S.; Karvanen, J.; Lindstorm, L. Sublethal pyrethroid insecticide exposure carries positive fitness effects over generations in a pest insect. Sci. Rep. 2019, 9, 11320–11330. [Google Scholar] [CrossRef] [Green Version]
- Alyokhin, A.; Baker, M.; Mota-Sanchez, D.; Dively, G.; Grafius, E. Colorado potato beetle resistance to insecticides. Am. J. Potato Res. 2008, 85, 395–413. [Google Scholar] [CrossRef]
- Bishop, B.A.; Grafius, E.J. Insecticide resistance in the Colorado potato beetle. In Chrysomelidae Biology; Jolivet, P., Cox, M.L., Eds.; SBP Academic Publishing: Amsterdam, The Netherlands, 1996; Volume 1, pp. 355–377. [Google Scholar]
- Sladan, S.; Miroslav, K.; Ivan, S.; Snezana, J.; Petar, K.; Goran, T.; Jevdovic, R. Resistance of colorado potato beetle (Coleoptera: Chrysomelidae) to neonicotinoids, pyrethroids and nereistoxins in Serbia. Rom. Biotechnol. Lett. 2012, 17, 7599–7609. [Google Scholar]
- Scott, I.M.; Tolman, J.H.; MacArthur, D.C. Insecticide resistance and cross-resistance development in Colorado potato beetle Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae) populations in Canada 2008-2011. Pest Manag. Sci. 2015, 71, 712–721. [Google Scholar] [CrossRef]
- Whalon, M.; Mota-Sanchez, D.; Hollingworth, R.; Duynslager, L. Arthropod Pesticide Resistance Database. 2013. Available online: http://www.pesticideresistance.com/ (accessed on 12 April 2021).
- Rondon, S.I.; Feldman, M.; Thompson, A.; Oppedisano, T.; Shrestha, G. Identifying resistance to the Colorado potato beetle (Leptinotarsa decemlineata Say) in potato germplasm: Review update. Front. Agron. 2021, 3, 642189. [Google Scholar] [CrossRef]
- Hemingway, J. The molecular basis of two contrasting metabolic mechanisms of insecticide resistance. Insect Biochem. Mol. Biol. 2000, 30, 1009–1015. [Google Scholar] [CrossRef]
- Zichova, T.; Kocourek, F.; Salava, J.; Nadova, K.; Stara, J. Detection of organophosphate and pyrethroid resistance alleles in Czech Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) populations by molecular methods. Pest Manag. Sci. 2010, 66, 853–860. [Google Scholar]
- Kim, H.J.; Yoon, K.S.; Clark, J.M. Functional analysis of mutations in expressed acetylcholinesterase that result in azinphosmethyl and carbofuran resistance in Colorado potato beetle. Pestic. Biochem. Physiol. 2007, 88, 181–190. [Google Scholar] [CrossRef]
- Zhu, K.Y.; Lee, S.H.; Clark, J.M. A point mutation of acetylcholinesterase associated with azinphosmethyl resistance and reduced fitness in Colorado potato beetle. Pestic. Biochem. Physiol. 1996, 55, 100–108. [Google Scholar] [CrossRef]
- Williamson, M.S.; Martinez-Torres, D.; Hick, C.A.; Devonshire, A.L. Identification of mutations in the housefly para-type sodium channel gene associated with knockdown resistance (kdr) to pyrethroid insecticides. Mol. Gen. Genet. 1996, 252, 51–60. [Google Scholar] [CrossRef]
- Miyazaki, M.; Ohyama, K.; Dunlap, D.Y.; Matsumura, F. Cloning and sequencing of the paratype sodium channel gene from susceptible and kdr-resistant German cockroaches (Blattella germanica) and house fly (Musca domestica). Mol. Gen. Genet. 1996, 252, 61–68. [Google Scholar]
- Dong, K. A single amino acid change in the para sodium channel protein is associated with knockdown-resistance (kdr) to pyrethroid insecticides in German cockroach. Insect Biochem. Mol. Biol. 1997, 27, 93–100. [Google Scholar] [CrossRef]
- Martinez-Torres, D.; Chandre, F.; Williamson, M.S.; Darriet, F.; Berge, J.B.; Devonshire, A.L.; Guillet, P.; Pasteur, N.; Pauron, D. Molecular characterization of pyrethroid knockdown resistance (kdr) in the major malaria vector Anopheles gambiae s. s. Insect Mol. Biol. 1998, 7, 179–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Torres, D.; Foster, S.P.; Field, L.M.; Devonshire, A.L.; Williamson, M.S. A sodium channel point mutation is associated with resistance to DDT and pyrethroid insecticides in the peachpotato aphid, Myzus persicae (Sulzer) (Hemiptera: Aphididae). Insect Mol. Biol. 1999, 8, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Zamojska, J.; Węgorek, P.; Mrowczynski, P. Changes in the Colorado potato beetle (Leptinotarsa decemlineata Say) susceptibility level to pyrethroids and the pest resistance mechanisms to deltamethrin. J. Plant Prot. Res. 2011, 3, 294–299. [Google Scholar] [CrossRef]
