
Evaluation of Drought Tolerance of Some Wheat (Triticum aestivum L.) Genotypes through Phenology, Growth, and Physiological Indices


 ,
,  , ,
, ,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Location and Duration
2.2. Soil
2.3. Climate
2.4. Experimental Design and Treatments
2.5. Experimentation
2.6. Data Collection
2.6.1. Phenological Indices
2.6.2. Physiological Indices
Relative Water Content (RWC)
Chlorophyll Estimation
Canopy Temperature Depression (CTD)
Estimation of Proline
Measurement of Photo-Assimilates
2.7. Statistical Analysis
3. Results and Discussion
3.1. Phenological Characteristics
3.1.1. Days to Booting (DB)
3.1.2. Days to Heading (DH)
3.1.3. Days to Anthesis (DA)
3.1.4. Days to Physiological Maturity (DPM)
3.2. Physiological Indices
3.2.1. Relative Water Content (RWC)
3.2.2. Chlorophyll Content
3.2.3. Canopy Temperature Depression (CTD)
3.2.4. Proline Content
3.3. Accumulation of Photo-Assimilates
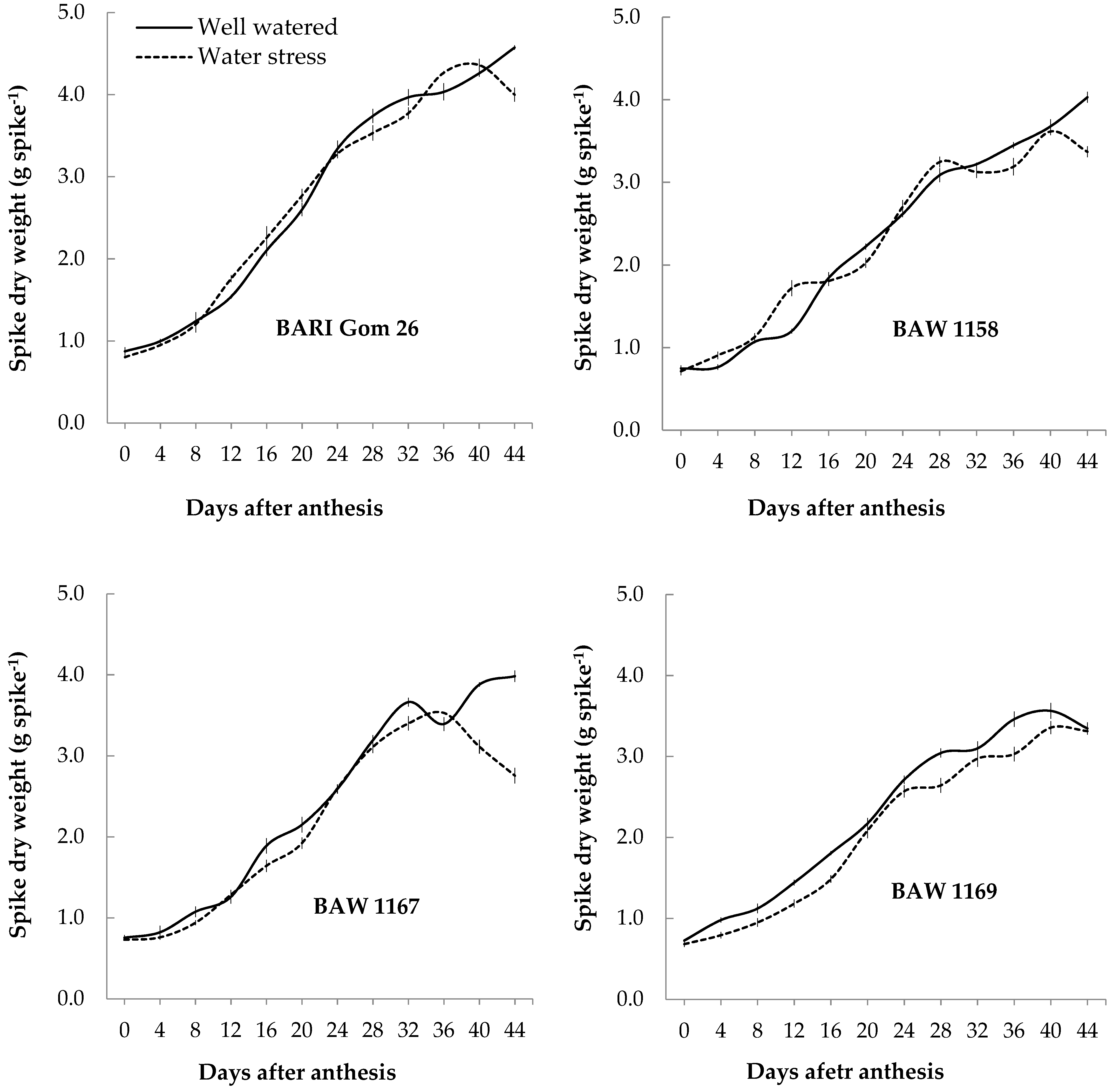
3.3.1. Spike Dry Matter (SDM)
3.3.2. Dry Weight of Grains Spike−1
3.3.3. Grain Yield
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Irshad, M.; Ullah, F.; Fahad, S.; Mehmood, S.; Khan, A.U.; Brtnicky, M.; Kintl, A.; Holatko, J.; Irshad, I.; El-Sharnouby, M.; et al. Evaluation of Jatropha curcas L. leaves mulching on wheat growth and biochemical attributes under water stress. BMC Plant Biol. 2021, 21, 303. [Google Scholar] [CrossRef]
- Asseng, S.; Foster, I.; Turner, N.C. The impact of temperature variability on wheat yields. Glob. Chang. Biol. 2011, 17, 997–1012. [Google Scholar] [CrossRef]
- EL Sabagh, A.; Islam, M.S.; Skalicky, M.; Raza, M.A.; Singh, K.; Anwar Hossain, M.; Hossain, A.; Mahboob, W.; Iqbal, M.A.; Ratnasekaera, D.; et al. Salinity Stress in Wheat (Triticum aestivum L.) in the Changing Climate: Adaptation and Management Strategies. Fron. Agron. 2021, 3, 43. [Google Scholar]
- Karim, M.A.; Hamid, A.; Rahman, S. Grain growth and yield performance of wheat under subtropical conditions: II. Effect of water stress at reproductive stress. Cereal Res. Comm. 2000, 28, 101–107. [Google Scholar] [CrossRef]
- Raza, A.; Mehmood, S.S.; Shah, T.; Zou, X.; Yan, L.; Zhang, X.; Khan, R.S.A. Applications of molecular markers to develop resistance against abiotic stresses in wheat. In Wheat Production in Changing Environments; Springer: Singapore, 2019; pp. 393–420. [Google Scholar] [CrossRef]
- Kramer, P.J. Water Relation of Plants; Academic Press: Orlando, FL, USA, 1983; pp. 342–389. Available online: http://www.sciencedirect.com/science/book/9780124250406 (accessed on 6 September 2021).
