
Effect of Water Stress on Grain Yield and Physiological Characters of Quinoa Genotypes

 ,
,  , ,
, ,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Hydroponic Culture
2.3. Morphological Traits
- W1 = root/shoot length at first harvest.
- W2 = root/shoot length at second harvest.
- T2 − T1 = time interval between two harvests (10 days).
2.4. Leaf Biochemical Analysis
2.5. Membrane Stability Index (MSI)
2.6. Relative Water Content (RWC)
2.7. Determination of Leaf K+ Concentrations
2.8. Pot Study
2.9. Biomass and Grain Yield
2.10. Drought Tolerance Index (DTI) Based on Seed Grain Yield
- Y1 = mean of a genotype seed yield at control.
- AY1 = average mean of all genotypes seed yield at control.
- Y2 = mean of a genotype seed yield at water stress.
- AY1 = average mean of all genotypes seed yield at water stress.
2.11. Statistical Analysis
3. Results
3.1. Quinoa Genotypes Performance in Hydroponic Culture
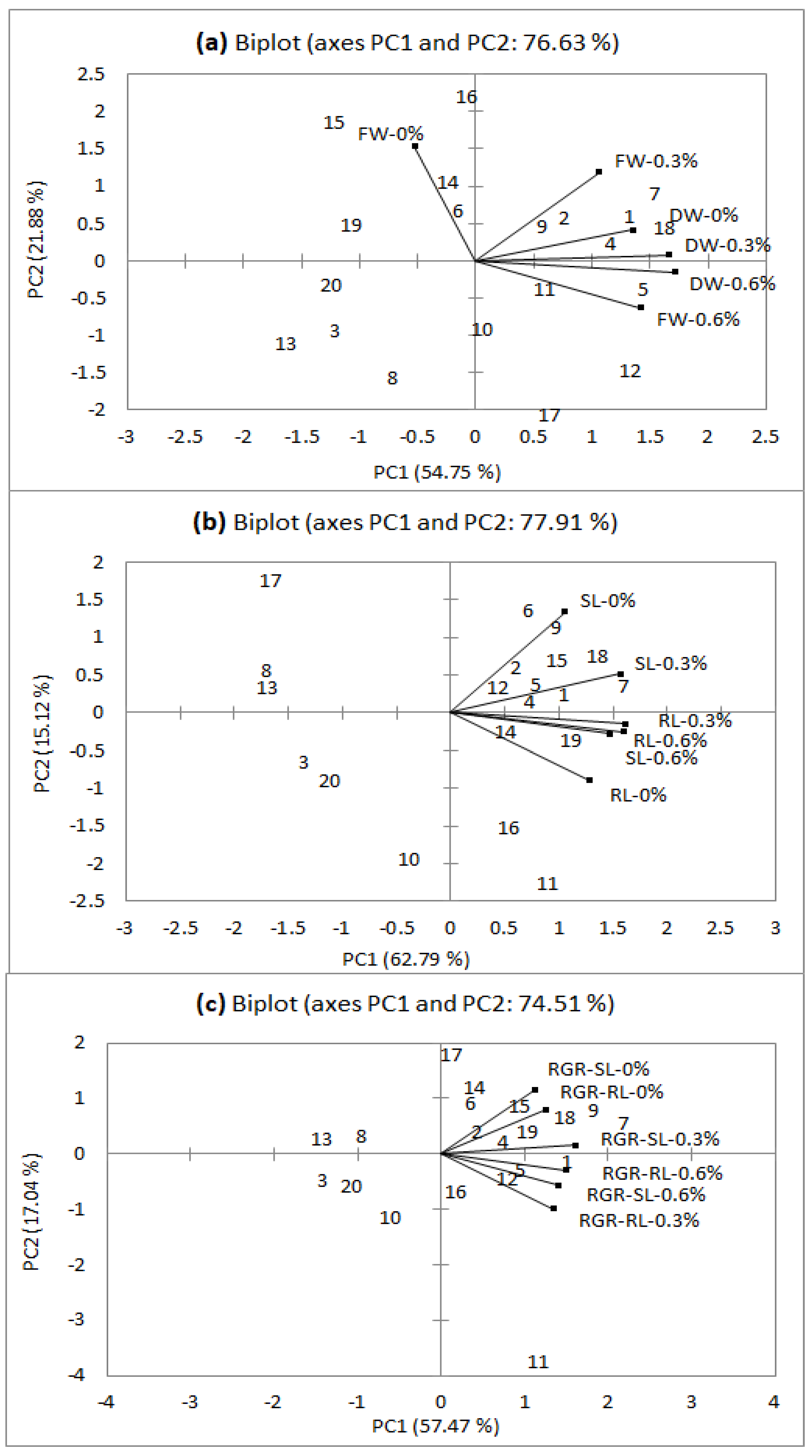
3.1.1. Biplot for Seedling Fresh Weight and Dry Weight
