
Feeding the Microbes: A Strategy to Control Verticillium Wilt



 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pathogen Preparation
2.2. Evaluation of Potato Dextrose and Nutrient Broth against V. dahliae under Glasshouse Conditions
2.3. Split Root Bioassay
2.4. Isolation and Identification of Rhizospheric Microorganisms
2.5. Genome Assembly of Strain Z13 and Phylogenetic Reconstruction
2.6. In Vitro and in Planta Evaluation of Microorganisms against V. dahliae
2.7. Statistics
3. Results
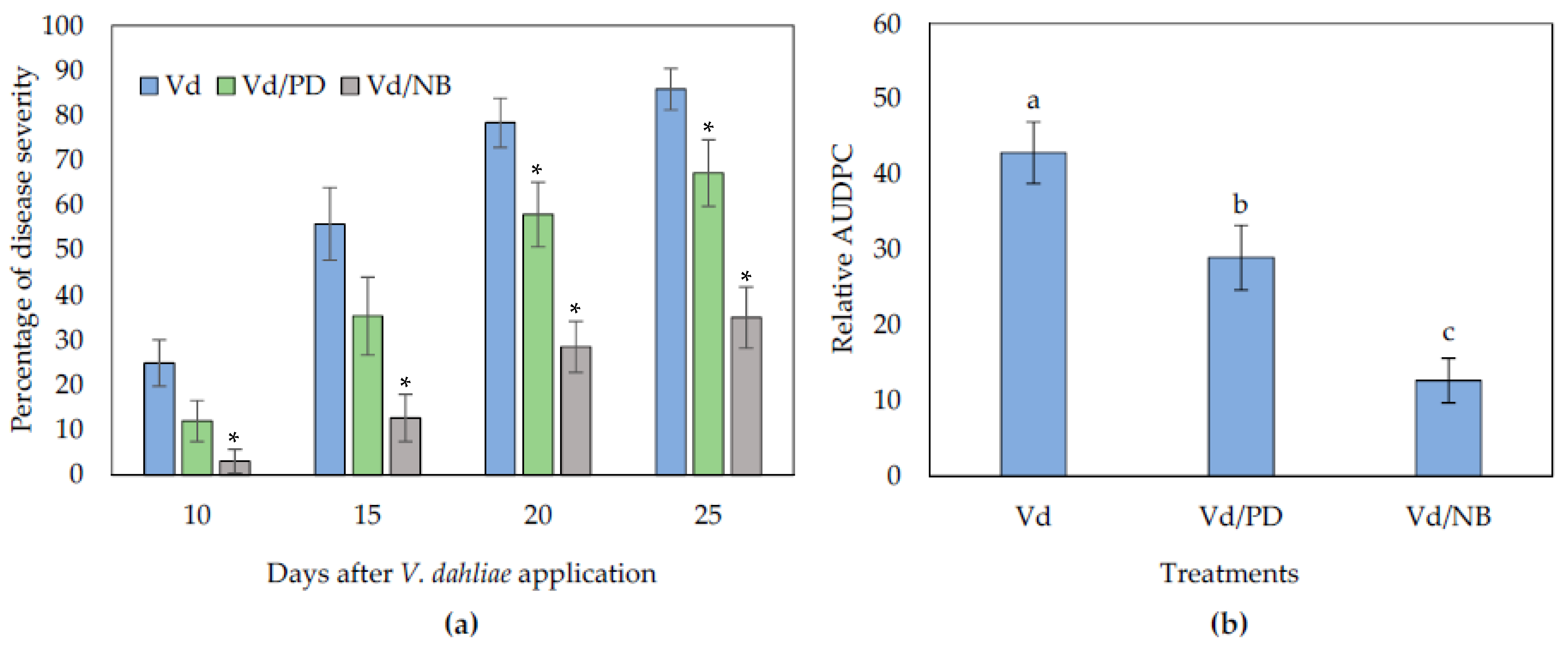
3.1. Potato Dextrose and Nutrient Broth Reduce Verticillium Wilt Symptoms in Eggplants Grown in Non-Sterile Substrate but Nutrient Broth Fails in Sterile Substrate
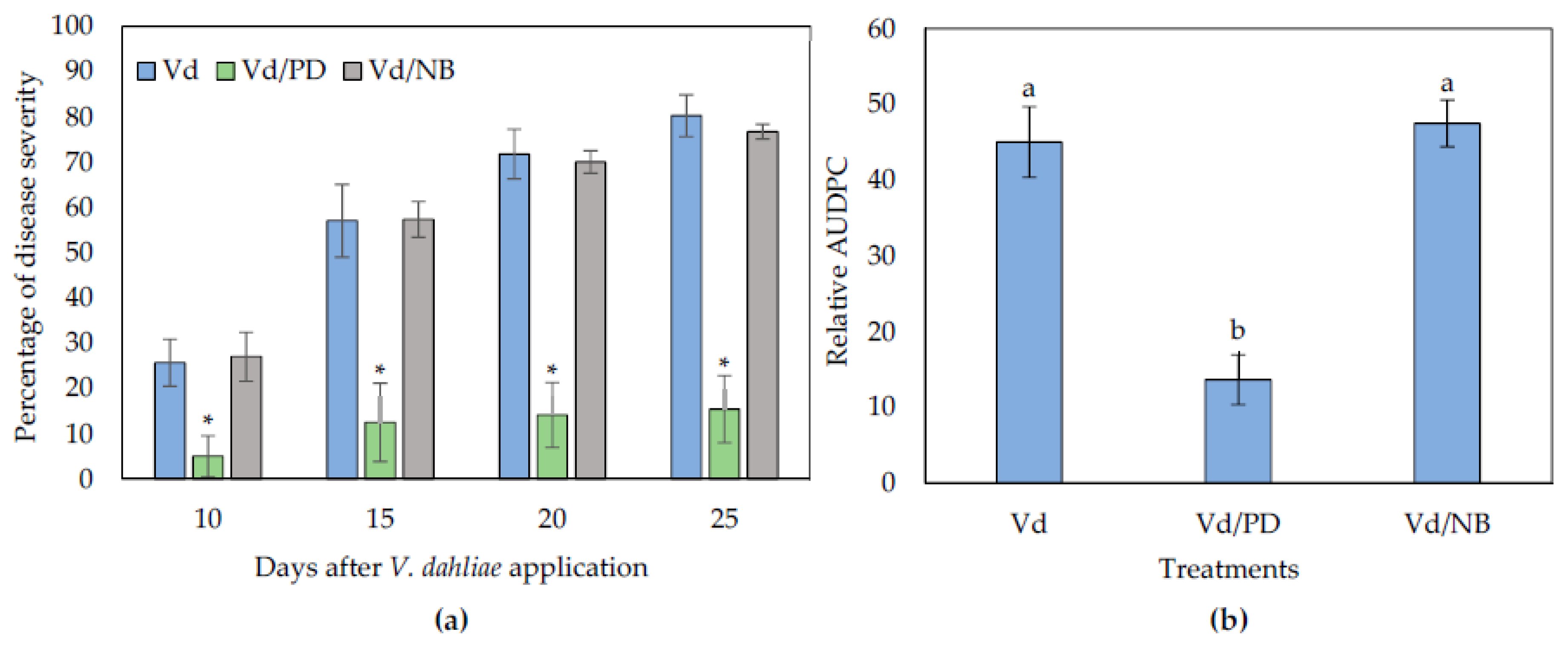
3.2. Potato Dextrose and Nutrient Broth Fail to Reduce Verticillium Wilt Symptoms in Eggplants Grown in a Split Root System
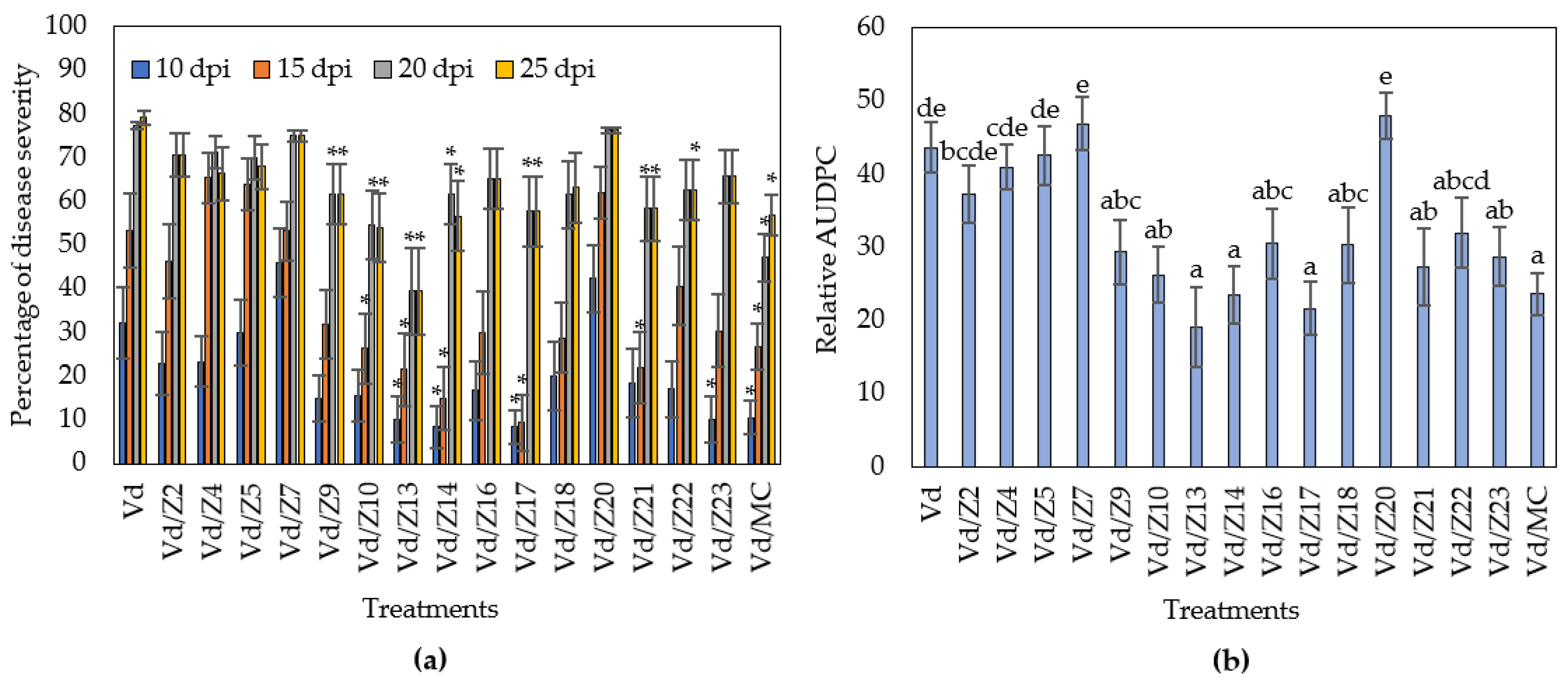
3.3. Microorganisms Isolated from the Rhizoshere of Nutrient Broth Treated Plants, Reduce Verticillium Wilt Symptoms in Eggplants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pegg, G.F.; Brady, B.L. Verticillium Wilts; CABI Publishing: New York, NY, USA, 2002. [Google Scholar]