- Araus, J.L.; Slafer, G.A.; Reynolds, M.P.; Royo, C. Plant Breeding and Drought in C3 Cereals: What Should We Breed For? Ann. Bot. 2002, 89, 925–940. [Google Scholar] [CrossRef]
- Mitra, J. Genetics and genetic improvement of drought resistance in crop plants. Curr. Sci. 2001, 80, 758–763. [Google Scholar]
- Datta, J.; Mondal, T.; Banerjee, A.; Mondal, N. Assessment of drought tolerance of selected wheat cultivars under laboratory condition. J. Agric. Technol. 2011, 7, 383–393. [Google Scholar]
- Nowsherwan, I.; Shabbir, G.; Malik, S.I.; Ilyas, M.; Iqbal, M.S.; Musa, M. Effect of drought stress on different physiological traits in bread wheat. SAARC J. Agric. 2018, 16, 1–6. [Google Scholar] [CrossRef]
- Bayoumi, T.Y.; Eid, M.H.; Metwali, E.M. Application of physiological and biochemical indices as a screening technique for drought tolerance in wheat genotypes. Afr. J. Biotechnol. 2008, 7, 2341–2352. [Google Scholar]
- Dawood, M.F.A.; Abeed, A.H.A.; Aldaby, E.E.S. Titanium dioxide nanoparticles model growth kinetic traitsof some wheat cultivars under different water regimes. Indian J. Plant Physiol. 2019, 24, 129–140. [Google Scholar] [CrossRef]
- Bowne, J.B.; Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.; Roessner, U. Drought responses of leaf tissues from wheat cultivars of differing drought tolerance at the metabolite level. Mol. Plant 2012, 5, 418–429. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Xin, Z.; Yang, T.; Ma, X.; Zhang, Y.; Wang, Z.; Ren, Y.; Lin, T. Metabolomics Response for Drought Stress Tolerance in Chinese Wheat Genotypes (Triticum aestivum). Plants 2020, 9, 520. [Google Scholar] [CrossRef] [Green Version]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and Drought Stresses in Crop Plants: Implications, Cross Talk, and Potential Management Opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Raza, A.; Charagh, S.; Sadaqat, N.; Jin, W. Arabidopsis thaliana: Model plant for the study of abiotic stress responses. In The Plant Family Brassicaceae; Springer: Singapore, 2020; pp. 129–180. [Google Scholar] [CrossRef]
- Blum, A.; Pnuel, Y. Physiological attributes associated with drought resistance of wheat cultivars in a Mediterranean environment. Aust. J. Agric. Res. 1990, 41, 799–810. [Google Scholar] [CrossRef]
- FAO-UNDP. Land Resources Appraisal of Bangladesh for Agricultural Development: Agro-Ecological Regions of Bangladesh; Technical Report No. 3, Volume II; United Nations Development Programme (UNDP)-Food and Agriculture Organization (FAO): Rome, Italy, 1988. [Google Scholar]
- BARC (Bangladesh Agricultural Research Council). Fertilizer Recommendation Guide: Agroecological Regions of Bangladesh; Bangladesh Agricultural Research Council: Dhaka, Bangladesh, 2012; p. 251.