3.1.2. Biplot for Seedling Growth (Root and Shoot Length) and Relative Growth Rates
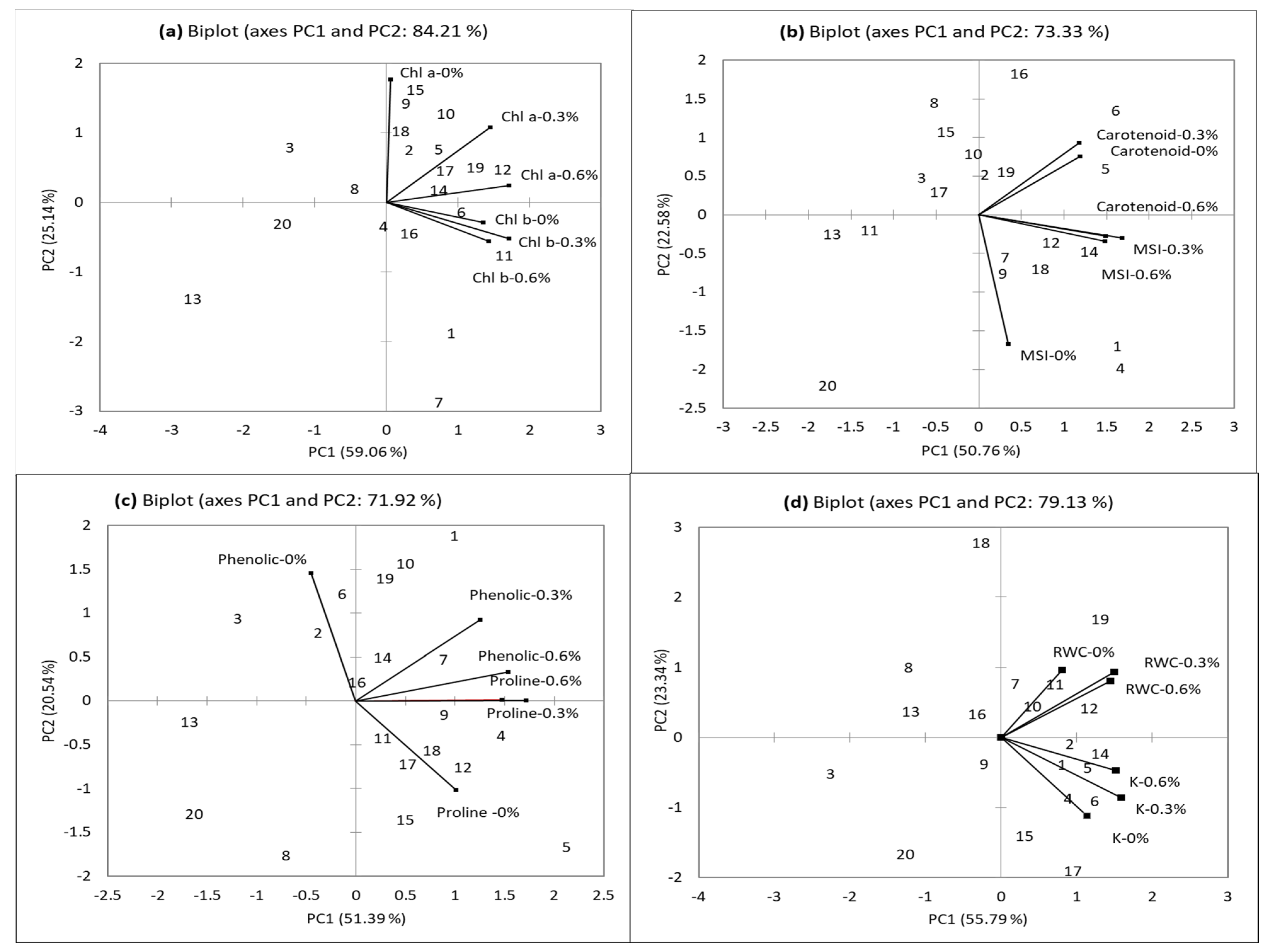
3.1.3. Biplot for Physiological Traits
3.2. Result of Pot Study
3.2.1. Yield Related Attributes
3.2.2. Drought Tolerance Index (DTI) Based on Seed Grain Yield
3.2.3. Correlation among the Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akram, M.Z.; Basra, S.M.A.; Hafeez, M.B.; Khan, S.; Nazeer, S.; Iqbal, S.; Saddiq, M.S.; Zahra, N. Adaptability and yield potential of new quinoa lines under agroecological conditions of Faisalabad-Pakistan. Asian J. Agric. Biol. 2021, 2, 1–8. [Google Scholar]
- Maradini-Filho, A. Quinoa: Nutritional aspects. J. Nutraceuticals Food Sci. 2017, 2, 1–5. [Google Scholar]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazile, D.; Bertero, H.D.; Nieto, C. State of the Art Report on Quinoa around the World in 2013; FAO: Roman, Italy, 2015; Available online: https://agritrop.cirad.fr/575489/1/document_575489.pdf (accessed on 1 August 2021).