- Schnathorst, W.C. Life cycle and epidemiology of Verticillium. In Fungal Wilt Diseases of Plants; Mace, M.A., Bell, A.A., Beckman, C.H., Eds.; Academic Press: New York, NY, USA, 1981; pp. 81–111. [Google Scholar]
- Poulaki, E.G.; Tsolakidou, M.-D.; Gkizi, D.; Pantelides, I.S.; Tjamos, S.E. The Ethylene Biosynthesis Genes ACS2 and ACS6 Modulate Disease Severity of Verticillium dahliae. Plants 2020, 9, 907. [Google Scholar] [CrossRef]
- Diwan, N.; Fluhr, R.; Eshed, Y.; Zamir, D.; Tanksley, S.D. Mapping of Ve in tomato: A gene conferring resistance to the broad-spectrum pathogen, Verticillium dahliae race 1. Theor. Appl. Genet. 1999, 98, 315–319. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Prieto, P.; Rincón, A.M.; Rodríguez, R.V.; Valderrama, R.; Barroso, J.B.; Mercado-Blanco, J. Fate of Trichoderma harzianum in the olive rhizosphere: Time course of the root colonization process and interaction with the fungal pathogen Verticillium dahliae. BioControl 2016, 61, 269–282. [Google Scholar] [CrossRef]
- Cabanás, C.G.-L.; Ruano-Rosa, D.; Legarda, G.; Pizarro-Tobías, P.; Valverde-Corredor, A.; Triviño, J.C.; Roca, A.; Mercado-Blanco, J. Bacillales Members from the Olive Rhizosphere Are Effective Biological Control