- Rinkevich, F.D.; Su, C.; Lazo, T.A.; Hawthorne, D.J.; Tingey, W.M.; Naimov, S.; Scott, J.G. Multiple evolutionary origins of knockdown resistance (kdr) in pyrethroid-resistant Colorado potato beetle, Leptinotarsa decemlineata. Pestic. Biochem. Physiol. 2012, 104, 192–200. [Google Scholar] [CrossRef]
- Węgorek, P. Current status of resistance in Colorado potato beetle (Leptinotarsa decemlineata Say) to selected active substances of insecticides in Poland. J. Plant Prot. Res. 2005, 45, 309–319. [Google Scholar]
- Clements, J.; Schoville, S.; Peterson, N.; Lan, Q.; Groves, R.L. Characterizing Molecular Mechanisms of Imidacloprid Resistance in Select Populations of Leptinotarsa decemlineata in the Central Sands Region of Wisconsin. PLoS ONE 2016, 11, e0147844. [Google Scholar] [CrossRef]
- Kaplanoglu, E.; Chapman, P.; Scott, I.M.; Donly, C. Overexpression of a cytochrome P450 and a UDP-glycosyltransferase is associated with imidacloprid resistance in the Colorado potato beetle, Leptinotarsa decemlineata. Sci. Rep. 2017, 7, 1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Bayo, F.; Tennekes, H.A.; Goka, K. Impact of Systemic Insecticides on Organisms and Ecosystems. In Insecticides-Development of Safer and More Effective Technologies; Trdan, S., Ed.; IntechOpen: London, UK, 2013; pp. 1–45. [Google Scholar]
- European Commission. Sustainable Use of Pesticides. Available online: https://www.eca.europa.eu/Lists/ECADocuments/SR20_05/SR_Pesticides_EN.pdf (accessed on 25 June 2021).
- EUROSTAT. Available online: http://appsso.eurostat.ec.europa.eu/nui/show.do?dataset=aei_fm_salpest09&lang=en (accessed on 4 May 2021).
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/RP (accessed on 11 May 2021).
- Huseth, A.S.; Frost, K.E.; Knuteson, D.L.; Wyman, J.A.; Groves, R.L. Effects of landscape composition and rotation distance on Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) abundance in cultivated potato. Environ. Entomol. 2012, 41, 1553–1564. [Google Scholar] [CrossRef]
- Stoner, K.A. Influence of mulches on the colonization by adults and survival of larvae of the Colorado potato beetle in eggplant. J. Entomol. Sci. 1997, 32, 7–16. [Google Scholar] [CrossRef]
- Hoy, C.W.; Vaught, T.T.; East, D.A. Increasing the effectiveness of spring trap crops for Leptinotarsa decemlineata. Entomol. Exp. Appl. 2000, 96, 193–204. [Google Scholar] [CrossRef]
- Yasar, B.; Gungor, M.A. Determination of life table and biology of Colorado potato beetle, Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae), feeding on five different potato varieties in Turkey. Appl. Entomol. Zool. 2005, 40, 589–596. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.L.; Farnsworth, B.L.; Gudmestad, N.C.; Secor, G.A.; Preston, D.A.; Sowokinos, J.R.; Glznn, M.; Hatterman-Valenti, H. Dakota Diamond: An exceptionally high yielding, cold chipping potato cultivar with long-term storage potential. Am. J. Pot. Res. 2008, 85, 171–182. [Google Scholar] [CrossRef]
- Göldel, B.; Lemic, D.; Bažok, R. Alternatives to Synthetic Insecticides in the Control of the Colorado Potato Beetle (Leptinotarsa decemlineata Say) and Their Environmental Benefits. Agriculture 2020, 10, 611. [Google Scholar] [CrossRef]
- Sablon, L.; Haubruge, E.; Verhegeen, F.J. Consumption of immature stages of Colorado potato beetle by Chrysoperla carnea (Neuroptera: Chrysopidae) larvae in the laboratory. Am. J. Pot. Res. 2013, 90, 51–57. [Google Scholar] [CrossRef]
- Moore, J. Sweeping fields controls some pests. Am. Veg. Grow. 1990, 1, 10–11. [Google Scholar]