- Azurita Silva, A.; Jacobsen, S.-E.; Razzaghi, F.; Álvarez Flores, R.; Ruiz, K.B.; Morales, A.; Silva Ascencio, H. Quinoa drought responses and adaptation. In State of the Art Report on Quinoa around the World; FAO: Roma, Italy, 2015; pp. 157–171. [Google Scholar]
- Jha, P.K.; Ines, A.V.; Singh, M.P. A multiple and ensembling approach for calibration and evaluation of genetic coefficients of CERES-maize to simulate maize phenology and yield in Michigan. Environ. Model. Softw. 2021, 135, 104901. [Google Scholar] [CrossRef]
- Jacobsen, S.-E.; Liu, F.; Jensen, C.R. Does root-sourced ABA play a role for regulation of stomata under drought in quinoa (Chenopodium quinoa Willd.). Sci. Hortic. 2009, 122, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Skirycz, A.; Inzé, D. More from less: Plant growth under limited water. Curr. Opin. Biotechnol. 2010, 21, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Roumet, C.; Urcelay, C.; Díaz, S. Suites of root traits differ between annual and perennial species growing in the field. New Phytol. 2006, 170, 357–368. [Google Scholar] [CrossRef]
- Nicotra, A.; Babicka, N.; Westoby, M. Seedling root anatomy and morphology: An examination of ecological differentiation with rainfall using phylogenetically independent contrasts. Oecologia 2002, 130, 136–145. [Google Scholar] [CrossRef]
- Garrido, M.; Silva, P.; Silva, H.; Muñoz, R.; Baginsky, C.; Acevedo, E. Evaluación del rendimiento de nueve genotipos de quinua (Chenopodium quinoa Willd.) j bajo diferentes disponibilidades hídricas en ambiente mediterráneo. Idesia 2013, 31, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Geerts, S.; Raes, D.; Garcia, M.; Mendoza, J.; Huanca, R. Crop water use indicators to quantify the flexible phenology of quinoa (Chenopodium quinoa Willd.) in response to drought stress. Field Crop. Res. 2008, 108, 150–156. [Google Scholar] [CrossRef]
- Zhao, B.; Ma, B.-L.; Hu, Y.; Liu, J. Source–sink adjustment: A mechanistic understanding of the timing and severity of drought stress on photosynthesis and grain yields of two contrasting oat (Avena sativa L.) genotypes. J. Plant Growth Regul. 2021, 40, 263–276. [Google Scholar] [CrossRef]
- Jha, P.K.; Kumar, S.N.; Ines, A.V. Responses of soybean to water stress and supplemental irrigation in upper Indo-Gangetic plain: Field experiment and modeling approach. Field Crop. Res. 2018, 219, 76–86. [Google Scholar] [CrossRef]
- Alghabari, F.; Ihsan, M.Z. Effects of drought stress on growth, grain filling duration, yield and quality attributes of barley (Hordeum vulgare L.). Bangladesh J. Bot. 2018, 47, 421–428. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Zohaib, A.; Tanveer, M.; Naeem, M.; Ali, I.; Tabassum, T.; Nazir, U. Growth and development responses of crop plants under drought stress: A review. Zemdirbyste 2017, 104, 267–276. [Google Scholar] [CrossRef]
- Jensen, C.; Jacobsen, S.-E.; Andersen, M.; Nunez, N.; Andersen, S.; Rasmussen, L.; Mogensen, V. Leaf gas exchange and water relation characteristics of field quinoa (Chenopodium quinoa Willd.) during soil drying. Eur. J. Agron. 2000, 13, 11–25. [Google Scholar] [CrossRef]
- Jellen, E.N.; Maughan, P.J.; Bertero, D.; Munir, H. Prospects for quinoa (Chenopodium Quinoa Willd.) improvement through biotechnology. In Biotechnology of Neglected and Underutilized Crops; Springer: Berlin, Germany, 2013; pp. 173–201. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. In California Agricultural Experiment Station; Circular 347; University of California: Berkeley, CA, USA, 1950. [Google Scholar]