Agents against the Defoliating Pathotype of Verticillium dahliae. Agriculture 2018, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Tjamos, S.E.; Flemetakis, E.; Paplomatas, E.J.; Katinakis, P. Induction of Resistance to Verticillium dahliae in Arabidopsis thaliana by the Biocontrol Agent K-165 and Pathogenesis-Related Proteins Gene Expression. Mol. Plant-Microbe Interact. 2005, 18, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Deketelaere, S.; Tyvaert, L.; França, S.D.C.; Höfte, M. Desirable Traits of a Good Biocontrol Agent against Verticillium Wilt. Front. Microbiol. 2017, 8, 1186. [Google Scholar] [CrossRef] [Green Version]
- Gkizi, D.; Gil, A.G.; Pardal, A.J.; Piquerez, S.J.M.; Sergaki, C.; Ntoukakis, V.; Tjamos, S.E. The bacterial biocontrol agent Paenibacillus alvei K165 confers inherited resistance to Verticillium dahliae. J. Exp. Bot. 2021, 72, 4565–4576. [Google Scholar] [CrossRef] [PubMed]
- Somers, E.; Vanderleyden, J.; Srinivasan, M. Rhizosphere Bacterial Signalling: A Love Parade Beneath Our Feet. Crit. Rev. Microbiol. 2004, 30, 205–240. [Google Scholar] [CrossRef]