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Barr, H.D.; Weatherley, P.E. A reexamination of the relative turgidity technique for estimating water deficit in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Witham, H.; Blades, D.F.; Devin, R.H. Exercise in Plant Physiology, 2nd ed.; PWS Publishers: Boston, MA, USA, 1986; pp. 128–131. [Google Scholar]
- Ayeneh, A.; Ginkel, M.; Reynolds, M.P.; Ammar, K. Comparison of leaf, spike, peduncle, and canopy temperature depression in wheat under heat stress. Field Crop. Res. 2002, 79, 173184. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. Available online: https://link.springer.com/article/10.1007/BF00018060 (accessed on 6 September 2021). [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedure for Agricultural Research, 2nd ed.; John Wiley & Sons: Los Baños, Philippines, 1984; pp. 28–192. [Google Scholar]
- Maman, N.; Lyon, D.J.; Mason, S.; Galusha, T.D.; Higgins, R. Pearl millet and grain sorghum yield response to water supply in Nebraska. Panhand. Res. Ext. Cent. 2003, 8. [Google Scholar] [CrossRef] [Green Version]
- Sokoto, M.B.; Abubakar, I.U.; Singh, A. Effect of Water Stress and Variety on Phenology of Bread Wheat (Triticumaestivum L.) in Sokoto, Sudan Savannah, Nigeria. J. Dryland Agric. 2015, 1, 33–43. [Google Scholar]
- Riaz, R. Genetic Analysis for Yield and Yield Components in Spring Wheat under Drought Conditions. Ph.D. Thesis, Department of Agriculture, Faisalabad Agriculture University, Faisalabad, Pakistan, 2003; p. 35. [Google Scholar]
- Majer, P.; Sass, L.; Lelley, T.; Cseuz, L.; Vass, I.; Dudits, D.; Pauk, J. Testing drought tolerance of wheat by a complex stress diagnostic system installed in greenhouse. Acta Biol. Szeged. 2008, 52, 97–100. [Google Scholar]
- Bilal, M.; Rashid, R.; Rehman, S.; Iqbal, F.; Ahmed, J.; Abid, M.; Ahmed, Z.; Hayat, A. Evaluation of wheat genotypes for drought tolerance. J. Green Physiol. Genet. Genom. 2015, 1, 11–21. [Google Scholar]
- Allahverdiyev, T.I. Effect of drought stress on some physiological parameters, yield, yield components of durum (Triticum durum desf.) and bread (Triticum aestivum L.) wheat genotypes. Ekin J. Crop Breed. Genet. 2015, 1, 50–62. [Google Scholar]
- Siddique, K.H.M.; Tennant, D.; Perry, M.W.; Belford, R.K. Water use and water use efficiency of old and modern wheat cultivars in a Mediterranean-type environment. Aust. J. Agril. Res. 1990, 41, 431–447. [Google Scholar] [CrossRef]
- Poudel, M.R.; Ghimire, S.; Pandey, M.P.; Dhakal, K.H.; Thapa, D.B.; Poudel, H.K. Evaluation of Wheat Genotypes under Irrigated, Heat Stress and Drought Conditions. J. Biol. Today’s World 2020, 9, 212. [Google Scholar]
- Khakwani, A.A.; Dennett, M.D.; Munir, M.; Abid, M. Growth and yield response of wheat varieties to water stress at booting and anthesis stages of development. Pak. J. Bot. 2012, 44, 879–886. [Google Scholar]
- Siddique, K.H.M.; Loss, S.P.; Thomson, B.D.; Saxena, N.P. Cool season grain legumes in dryland Mediterranean environments of Western Australia: Significance of early flowering. In Management of Agricultural Drought: Agronomic and Genetic Options; Saxena, N.P., Ed.; Oxford University Press: New Delhi, India, 2003; pp. 151–162. [Google Scholar]
- Kirda, C.; Kanber, R.; Tulucu, K. Yield response of cotton, maize, soybean, sugar beet, sunflower, and wheat to deficit irrigation. In Crop Yield Response to Deficit Irrigations; Kluwer Academic: Dordrecht, The Netherlands, 1999; pp. 21–38. [Google Scholar]
- Gonzalez, A.; Bermejo, V.; Gimeno, B.S. Effect of different physiological traits on grain yield in barley grown under irrigated and terminal water deficit conditions. J. Agric. Sci. 2010, 148, 319–328. [Google Scholar] [CrossRef]
- Lopes, M.S.; Reynolds, M.P. Stay-green in spring wheat can be determined by spectral reflectance measurements (normalized difference vegetation index) independently from phenology. J. Exp. Bot. 2012, 63, 3789. [Google Scholar] [CrossRef] [Green Version]
- Mehraban, A.; Ahmad Tobe, A.; Gholipouri, A.; Amiri, E.; Ghafari, A.; MozffarRostaii, M. The Effects of Drought Stress on Yield, Yield Components, and Yield Stability at Different Growth Stages in Bread Wheat Cultivar (Triticum aestivum L.). Pol. J. Environ. Stud. 2019, 28, 739–746. [Google Scholar] [CrossRef]
- Nezhadahmadi, A.; Prodhan, Z.; Faruq, G. Drought Tolerance in Wheat. Sci. World J. 2013, 2013, 610721. [Google Scholar] [CrossRef] [Green Version]
- Rajala, A.; Hakala, K.; Mäkelä, P.; Peltonen-Sainio, P. Drought effect on grain number and grain weight at spike and spikelet level in six-row spring barley. J. Agron. Crop Sci. 2011, 197, 103–112. [Google Scholar] [CrossRef]