- Gardner, F.; Pearce, R.; Mitchell, R. Physiology of Crop Plants, 2nd ed.; Iowa State University Press: Ames, IA, USA, 1985. [Google Scholar]
- Nagata, M.; Yamashita, I. Simple method for simultaneous determination of chlorophyll and carotenoids in tomato fruit. Nippon Shokuhin Kogyo Gakkaishi 1992, 39, 925–928. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Premachandra, G.S.; Saneoka, H.; Ogata, S. Cell membrane stability, an indicator of drought tolerance, as affected by applied nitrogen in soyabean. J. Agric. Sci. 1990, 115, 63–66. [Google Scholar] [CrossRef]
- Barrs, H.; Weatherley, P. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Munns, R.; James, R.A. Screening methods for salinity tolerance: A case study with tetraploid wheat. Plant Soil 2003, 253, 201–218. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Langridge, P.; Tester, M. Salinity tolerance and sodium exclusion in genus Triticum. Breed. Sci. 2009, 59, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Faran, M.; Farooq, M.; Rehman, A.; Nawaz, A.; Saleem, M.K.; Ali, N.; Siddique, K.H. High intrinsic seed Zn concentration improves abiotic stress tolerance in wheat. Plant Soil 2019, 437, 195–213. [Google Scholar] [CrossRef]
- Yan, W. GGEbiplot—A windows application for graphical analysis of multi environment trial data and other types of two-way data. Agron. J. 2001, 93, 1111–1118. [Google Scholar] [CrossRef] [Green Version]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Mukami, A.; Ngetich, A.; Mweu, C.; Oduor, R.O.; Muthangya, M.; Mbinda, W.M. Differential characterization of physiological and biochemical responses during drought stress in finger millet varieties. Physiol. Mol. Biol. Plants 2019, 25, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Al-Naggar, A.; Abd El-Salam, R.; Badran, A.; El-Moghazi, M.M. Drought tolerance of five Quinoa (Chenopodium quinoa Willd.) genotypes and its association with other traits under moderate and severe drought stress. Asian J. Adv. Agric. Res. 2017, 3, 1–13. [Google Scholar] [CrossRef]
- Adolf, V.I.; Jacobsen, S.-E.; Shabala, S. Salt tolerance mechanisms in quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Shabala, L.; Mackay, A.; Tian, Y.; Jacobsen, S.E.; Zhou, D.; Shabala, S. Oxidative stress protection and stomatal patterning as components of salinity tolerance mechanism in quinoa (Chenopodium quinoa). Physiol. Plant. 2012, 146, 26–38. [Google Scholar] [CrossRef]
- Claeys, H.; Inzé, D. The agony of choice: How plants balance growth and survival under water-limiting conditions. Plant Physiol. 2013, 162, 1768–1779. [Google Scholar] [CrossRef] [PubMed]
- Kranner, I.; Minibayeva, F.V.; Beckett, R.P.; Seal, C.E. What is stress? Concepts, definitions and applications in seed science. New Phytol. 2010, 188, 655–673. [Google Scholar] [CrossRef]
- Khan, S.; Basit, A.; Hafeez, M.B.; Irshad, S.; Bashir, S.; Bashir, S.; Maqbool, M.M.; Saddiq, M.S.; Hasnain, Z.; Aljuaid, B.S. Moringa leaf extract improves biochemical attributes, yield and grain quality of rice (Oryza sativa L.) under drought stress. PLoS ONE 2021, 16, e0254452. [Google Scholar] [CrossRef]