- Laguë, C.; Khelifi, M.; Gill, J.; Lacasse, B. Pneumatic and thermal control of Colorado potato beetle. Can. Agric. Eng. 1999, 41, 53–58. [Google Scholar]
- Rondon, S.I.; Pantoja, A.; Hagerty, A.; Horneck, D.A. Ground beetle (Coleoptera: Carabidae) populations in commercial organic and conventional potato production. Fla. Entomol. 2013, 96, 1492–1499. [Google Scholar] [CrossRef]
- Wraight, S.P.; Ramos, M.E. Delayed efficacy of Bacillus thuringensis foliar spray applications against Colorado potato beetle: Impacts of number and timing of applications on larval and next generation adult population. Biol. Control 2015, 83, 51–67. [Google Scholar] [CrossRef]
- Öztürk, H.E.; Güven, Ö.; Karaca, I. Effects of some bioinsecticides and entomopathogenic fungi on Colorado potato beetle (Leptinotarsa decemlineata L.). Commun. Agric. Appl. Biol. Sci. 2015, 80, 205–211. [Google Scholar]
- Scott, I.M.; Jensen, H.; Scott, J.G.; Isman, M.B.; Arnason, J.T.; Philogène, B.J. Botanical insecticides for controlling agricultural pests: Piperamides and the Colorado Potato Beetle Leptinotarsa decemlineata say (Coleoptera: Chrysomelidae). Arch. Insect Biochem. Physiol. 2003, 54, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.M.; Srinivasan, R.; Cervantes, F.A. Occurrence of the carabid beetle, Pterostichus melanarius (Illiger), in potato ecosystems of Idaho and its predatory potential on the Colorado potato beetle and aphids. Am. J. Potato Res. 2013, 90, 83–92. [Google Scholar] [CrossRef]
- Lopez, E.R.; Ferro, D.N.; Van Driesche, R.G. Direct measurement of host and parasitoid recruitment for assessment of total losses due to parasitism in the Colorado potato beetle Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae) and Myiopharus doryphorae (Riley) (Diptera: Tach). Biol. Control 1993, 3, 85–92. [Google Scholar] [CrossRef]
- Lopez, E.R.; Roth, L.C.; Ferro, D.N.; Hosmer, D.; Mafra-Neto, A. Behavioral ecology of Myiopharus doryphorae (Riley) and M. aberrans (Townsend), tachinid parasitoids of the Colorado potato beetle. J. Insect Behav. 1997, 10, 49–78. [Google Scholar] [CrossRef]
- Hough-Goldstein, J.; McPherson, D. Comparison of Perillus bioculatus and Podisus maculiventris (Hemiptera: Pentatomidae) as potential control agents of the Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1996, 89, 1116–1123. [Google Scholar] [CrossRef]
- Chang, G.C.; Snyder, W.E. The relationship between predator density, community composition, and field predation of Colorado potato beetle eggs. Biol. Control 2004, 31, 453–461. [Google Scholar] [CrossRef]
- Koss, A.M.; Chang, G.C.; Snyder, W.E. Predation of green peach aphids by generalist predators in the presence of alternative, Colorado potato beetle egg prey. Biol. Control 2004, 31, 237–244. [Google Scholar] [CrossRef]
- Koss, A.M.; Jensen, A.S.; Schreiber, A.; Pike, K.S. Comparison of predator and pest communities in Washington potato fields treated with broad-spectrum, selective, or organic insecticides. Environ. Entomol. 2005, 34, 87–95. [Google Scholar] [CrossRef]
- Ertürk, O.; Yaman, M.; Aslam, I. Effects of four Bacillus spp. of soil origin on the Colorado potato beetle Leptinotarsa decemlineata (Say). Entomol. Res. 2008, 38, 135–138. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Carrière, Y.; Tabashnik, B.E. Fitness costs of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 2009, 54, 147–163. [Google Scholar] [CrossRef]
- Walker, K.; Mendelsohn, M.; Matten, S.; Alphin, M.; Alven, D. The role of microbial Bt products in US crop protection. J. New. Seeds 2003, 5, 31–51. [Google Scholar] [CrossRef]
- Ghassemi-Kahrizeh, A.; Aramideh, S. Sub-lethal effects of Bacillus thuringiensis Berliner on larvae of Colorado potato beetle, Leptinotarsa decemlineata (say) (Coleoptera: Chrysomelidae). Arch. Phytopathol. Pflanzenschutz 2015, 48, 259–267. [Google Scholar] [CrossRef]