- Kilic, H.; Yağbasanlar, T. The effect of drought stress on grain yield, yield components and some quality traits of durum wheat (Triticum turgidum) cultivars. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2010, 38, 164–170. [Google Scholar] [CrossRef]
- Afzal, I.; Imran, S.; Javed, T.; Basra, S.M.A. Evaluating the integrative response of moringa leaf extract with synthetic growth promoting substances in maize under early spring conditions. S. Afr. J. Bot. 2020, 132, 378–387. [Google Scholar] [CrossRef]
- Motzo, R.; Giunta, F. The Effect of breeding on the phenology of Italian durum wheats: From landraces to modern cultivars. Eur. J. Agron. 2007, 26, 462–470. [Google Scholar] [CrossRef]
- Attarbashi, M.R.; Galeshi, S.; Soltani, A.; Zinali, E. Relationship of phenology and physiological traits with grain yield in wheat under rain-fed conditions. Iran. J. Agric. Sci. 2002, 33, 8–21. [Google Scholar]
- Peterman, L.T.; Sears, R.G.; Kanemasu, E.T. Rate and duration of spikelet initiation in 10 winter wheat cultivar. Crop Sci. J. 1985, 25, 222–225. [Google Scholar] [CrossRef]
- Sial, M.A.; Dahot, M.U.; Arain, M.A.; Markhand, G.S.; Naqvi, M.H.; Laghari, K.A.; Mirbahar, A.A. Effect of water stress on yield and yield components of semi-dwarf bread wheat (Triticum aestivum L.). Pak. J. Bot. 2009, 41, 1715–1728. [Google Scholar]
- Belay, G.A.; Zhang, Z.; Xu, P. Physio-Morphological and Biochemical Trait-Based Evaluation of Ethiopian and Chinese Wheat Germplasm for Drought Tolerance at the Seedling Stage. Sustainability 2021, 13, 4605. [Google Scholar] [CrossRef]
- Boyer, J.S.; James, R.A.; Munns, R.; Condon, T.; Passioura, J.B. Osmotic adjustment leads to anomalously low estimates of relative water content in wheat and barley. Funct. Plant Biol. 2008, 35, 1172–1182. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Patel, A.L. Water status, gaseous exchange, proline accumulation and yield of wheat un response to water stress. Ann. Biol. Ludhiana 1996, 12, 26–28. [Google Scholar]
- Lonbani, M.; Arzani, A. Morpho-physiological traits associated with terminal drought-stress tolerance in triticale and wheat. Agron. Res. 2011, 9, 315–329. [Google Scholar]
- Teulat, B.; Monneveux, P.; Wery, J.; Borries, C.; Souyris, I.; Charrier, A.; This, D. Relationships between relative water content and growth parameters under water stress in barley: A QTL study. New Phytol. 1997, 137, 99–107. [Google Scholar] [CrossRef]
- Hasheminasab, H.; Assad, A.M.; Aliakbari, T.; Sahhafi, S.R. Evaluation of some physiological traits associated with improved drought tolerance in Iranian wheat. Ann. Biol. Res. 2012, 3, 1719–1725. [Google Scholar]
- Colom, M.R.; Vazzana, C. Photosynthesis and PSII functionality of drought-resistant and drought sensitive weeping love grass plants. Environ. Exp. Bot. 2003, 49, 135–144. [Google Scholar] [CrossRef]
- Sinclair, T.; Ludlow, M. Who taught plants thermodynamics? The unfulfilled potential of plant water potential. Aust. J. Plant Physiol. 1985, 12, 213–217. [Google Scholar] [CrossRef]
- Tatar, O.; Gevrek, M.N. Influence of water stress on prolin accumulation, lipid peroxidation and water content of wheat. Asian J. Plant Sci. 2008, 7, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Siddique, M.R.B.; Hamid, A.; Aslam, M.S. Drought stress effects on water relations of wheat. Bot. Bull. Acad. Sin. 2000, 41, 35–39. [Google Scholar]
- Ritchie, S.W.; Nguyan, H.T.; Holaday, A.S. Leaf Water content and gas exchange parameters of two wheat genotypes differing in drought resistance. Crop Sci. 1990, 30, 105–111. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Afkhami, N. Changes in some morpho-physiological traits of safflower in response to water deficit and nano-fertilizers. J. Bio. Environ. Sci. 2018, 12, 391–398. [Google Scholar]
- Mamnabi, S.; Safar Nasrollahzadeh, S.; Ghassemi-Golezani, K.; Raei, M. Improving yield-related physiological characteristics of spring rapeseed by integrated fertilizer management under water deficit conditions. Saudi J. Biol. Sci. 2020, 27, 797–804. [Google Scholar] [CrossRef]
- Schonfeld, M.A.; Johnson, R.C.; Carver, B.F.; Mornhinweg, D.W. Water relations in winter wheat as drought resistance indicators. Crop Sci. 1988, 28, 526–531. [Google Scholar] [CrossRef]
- Almeselmani, M.; Abdullah, F.; Hareri, F.; Naaesan, M.; Ammar, M.A.; ZuherKanbar, O.; Saud, A.A. Effect of drought on different physiological characters and yield component in different varieties of Syrian durum wheat. J. Agric. Sci. 2011, 3, 127–133. [Google Scholar] [CrossRef]