- Li, Z.; Su, X.; Chen, Y.; Fan, X.; He, L.; Guo, J.; Wang, Y.; Yang, Q. Melatonin improves drought resistance in maize seedlings by enhancing the antioxidant system and regulating abscisic acid metabolism to maintain stomatal opening under PEG-induced drought. J. Plant Biol. 2021, 64, 299–312. [Google Scholar] [CrossRef]
- Fghire, R.; Anaya, F.; Ali, O.I.; Benlhabib, O.; Ragab, R.; Wahbi, S. Physiological and photosynthetic response of quinoa to drought stress. Chil. J. Agric. Res. 2015, 75, 174–183. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Liu, F.; Bendevis, M.; Shabala, S.; Jacobsen, S.E. Sensitivity of two quinoa (Chenopodium quinoa Willd.) varieties to progressive drought stress. J. Agron. Crop Sci. 2014, 200, 12–23. [Google Scholar] [CrossRef]
- González, J.; Bruno, M.; Valoy, M.; Prado, F. Genotypic variation of gas exchange parameters and leaf stable carbon and nitrogen isotopes in ten quinoa cultivars grown under drought. J. Agron. Crop Sci. 2011, 197, 81–93. [Google Scholar] [CrossRef]
- Spehar, C.R.; Santos, R.L.d.B. Agronomic performance of quinoa selected in the Brazilian Savannah. Pesqui. Agropecu. Bras. 2005, 40, 609–612. [Google Scholar] [CrossRef]
- Mignone, C.; Bertero, H. Identificación del período crítico de determinación del rendimiento en quínoas de nivel del mar. In Proceedings of the Congreso Internacional de la Quinua, Iquique, Chile, 23–27 October 2007; pp. 23–26. [Google Scholar]
- Bhargava, A.; Shukla, S.; Rajan, S.; Ohri, D. Genetic diversity for morphological and quality traits in quinoa (Chenopodium quinoa Willd.) germplasm. Genet. Resour. Crop Evol. 2007, 54, 167–173. [Google Scholar] [CrossRef]
- Yang, A.; Akhtar, S.; Amjad, M.; Iqbal, S.; Jacobsen, S.E. Growth and physiological responses of quinoa to drought and temperature stress. J. Agron. Crop Sci. 2016, 202, 445–453. [Google Scholar] [CrossRef]
- Raza, A.; Charagh, S.; Sadaqat, N.; Jin, W. Arabidopsis thaliana: Model plant for the study of abiotic stress responses. In The Plant Family Brassicaceae; Springer: Singapore, 2020; pp. 129–180. [Google Scholar]



{kind=link}
{kind=link}
| Code No. | Accession No. | Plant Name | Origin |
|---|---|---|---|
| 1 | Ames 13724 | 18GR | United States, New Mexico |
| 2 | Ames 13726 | 49ALC | United States, New Mexico |
| 3 | Ames 13729 | 23P | United States, New Mexico |
| 4 | Ames 13731 | 42TES | United States, New Mexico |
| 5 | Ames 13735 | 17GR | United States, New Mexico |
| 6 | Ames 13737 | V7 | United States, New Mexico |
| 7 | Ames 13742 | 6P | United States, New Mexico |
| 8 | Ames 13744 | 409 | United States, New Mexico |
| 9 | Ames 13747 | APELAWA | Bolivia |
| 10 | Ames 13750 | A5P | United States, New Mexico |
| 11 | PI 433232 | - | Chile |
| 12 | PI 478414 | R-70 | Bolivia, La Paz |
| 13 | PI 478415 | R-71 | Bolivia, La Paz |
| 14 | PI 478418 | R-132 | Bolivia, Potosi |
| 15 | PI 510536 | Quiona Negra (Spain) | Peru |
| 16 | PI 510549 | Yulaj K’oyto (Quechua) | Peru |
| 17 | PI 614880 | QQ065 | Chile, Los Lagos |
| 18 | PI 614887 | QQ63 | Chile, Bio-Bio |
| 19 | PI 614888 | QQ61 | Chile, Bio-Bio |
| 20 | PI 665283 | Col.#6197 | United States, Colorado |
| Characteristics | Units | Value |
|---|---|---|
| Sand | % | 51.1 |
| Silt | % | 26.1 |
| Clay | % | 18.2 |
| Textural class | - | Sandy clay loam |
| Saturation percentage | % | 30.0 |
| pH | - | 7.6 |
| EC | dS m−1 | 1.31 |
| Available phosphorous (olson) | mg kg−1 | 3.31 |