- Cingel, A.; Savić, J.; Lazarević, J.; Ćosić, T.; Raspor, M.; Smigocki, A.; Ninković, S. Extraordinary adaptive plasticity of Colorado potato beetle: “Ten-Striped Spearman” in the era of biotechnological warfare. Int. J. Mol. Sci. 2016, 17, 1538. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; He, W.; Xu, S.; Xu, L.; Zhang, J. RNA interference in Colorado potato beetle (Leptinotarsa decemlineata): A potential strategy for pest control. J. Integr. Agric. 2020, 19, 428–437. [Google Scholar] [CrossRef]
- Gui, S.; Taning, C.N.T.; Wei, D.; Smagghe, G. First report on CRISPR/Cas9-targeted mutagenesis in the Colorado potato beetle, Leptinotarsa decemlineata. J. Insect Physiol. 2020, 121, 104013. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, Y. Level and genetic variability of resistance to the Colorado potato beetle (Leptinotarsa decemlineata (Say)) in wild Solanum species. Am. J. Potato Res. 2007, 84, 143–148. [Google Scholar] [CrossRef]
- Mărgineanu, A.-M.; Erdelyi-Molnar, I.; Rakosy-Tican, E. Trichomes types analysis and their density in parental species Solanum tuberosum and S. chacoense and their derived somatic hybrids. An. Ştiinţifice Univ. Al. I. Cuza Iaşi s. II a. Biol. Veg. 2014, 60, 33–42. [Google Scholar]
- Cruceriu, D.; Erdelyi-Molnar, I.; Diaconeasa, Z.; Măgineanu, A.-M.; Aurori, A.; Rakosy-Tican, E. Comparative characterization of somatic hybrids of Solanum bulbocastanum + S. tuberosum cv. ‘Rasant’ with their parents in relation to biochemical responses to wound stress and trichome composition. Studia UBB Chem. 2020, 65, 133–148. [Google Scholar] [CrossRef]
- Thieme, R.; Rakosy-Tican, E. Somatic cell genetics and its application in potato breeding. In The Potato Genome; Chakrabarti, S.K., Xie, C., Tiwari, J.K., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 217–269. [Google Scholar]
- Thieme, R.; Rakosy-Tican, E.; Gavrilenko, T.; Antonova, O.; Schubert, J.; Nachtigall, M.; Heimbach, U.; Thieme, T. Novel somatic hybrids (Solanum tuberosum L. + Solanum tarnii) and their fertile BC1 progenies express extreme resistance to potato virus Y and late blight. Theor Appl. Genet. 2008, 116, 691–700. [Google Scholar] [CrossRef]
- Thieme, R.; Rakosy-Tican, E.; Nachtigall, M.; Schubert, J.; Hammann, T.; Antonova, O.; Gavrilenko, T.; Heimbach, U.; Thieme, T. Characterization of multiple resistance traits of somatic hybrids between Solanum cardiophyllum Lindl. and two commercial potato cultivars. Plant Cell Rep. 2010, 29, 1187–1201. [Google Scholar] [CrossRef] [PubMed]
- Molnár, I.; Besenyei, E.; Thieme, R.; Thieme, T.; Aurori, A.; Baricz, A.; Banciu, H.L.; Rakosy-Tican, E. Mismatch repair deficiency increases the transfer of antibiosis and antixenosis properties against Colorado potato beetle in somatic hybrids of Solanum tuberosum + S. chacoense. Pest Manag. Sci. 2016, 73, 1428–1437. [Google Scholar] [CrossRef] [PubMed]
- Rakosy-Tican, E.; Besenyei, E.; Molnár, I.; Thieme, R.; Hartung, F.; Sprink, T.; Antonova, O.; Famelaer, I.; Angenon, G.; Aurori, A. New phenotypes of potato co-induced by mismatch repair deficiency and somatic hybridization. Front. Plant Sci. 2019, 10, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molnar, I.; Rakosy-Tican, E. Difficulties in Potato Pest Control: The Case of Pyrethroids on Colorado Potato Beetle. Agronomy 2021, 11, 1920. https://doi.org/10.3390/agronomy11101920
Molnar I, Rakosy-Tican E. Difficulties in Potato Pest Control: The Case of Pyrethroids on Colorado Potato Beetle. Agronomy. 2021; 11(10):1920. https://doi.org/10.3390/agronomy11101920
Chicago/Turabian StyleMolnar, Imola, and Elena Rakosy-Tican. 2021. "Difficulties in Potato Pest Control: The Case of Pyrethroids on Colorado Potato Beetle" Agronomy 11, no. 10: 1920. https://doi.org/10.3390/agronomy11101920
APA StyleMolnar, I., & Rakosy-Tican, E. (2021). Difficulties in Potato Pest Control: The Case of Pyrethroids on Colorado Potato Beetle. Agronomy, 11(10), 1920. https://doi.org/10.3390/agronomy11101920