- Sairam, R.K.; Deshmukh, P.S.; Shukla, D.S. Increased Antioxidant Enzyme Activity in Response to Drought and Temperature Stress Related with Stress Tolerance in Wheat Genotypes. Abstract: National Seminar (ISSP); IARI: New Delhi, India, 1997; p. 69. [Google Scholar]
- Manivannan, P.; Jaleel, C.A.; Sankar, B.; Kishorekumar, A.; Somasundaram, R.; Lakshmanan, G.A.; Panneerselvam, R. Growth, biochemical modifications and proline metabolism in Helianthus annuus L. as induced by drought stress. Colloids Surf. B Biointerfaces 2007, 59, 141–159. [Google Scholar] [CrossRef] [PubMed]
- Bijanzadeh, E.; Emam, Y. Effect of Defoliation and drought stress on yield components and chlorophyll content of wheat. Pak. J. Biol. Sci. 2010, 13, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.B.; Houghton, J.D.; Hendry, G.A.F. Chlorophyll breakdown. In Chlorophyllus; Scheer, H., Ed.; CRC Press: Boca Raton, FL, USA, 1991; p. 465. [Google Scholar]
- Javed, T.; Shabbir, R.; Ali, A.; Afzal, I.; Zaheer, U.; Gao, S.J. Transcription factors in plant stress responses: Challenges and potential for sugarcane improvement. Plants 2020, 9, 491. [Google Scholar] [CrossRef] [PubMed]
- Farshadfar, E.; Ghasemi, M.; Rafii, F. Evaluation of physiological parameters as a screening technique for drought tolerance in bread wheat. J. Biodiv. Environ. Sci. 2014, 4, 175–186. [Google Scholar]
- Sikuku, P.A.; Netondo, G.W.; Onyango, J.C.; Musyimi, D.M. Chlorophyll fluorescence, protein and chlorophyll content of three nerica rainfed rice varieties under varying irrigation regimes. J. Agric. Biol. Sci. 2010, 5, 19–25. [Google Scholar]
- Rana, M.S.; Hasan, M.A.; Bahadur, M.M.; Islam, M.R. Physiological evaluation of wheat genotypes for tolerance to water deficit stress. Bangladesh Agron. J. 2017, 20, 37–52. [Google Scholar] [CrossRef]
- Medrano, H.; Escalona, J.M.; Bota, J.; Gulias, J.; Flexas, J. Regulation of photosynthesis of C3 plants in response to progressive drought: Stomatal conductance as a reference parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef]
- Wright, H.; De Longa, J.; Ladab, R.; Prangea, R. The relationship between water status and chlorophyll a fluorescence in grapes (Vitis spp.). Postharvest Biol. Technol. 2009, 51, 193–199. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Energy dissipation in C3 plants under drought. Funct. Plant Biol. 2002, 29, 1209–1215. [Google Scholar] [CrossRef]
- Herzog, H. Source and Sink during Reproductive Period of Wheat; Scientific Publishers: Berlin/Hamburg, Germany, 1986; pp. 147–148. [Google Scholar]
- Hosseinzadeh, S.R.; Amiri, H.; Ismaili, A. Evaluation of photosynthesis, physiological, and biochemical responses of chickpea (Cicer arietinum L. cv. Pirouz) under water deficit stress and use of vermicompost fertilizer. J. Integr. Agric. 2018, 17, 2426–2437. [Google Scholar] [CrossRef] [Green Version]
- Salehi, A.; Tasdighi, H.; Gholamhoseini, M. Evaluation of proline, chlorophyll, soluble sugar content and uptake of nutrients in the German chamomile (Matricaria chamomilla L.) under drought stress and organic fertilizer treatments. Asian Pac. J. Trop. Biomed. 2016, 6, 886–891. [Google Scholar] [CrossRef] [Green Version]
- Navari-Izzo, F.; Quartacci, M.F.; Izzo, R. Water stress induced changes in protein and free amino acids in field grown maize and sunflower. Plant. Physiol. Biochem. 1990, 28, 531–537. [Google Scholar]
- Buttar, G.S.; Singh, C.J.; Ahuja, M.S.; Saini, K.S. Canopy temperature: A method to estimate plant water stress and scheduling irrigation in cotton and wheat. J. Agric. Physics. 2005, 5, 79–83. [Google Scholar]
- Jackson, R.D.; Idso, S.B.; Reginato, R.J.; Pinter, P.J., Jr. Canopy temperature as a crop water stress indicator. Water Resour. Res. 1981, 17, 1133–1138. [Google Scholar] [CrossRef]
- Maes, W.H.; Steppe, K. Estimating evapotranspiration and drought stress with ground-based thermal remote sensing in agriculture: A review. J. Exp. Bot. 2012, 63, 4671–4712. [Google Scholar] [CrossRef] [Green Version]
- Lopes, M.S.; Reynolds, M.P. Partitioning of assimilates to deeper roots is associated with cooler canopies and increased yield under drought in wheat. Funct. Plant Biol. 2010, 37, 147–156. [Google Scholar] [CrossRef]
- Blum, A.; Shipiler, L.; Golan, G.; Mayer, J. Yield stability and canopy temperature of wheat Osmotic adjustment and growth of barley genotypes under drought stress. Crop Sci. 1989, 29, 230–233. [Google Scholar] [CrossRef]