| Extractable potassium (NH4OAC) | mg kg−1 | 81.0 |
| Organic matter | % | 0.72 |
| Total nitrogen | % | 0.07 |
| Hydroponic Study | |||
|---|---|---|---|
| Source | G | WSL | G * WSL |
| Degree of freedom | 19 | 2 | 38 |
| Seedling fresh weight (g) | 2.100 n.s. | 260.675 ** | 1.956 n.s. |
| Seedling dry weight (g) | 1.7969 ** | 43.9225 ** | 0.3442 ** |
| Root length (cm) | 83.17 ** | 1655.57 ** | 12.61 ** |
| Shoot length (cm) | 36.343 ** | 968.110 ** | 9.193 n.s. |
| Relative growth rate of root length | 0.00121 ** | 0.00093 ** | 0.00038 |
| Relative growth rate of shoot length | 0.00020 ** | 0.00463 ** | 0.00006 * |
| Chlorophyll a | 0.00099 * | 0.04190 ** | 0.00029 n.s. |
| Chlorophyll b | 0.01063 ** | 0.87907 ** | 0.00187 n.s. |
| Carotenoid | 0.7644 ** | 45.667 ** | 0.275 ** |
| Leaf phenolic | 0.00298 ** | 0.16672 ** | 0.00161 ** |
| Leaf proline | 0.1579 ** | 15.2162 ** | 0.0455 ** |
| Membrane stability index | 0.00019 ** | 0.03517 ** | 0.00009 ** |
| Water use efficiency | 138.6 ** | 25,152.4 ** | 35.6 ** |
| K+ in leaf | 267.63 ** | 4667.54 ** | 36.30 ** |
| Pot Study | |||
| Source | G | WSL | GN * WSL |
| Degree of freedom | 19 | 2 | 38 |
| Plant height (cm) | 5.659 ** | 433.068 ** | 3.448 ** |
| Panicle length (cm) | 1.927 ** | 102.050 ** | 0.310 n.s. |
| 1000 grain weight (g) | 0.3159 ** | 52.7254 ** | 0.1827 * |
| Grain yield plant−1 (g) | 0.4065 ** | 88.6783 ** | 0.2133 ** |
| Biological yield plant−1 (g) | 8.44 ** | 1093.09 ** | 4.51 n.s. |
| Drought tolerance index (DTI) | 0.06933 *** | 0.00024 n.s. | 0.00438 n.s. |
| Genotype | Plant Height (cm) | 1000 Grain Weight (g) | Grain Yield Plant−1 (g) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 0% | 50% | 75% | 0% | 50% | 75% | 0% | 50% | 75% | |
| Ames 13724 (1) | 14.33 b–f | 13.33 e–j | 9.00 q–t | 2.98 b–i | 2.73 e–m | 1.43 r–u | 4.39 d–k | 4.55 b–h | 2.62 qr |
| Ames 13726 (2) | 15.33 a–e | 13.33 e–j | 9.33 o–t | 2.91 c–j | 2.55 g–n | 1.41 r–u | 4.25 f–l | 4.07 h–m | 2.43 rs |
| Ames 13729 (3) | 14.00 c–g | 11.00 k–q | 8.00 r–t | 2.85 d–l | 2.08 n–q | 1.20 r–u | 4.37 e–k | 3.60 m–p | 2.22 r–t |
| Ames 13731 (4) | 14.00 c–g | 13.50 d–i | 10.17 l–r | 3.29 b–d | 2.48 i–n | 1.46 r–u | 4.81 b–e | 4.00 i–m | 2.48 rs |
| Ames 13735 (5) | 15.33 a–e | 13.17 e–k | 10.67 l–q | 3.11 b–e | 2.68 e–m | 1.52 r–u | 4.63 b–f | 4.20 f–l | 2.54 rs |
| Ames 13737 (6) | 15.67 a–d | 12.33 f–l | 10.33 l–q | 2.91 c–j | 2.75 e–l | 1.51 r–u | 4.43 c–i | 4.27 f–k | 2.53 rs |
| Ames 13742 (7) | 14.33 b–f | 11.17 j–q | 9.17 p–t | 3.10 b–f | 2.71 e–m | 1.43 r–u | 4.62 b–f | 4.23 f–l | 2.45 rs |
| Ames 13744 (8) | 14.00 c–g | 10.33 l–q | 7.83 st | 2.87 d–l | 1.58 q–t | 1.08 tu | 4.39 d–k | 3.10 pq | 1.85 t |
| Ames 13747 (9) | 15.33 a–e | 10.33 l–q | 9.17 p–t | 3.07 b–g | 2.36 l–n | 1.15 s–u | 4.59 b–g | 3.88 k–m | 2.10 r–t |
| Ames 13750 (10) | 15.33 a–e | 11.50 h–o | 9.67 n–t | 4.01 a | 2.15 n–p | 1.15 s–u | 5.53 a | 3.67 m–o | 2.10 r–t |
| PI 433232 (11) | 13.67 c–h | 11.67 h–n | 10.17 l–r | 3.42 bc | 2.21 m–o | 1.05 u | 4.94 bc | 3.73 l–n | 2.07 st |
| PI 478414 (12) | 14.67 b–e | 14.00 c–g | 10.50 l–q | 3.01 b–h | 2.35 l–n | 1.59 q–t | 4.53 b–h | 3.87 k–m | 2.61 qr |
| PI 478415 (13) | 17.00 a | 9.50 n–t | 7.67 st | 3.10 b–f | 1.72 o–r | 1.20 | 4.62 b–g | 3.24 n–p | 2.06 st |
| PI 478418 (14) | 14.00 c–g | 11.00 k–q | 9.83 m–s | 3.08 b–f | 2.21 m–o | 1.25 r–u | 4.60 b–g | 3.73 l–n | 2.29 r–t |