- Gurumurthy, S.; Sarkar, B.; Vanaja, M.; Lakshmi, J.; Yadav, S.K.; Maheswari, M. Morpho-physiological and biochemical changes in black gram (Vigna mungo L. Hepper) genotypes under drought stress at flowering stage. Acta Physiol. Plant. 2019, 41, 42. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Mwadzingeni, L.; Shimelis, H.; Tesfay, S.; Tsilo, T.J. Screening of bread wheat genotypes for drought tolerance using phenotypic and proline analyses. Front. Plant Sci. 2016, 7, 1276. [Google Scholar] [CrossRef] [Green Version]
- Delauney, A.J.; Verma, D.P.S. Proline biosynthesis and osmoregulation in plants. Plant J. 1993, 4, 215–223. [Google Scholar] [CrossRef]
- Marcińska, I.; Czyczyło-Mysza, I.; Skrzypek, E.; Filek, M.; Grzesiak, S.; Grzesiak, M.T.; Janowiak, F.; Hura, T.; Dziurka Michałand Dziurka, K.; Nowakowska, A.; et al. Impact of osmotic stress on physiological and biochemical characteristics in drought-susceptible and drought-resistant wheat genotypes. Acta Physiol. Planta 2013, 35, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Sultan, M.A.R.F.; Faisal, R.; Hui, L.; Yang, L.J.; Xian, Z.H. Assessment of Drought Tolerance of Some Triticum aestivum L. Species through Physiological Indices. Czech J. Genet. Plant Breed. 2012, 48, 178–184. [Google Scholar] [CrossRef]
- Wasaya, A.; Manzoor, S.; Yasir, T.A.; Sarwar, N.; Mubeen, K.; Ismail, I.A.; Raza, A.; Rehman, A.; Hossain, A.; EL Sabagh, A. Evaluation of Fourteen Bread Wheat (Triticum aestivum L.) Genotypes by Observing Gas Exchange Parameters, Relative Water and Chlorophyll Content, and Yield Attributes under Drought Stress. Sustainability 2021, 13, 4799. [Google Scholar] [CrossRef]
- Hasan, M.A.; Ahmed, J.U.; Bahadur, M.M.; Haque, M.M.; Sikder, S. Effect of late planting heat stress on membrane thermostability, proline content and heat susceptibility index of different wheat cultivars. J. Nat. Sci. Foun. Srilanka 2007, 35, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Budak, H.; Kantar, M.; Kurtoglu, K.Y. Drought tolerance in modern and wild wheat. Sci. World J. 2013, 2013, 548246. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M. Response of durum and bread wheat genotypes to drought stress: Biomass and yield components. Asian J. Plant Sci. 2003, 2, 290–293. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.; Osorio, M.L.; Carvalho, J.; Faria, T.; Pinheiro, C. How Plants Cope with Water Stress in the Field? Photosynthesis and Growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Bao, W. Influence of water deficit and genotype on photosynthetic activity, dry mass partitioning and grain yield changes of winter wheat. Afr. J. Agril. Res. 2011, 6, 5567–5574. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef]
- Tang, A.C.; Kawamitsa, Y.; Kanechi, M.; Boyer, J.S. Photosynthesis at low water potentials in leaf discs lacking epidermis. Ann. Bot. 2002, 89, 861–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serragoa, R.A.; Alzuetaa, I.; Savinb, R.; Slafer, G.A. Understanding grain yield responses to source-sink ratios during grain filling in wheat and barley under contrasting environments. Field Crop. Res. 2013, 150, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Maralian, F.; Ebadi, A.; Didar, T.R.; Haji-Eghrari, B. Influence of water stress on wheat grain yield and proline accumulation rate. Afr. J. Agril. Res. 2010, 5, 286–289. [Google Scholar] [CrossRef]
- Li, Y.P.; Ye, W.; Wang, M.; Yan, X.D. Climate change and drought: A risk assessment of crop yield impacts. Clim. Res. 2009, 39, 31–46. [Google Scholar] [CrossRef]
- Sallam, A.; Alqudah, A.M.; Mona, F.A.; Dawood, M.F.A.; Baenziger, P.S.; Andreas Börner, A. Drought Stress Tolerance in Wheat and Barley: Advances in Physiology, Breeding and Genetics Research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef] [Green Version]
- Youldash, K.M.; Barutcular, C.; EL Sabagh, A.; Toptas, I.; Kayaalp, G.T.; Hossain, A.; Alharby, H.; Bamagoos, A.; Saneoka, H.; Farooq, M. Evaluation of grain yield in fifty-eight spring bread wheat genotypes grown under heat stress. Pak. J. Bot. 2020, 52, 33–42. [Google Scholar] [CrossRef]
- Farooq, M.O.; Kashif, M.; Khaliq, I.; Ahmed, N. Phenotypic Selection of Wheat Genotypes for Drought Stress Tolerance. Int. J. Agric. Biol. 2020, 23, 509–514. [Google Scholar] [CrossRef]
- Farooq, S.; Shahid, M.; Khan, M.B.; Hussain, M.; Farooq, M. Improving the productivity of bread wheat by good management practices under terminal drought. J. Agron. Crop Sci. 2015, 201, 173–188. [Google Scholar] [CrossRef]
- Qadir, S.A.; Khursheed, M.Q.; Huyop, F.Z. Drought tolerance and genetic diversity among selected wheat cultivars. Zanco J. Pure Appl. Sci. 2017, 29, 110–117. Available online: https://zancojournals.su.edu.krd/index.php/JPAS/article/view/893 (accessed on 6 September 2021).