| PI 510536 (15) | 15.67 a–d | 9.67 n–t | 10.33 l–q | 3.38 b–d | 2.38 k–n | 1.24 r–u | 4.90 b–d | 3.90 j–m | 2.26 r–t |
| PI 510549 (16) | 15.83 a–c | 12.00 g–m | 10.50 l–q | 3.48 b | 2.58 f–n | 1.29 r–u | 5.00 b | 4.10 g–m | 2.31 r–t |
| PI 614880 (17) | 16.33 ab | 11.33 i–p | 11.00 k–q | 3.10 b–f | 2.40 j–n | 1.10 tu | 4.62 b–g | 3.92 i–m | 2.11 r–t |
| PI 614887 (18) | 15.33 a–e | 12.33 f–l | 12.00 g–m | 3.15 b–e | 2.52 h–n | 1.30 r–u | 4.67 b–f | 4.04 h–m | 2.26 r–t |
| PI 614888 (19) | 15.33 a–e | 11.67 h–n | 11.50 h–o | 3.20 b–e | 2.15 n–p | 1.11 tu | 4.72 b–f | 3.67 m–o | 2.13 r–t |
| PI 665283 (20) | 15.00 a–e | 9.83 m–s | 7.50 t | 2.90 c–k | 1.65 p–s | 1.01 u | 4.42 d–j | 3.17 o–p | 1.83 t |
| CVCs | G * WSL = 2.1889 | G * WSL = 0.5234 | G * WSL = 0.5233 | ||||||
| Genotype | Panicle Length (cm) | Biological Yield Plant−1 (g) | DTI | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0% | 50% | 75% | Mean | 0% | 50% | 75% | Mean | 50% | 75% | Mean | |
| Ames 13724 (1) | 6.17 | 5.17 | 4.10 | 5.14 a–c | 19.83 | 15.6 | 11.00 | 15.5 a | 1.12 (T) | 1.13 (T) | (T) 1.10 a–c |
| Ames 13726 (2) | 6.00 | 4.83 | 3.73 | 4.85 a–e | 17.33 | 14.67 | 10.57 | 14.2 a–d | 0.97 (S) | 0.98 (S) | (S) 0.97 b–e |
| Ames 13729 (3) | 6.33 | 4.00 | 2.83 | 4.38 ef | 17.00 | 14.50 | 8.90 | 13.5 d–f | 0.88 (S) | 0.92 (S) | (S) 0.90 d–f |
| Ames 13731 (4) | 6.17 | 5.50 | 4.00 | 5.22 ab | 16.83 | 14.33 | 10.40 | 13.9 a–e | 1.07 (T) | 1.14 (T) | (T) 1.10 a–c |
| Ames 13735 (5) | 6.33 | 5.00 | 4.00 | 5.11 a–d | 19.33 | 16.00 | 10.33 | 15.2 ab | 1.09 (T) | 1.12 (T) | (T) 1.10 a–c |
| Ames 13737 (6) | 6.33 | 5.83 | 4.17 | 5.44 a | 19.00 | 15.83 | 10.67 | 15.2 a | 1.06 (T) | 1.07 (T) | (T) 1.06 a–c |
| Ames 13742 (7) | 5.33 | 4.00 | 3.83 | 4.38 ef | 17.67 | 15.67 | 10.00 | 14.4 a–e | 1.09 (T) | 1.08 (T) | (T) 1.09 a–c |
| Ames 13744 (8) | 5.67 | 3.83 | 2.50 | 4.00 fg | 18.33 | 12.67 | 6.33 | 12.4 f | 0.76 (S) | 0.77 (S) | (S) 0.76 f |
| Ames 13747 (9) | 6.00 | 4.50 | 3.17 | 4.55 b–f | 19.57 | 14.33 | 10.17 | 14.7 a | 1.00 (T) | 0.92 (S) | (S) 0.95 c–e |
| Ames 13750 (10) | 5.50 | 4.17 | 3.17 | 4.27 e–g | 19.67 | 14.33 | 8.73 | 14.2 a–e | 1.13 (T) | 1.11 (T) | (T) 1.12 ab |
| PI 433232 (11) | 6.33 | 4.50 | 3.17 | 4.66 b–f | 16.00 | 16.07 | 8.77 | 13.6 c–f | 1.03 (T) | 0.97 (S) | (T) 1.00 a–e |
| PI 478414 (12) | 6.67 | 5.00 | 4.57 | 5.41 a | 17.33 | 14.67 | 10.33 | 14.1 a–e | 0.98 (S) | 1.13 (T) | (T) 1.05 a–c |
| PI 478415 (13) | 6.00 | 3.83 | 3.00 | 4.27 e–g | 18.27 | 13.00 | 7.83 | 13.0 d–f | 0.84 (S) | 0.91 (S) | (S) 0.86 ef |
| PI 478418 (14) | 6.33 | 4.83 | 3.33 | 4.83 a–e | 17.33 | 15.10 | 9.87 | 14.1 a–d | 0.96 (S) | 1.00 (T) | (S) 0.97 b–e |
| PI 510536 (15) | 6.00 | 4.00 | 3.50 | 4.50 c–f | 18.17 | 15.33 | 10.00 | 14.5 a–e | 1.07 (T) | 1.05 (T) | (T) 1.06 a–c |
| PI 510549 (16) | 6.17 | 4.93 | 3.67 | 4.92 a–e | 18.33 | 15.67 | 10.37 | 14.8 a–c | 1.15 (T) | 1.10 (T) | (T) 1.12 a |
| PI 614880 (17) | 6.33 | 4.83 | 3.33 | 4.83 a–e | 16.33 | 15.17 | 10.40 | 14.0 a–e | 1.01 (T) | 0.93 (S) | (S) 0.96 c–e |
| PI 614887 (18) | 6.00 | 4.33 | 3.17 | 4.5 c–f | 18.33 | 13.83 | 9.63 | 13.9 a–e | 1.05 (T) | 1.00 (T) | (T) 1.02 a–d |
| PI 614888 (19) | 5.83 | 4.33 | 3.17 | 4.44 d–f | 16.33 | 11.67 | 9.67 | 12.6 b–f | 0.97 (T) | 0.96 (S) | (S) 0.96 c–e |
| PI 665283 (20) | 5.33 | 3.17 | 2.50 | 3.67 g | 18.67 | 14.17 | 7.07 | 13.3 ef | 0.78 (T) | 0.77 (S) | (S) 0.77 f |
| Mean | 6.04 a | 4.53 b | 3.44 c | 18.0 a | 12.7 b | 9.55 c | 1.00 | 0.99 | |||
| CVCs | G = 0.6908 WSL = 0.2676 | G = 1.8344 WSL = 0.7105 | G = 0.1471 T = Tolerant, S = Sensitive | ||||||||
| Traits | Yield Traits | Growth Traits | Physiological Traits | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GY | BY | PL | 1000 Wt | RL | SL | SFW | SDW | K+ | RWC | MSI | Chl a | Chl b | Chl c | Phenol | Proline | |
| BY | 0.80 *** | |||||||||||||||
| PL | 0.80 *** | 0.77 ** | ||||||||||||||
| 1000 Wt | 0.98 | 0.80 | 0.80 | |||||||||||||
| RL | 0.72 ** | 0.77 ** | 0.67 ** | 0.71 ** | ||||||||||||
| SL | 0.65 ** | 0.68 ** | 0.63 ** | 0.65 ** | 0.66 ** | |||||||||||
| SFW | 0.68 ** | 0.66 ** | 0.60 ** | 0.60 ** | 0.68 ** | 0.65 ** | ||||||||||
| SDW | 0.66 ** | 0.69 ** | 0.67 ** | 0.63 ** | 0.64 ** | 0.67 ** | 0.64 ** | |||||||||
| K+ | 0.74 ** | 0.74 ** | 0.73 ** | 0.73 ** | 0.68 ** | 0.60 ** | 0.59 ** | 0.63 ** | ||||||||
| RWC | 0.90 *** | 0.73 ** | 0.78 ** | 0.87 *** | 0.71 ** | 0.65 ** | 0.71 ** | 0.66 ** | 0.76 ** | |||||||
| MSI | 0.83 *** | 0.81 *** | 0.80 *** | 0.83 *** | 0.71 ** | 0.70 ** | 0.76 ** | 0.74 ** | 0.73 ** | 0.86 *** | ||||||
| Chl a | 0.64 ** | 0.63 ** | 0.62 ** | 0.63 ** | 0.51 ** | 0.46 * | 0.43 * | 0.57 ** | 0.65 ** | 0.71 ** | 0.63 ** | |||||
| Chl b | 0.82 *** | 0.77 ** | 0.75 ** | 0.81 *** | 0.68 ** | 0.58 ** | 0.61 ** | 0.64 ** | 0.76 ** | 0.86 *** | 0.82 *** | 0.83 *** | ||||
| Chl c | 0.83 *** | 0.73 ** | 0.75 ** | 0.57 ** | 0.62 ** | 0.57 ** | 0.63 ** | 0.66 ** | 0.71 ** | 0.85 *** | 0.77 ** | 0.71 ** | 0.83 *** | |||
| Phenol | 0.64 ** | 0.61 ** | 0.53 ** | 0.63 ** | 0.51 ** | 0.52 ** | 0.56 ** | 0.53 ** | 0.48 * | 0.65 ** | 0.68 ** | 0.51 ** | 0.63 ** | 0.55 ** | ||
| Proline | 0.78 ** | 0.72 ** | 0.66 ** | 0.78 ** | 0.64 ** | 0.63 ** | 0.68 ** | 0.68 ** | 0.57 ** | 0.78 ** | 0.82 *** | 0.52 ** | 0.73 ** | 0.66 ** | 0.82 *** | |
| DTI | 0.99 *** | 0.74 ** | 0.66 ** | 0.98 *** | 0.70 ** | 0.58 ** | 0.60 ** | 0.59 ** | 0.52 ** | 0.58 | 0.21 * | 0.42 * | 0.51 ** | 0.58 ** | 0.69 ** | 0.71 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saddiq, M.S.; Wang, X.; Iqbal, S.; Hafeez, M.B.; Khan, S.; Raza, A.; Iqbal, J.; Maqbool, M.M.; Fiaz, S.; Qazi, M.A.; et al. Effect of Water Stress on Grain Yield and Physiological Characters of Quinoa Genotypes. Agronomy 2021, 11, 1934. https://doi.org/10.3390/agronomy11101934
Saddiq MS, Wang X, Iqbal S, Hafeez MB, Khan S, Raza A, Iqbal J, Maqbool MM, Fiaz S, Qazi MA, et al. Effect of Water Stress on Grain Yield and Physiological Characters of Quinoa Genotypes. Agronomy. 2021; 11(10):1934. https://doi.org/10.3390/agronomy11101934
Chicago/Turabian StyleSaddiq, Muhammad Sohail, Xiukang Wang, Shahid Iqbal, Muhammad Bilal Hafeez, Shahbaz Khan, Ali Raza, Javaid Iqbal, Muhammad Mudassar Maqbool, Sajid Fiaz, Muhammad Akram Qazi, and et al. 2021. "Effect of Water Stress on Grain Yield and Physiological Characters of Quinoa Genotypes" Agronomy 11, no. 10: 1934. https://doi.org/10.3390/agronomy11101934
APA StyleSaddiq, M. S., Wang, X., Iqbal, S., Hafeez, M. B., Khan, S., Raza, A., Iqbal, J., Maqbool, M. M., Fiaz, S., Qazi, M. A., Bakhsh, A., Jahanzaib, & Gulshan, A. B. (2021). Effect of Water Stress on Grain Yield and Physiological Characters of Quinoa Genotypes. Agronomy, 11(10), 1934. https://doi.org/10.3390/agronomy11101934