- Nazir, M.F.; Sarfraz, Z.; Mangi, N.; Nawaz Shah, M.K.; Mahmood, T.; Mahmood, T.; Iqbal, M.S.; Ishaq Asif Rehmani, M.; El-Sharnouby, M.; Shabaan, M.K.A.; et al. Post-Anthesis Mobilization of Stem Assimilates in Wheat under Induced Stress. Sustainability 2021, 13, 5940. [Google Scholar] [CrossRef]
- Yasir, T.A.; Wasaya, A.; Hussain, M.; Ijaz, M.; Farooq, M.; Farooq, O.; Nawaz, A.; Hu, Y.G. Evaluation of physiological markers for assessing drought tolerance and yield potential in bread wheat. Physiol. Mol. Biol. Plants 2019, 25, 1163–1174. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| General Characters | Description |
|---|---|
| Location | Crop Physiology and Ecology, HSTU, Dinajpur |
| AEZ | Old Himalayan Piedmont Plain (AEZ-1) |
| General Soil type | Non-Calcareous Brown Floodplain Soil |
| Parent material | Piedmont alluvium |
| Soil series | Ranishankail |
| Drainage | Moderately well-drained |
| Flood level | Above flood level |
| Topography | High land |
| Soil Characters | Analytical Value | Critical Level | Soil Test Values Interpretation | Range of Value Used within the Interpretation Class |
|---|---|---|---|---|
| Physical properties | ||||
| Sand (%) | 60.00 | - | - | - |
| Silt (%) | 27.00 | - | - | - |
| Clay (%) | 13.00 | - | - | - |
| Textural class | Sandy loam | - | - | - |
| Bulk density(g/cc) | 0.86–1.07 | - | - | - |
| Field capacity (%) | 25.8 | - | - | - |
| Permanent wilting point | 12.0 | - | - | - |
| Chemical properties | ||||
| Soil pH (1:1.25, Soil: H2O) | 5.40–5.50 | - | - | - |
| Organic matter (%) (Wet oxidation method) | 0.69 | - | Low | - |
| CEC (meq/100g soil) (BaCl2- compulsive exchange method) | 5.60 | - | Low | - |
| Total nitrogen (%) (Micro-Kjeldahl method) | 0.07 | 0.12 | Low | ≤0.09 |
| Available phosphorus (µg g−1) (Molybdate blue ascorbic acid method) | 16.75 | 10.00 | Medium | 7.51–15.0 |
| Exchangeable potassium (meq100 g−1 soil) (Flame photometer method) | 0.17 | 0.12 | Low | 0.18–0.27 |
| Available sulphur (µg g−1) (Turbidity method using BaCl2) | 17.53 | 10.00 | Medium | 15.10–22.50 |
| Available boron (µg g−1) (Calcium chloride extraction method) | 0.15 | 0.20 | Very low | ≤0.15 |
| Available zinc (µg g−1) (Atomic Absorption Spectrophotometer method) | 0.88 | 0.60 | Low | 0.45–0.90 |
| Months | Relative Humidity (%) | Temperature | Total Rainfall (mm) | |
|---|---|---|---|---|
| Minimum (°C) | Maximum (°C) | |||
| November | 87 | 15.9 | 25.6 | 0.0 |
| December | 85 | 10.9 | 21.9 | 0.0 |
| January | 72 | 9.6 | 18.8 | 4.0 |
| February | 78 | 14.7 | 25.2 | 8.0 |
| March | 81 | 17.5 | 29.7 | 3.0 |
| April | 89 | 22.7 | 30.9 | 33.0 |
| Sr. No. | Genotypes | Pedigree/Selection History |
|---|---|---|
| 1 | BARI Gom 26 (BAW 1064) | Variety, ICTAL123/3/RAWAL87//VEE/HD2285 BD(JOY) 86-0JO-3JE-010JE-010JE-HRDI-RC5DI |
| 2 | BAW 1158 | BAW 968/SHATABDI |
| 3 | BAW 1167 | BL 3877 = KAUZ/STAR/CMH 81.749//BL 2224 |
| 4 | BAW 1169 | SHATABDI/BAW 923 |
| Treatment | Days to Booting | Days to Heading | Days to Anthesis | Days to Physiological Maturity | |
|---|---|---|---|---|---|
| Water levels | |||||
| Well watered | 51.21 a | 63.54 a | 75.22 a | 107.78 a | |
| Water stress | 49.11 a | 59.23 b | 70.54 b | 101.41 b | |
| CV (%) | 1.41 | 1.52 | 1.77 | 1.57 | |
| Genotypes | |||||
| BARI Gom 26 | 50.23 b | 59.85 b | 71.24 b | 103.58 b | |
| BAW 1158 | 54.65 a | 63.56 a | 75.63 a | 105.11 a | |
| BAW 1167 | 46.42 b | 58.96 b | 68.42 b | 102.62 b | |
| BAW 1169 | 55.33 a | 66.23 a | 75.63 a | 106.21 a | |
| CV (%) | 1.24 | 2.00 | 0.75 | 0.66 | |
| Genotypes × Water levels | |||||
| BARI Gom 26 | WW | 49.72 b | 60.23 b | 75.22 a | 106.62 a |
| DS | 46.85 c | 56.63 c | 70.06 b | 99.15 c | |
| BAW 1158 | WW | 56.61 a | 65.43 a | 74.23 a | 107.98 a |
| DS | 54.96 ab | 63.85 a | 72.40 ab | 104.16 ab | |
| BAW 1167 | WW | 48.96 b | 59.11 b | 72.23 ab | 104.25 b |
| DS | 45.65 c | 54.63 c | 66.30 c | 94.04 d | |
| BAW 1169 | WW | 57.23 a | 67.11 a | 75.84 a | 108.83 a |
| DS | 56.42 a | 65.81 a | 74.16 a | 105.77 a | |
| CV (%) | 1.24 | 2.00 | 0.75 | 0.66 | |
| Genotypes | Water Levels | Relative Water Content (RWC) | |
|---|---|---|---|
| % | % Change Over Normal | ||
| BARI Gom 26 | Well watered | 87.96 b | - |
| Water stress | 81.19 d | −7.70 | |
| BAW 1158 | Well watered | 88.17 ab | - |
| Water stress | 85.11 c | −3.47 | |
| BAW 1167 | Well watered | 91.41 a | - |
| Water stress | 83.71 cd | −8.42 | |
| BAW 1169 | Well watered | 89.56 a | - |
| Water stress | 87.24 b | −2.59 | |
| Level of significance | * | ||
| CV (%) | 1.28 | ||
| Genotypes | Water Levels | Chlorophyll Content in Flag Leaf at 8 DAA | Chlorophyll Content in Flag Leaf at 24 DAA | ||
|---|---|---|---|---|---|
| mg g−1 FW | % Change Over Normal | mg g−1 FW | % Change Over Normal | ||
| BARI Gom 26 | Well watered | 2.55 a | - | 2.09 b | - |
| Water stress | 2.11 cd | −17.25 | 1.38 d | −33.97 | |
| BAW 1158 | Well watered | 2.44 b | - | 2.20 a | - |
| Water stress | 2.20 c | −9.76 | 1.85 c | −15.91 | |
| BAW 1167 | Well watered | 2.49 b | - | 2.15 ab | - |
| Water stress | 2.06 d | −16.60 | 1.45 d | −32.56 | |
| BAW 1169 | Well watered | 2.62 a | - | 2.29 a | - |
| Water stress | 2.40bc | −8.40 | 1.95 bc | −14.85 | |
| Level of significance | ** | * | |||
| CV (%) | 2.20 | 3.50 | |||
| Genotypes | Water Levels | Canopy Temperature Depression (CTD) | |
|---|---|---|---|
| (°C) | % Change Over Control | ||
| BARI Gom 26 | WW | 9.40 ab | - |
| DS | 5.97 e | −36.49 | |
| BAW 1158 | WW | 8.60 c | - |
| DS | 6.30 e | −26.74 | |
| BAW 1167 | WW | 8.73 bc | - |
| DS | 5.13 f | −41.23 | |
| BAW 1169 | WW | 9.53 a | - |
| DS | 7.27 d | −23.71 | |
| Level of significance | ** | - | |
| CV (%) | 3.82 | - | |
| Genotypes | Water Levels | Proline Content in Flag Leaf | Proline Content in Kernel | ||
|---|---|---|---|---|---|
| µmole g−1 FW | % Change Over Normal | µmole g−1 FW | % Change Over Normal | ||
| BARI Gom 26 | Well watered | 1.90 c | - | 2.60 cd | - |
| Water stress | 2.10 a | 10.53 | 2.71 c | 4.25 | |
| BAW 1158 | Well watered | 1.81 d | - | 2.50 d | - |
| Water stress | 2.20 a | 21.55 | 2.95 b | 18.00 | |
| BAW 1167 | Well watered | 1.83 d | - | 2.54 d | - |
| Water stress | 1.99 b | 8.19 | 2.64 c | 3.94 | |
| BAW 1169 | Well watered | 1.79 d | - | 2.58 d | - |
| Water stress | 2.22 a | 24.02 | 3.10 a | 20.16 | |
| Level of significance | * | * | |||
| CV (%) | 4.04 | 2.56 | |||
| Genotypes | Water Levels | Grain Dry Weight Spike−1 | Grain Yield | ||
|---|---|---|---|---|---|
| g Spike−1 | % Change Over Normal | t ha−1 | % Change Over Normal | ||
| BARI Gom 26 | Well watered | 3.60 a | 5.58 a | - | |
| Water stress | 3.30 b | −8.33 | 3.64 d | −34.77 | |
| BAW 1158 | Well watered | 2.82 d | - | 5.56 a | - |
| Water stress | 2.76 d | −2.13 | 4.23 c | −23.92 | |
| BAW 1167 | Well watered | 2.97 c | - | 5.07 b | - |
| Water stress | 2.62 e | −11.78 | 3.27 d | −35.50 | |
| BAW 1169 | Well watered | 3.07 c | - | 5.65 a | - |
| Water stress | 2.97 c | −3.26 | 4.39 c | −22.30 | |
| Level of significance | ** | - | ** | - | |
| CV (%) | 1.88 | - | 2.67 | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chowdhury, M.K.; Hasan, M.A.; Bahadur, M.M.; Islam, M.R.; Hakim, M.A.; Iqbal, M.A.; Javed, T.; Raza, A.; Shabbir, R.; Sorour, S.; et al. Evaluation of Drought Tolerance of Some Wheat (Triticum aestivum L.) Genotypes through Phenology, Growth, and Physiological Indices. Agronomy 2021, 11, 1792. https://doi.org/10.3390/agronomy11091792
Chowdhury MK, Hasan MA, Bahadur MM, Islam MR, Hakim MA, Iqbal MA, Javed T, Raza A, Shabbir R, Sorour S, et al. Evaluation of Drought Tolerance of Some Wheat (Triticum aestivum L.) Genotypes through Phenology, Growth, and Physiological Indices. Agronomy. 2021; 11(9):1792. https://doi.org/10.3390/agronomy11091792
Chicago/Turabian StyleChowdhury, M. Kaium, M. A. Hasan, M. M. Bahadur, Md. Rafiqul Islam, Md. Abdul Hakim, Muhammad Aamir Iqbal, Talha Javed, Ali Raza, Rubab Shabbir, Sobhy Sorour, and et al. 2021. "Evaluation of Drought Tolerance of Some Wheat (Triticum aestivum L.) Genotypes through Phenology, Growth, and Physiological Indices" Agronomy 11, no. 9: 1792. https://doi.org/10.3390/agronomy11091792
APA StyleChowdhury, M. K., Hasan, M. A., Bahadur, M. M., Islam, M. R., Hakim, M. A., Iqbal, M. A., Javed, T., Raza, A., Shabbir, R., Sorour, S., Elsanafawy, N. E. M., Anwar, S., Alamri, S., Sabagh, A. E., & Islam, M. S. (2021). Evaluation of Drought Tolerance of Some Wheat (Triticum aestivum L.) Genotypes through Phenology, Growth, and Physiological Indices. Agronomy, 11(9), 1792. https://doi.org/10.3390/agronomy11091792