- Dennis, P.G.; Miller, T.; Hirsch, P.R. Are root exudates more important than other sources of rhizodeposits in structuring rhizosphere bacterial communities? FEMS Microbiol. Ecol. 2010, 72, 313–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, E.; Carson, W.P. The Ecology and Natural History of Foliar Bacteria with a Focus on Tropical Forests and Agroecosystems. Bot. Rev. 2015, 81, 105–149. [Google Scholar] [CrossRef]
- Lugtenberg, B.J.J.; Kravchenko, L.V.; Simons, M. Tomato seed and root exudate sugars: Composition, utilization by Pseudomonas biocontrol strains and role in rhizosphere colonization. Environ. Microbiol. 1999, 1, 439–446. [Google Scholar] [CrossRef]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Genet. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Lindow, S.E.; Brandl, M.T. Microbiology of the Phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [Green Version]
- Monier, J.-M.; Lindow, S.E. Frequency, Size, and Localization of Bacterial Aggregates on Bean Leaf Surfaces. Appl. Environ. Microbiol. 2004, 70, 346–355. [Google Scholar] [CrossRef] [Green Version]
- Delmotte, N.; Knief, C.; Chaffron, S.; Innerebner, G.; Roschitzki, B.; Schlapbach, R.; von Mering, C.; Vorholt, J.A. Community proteogenomics reveals insights into the physiology of phyllosphere bacteria. Proc. Natl. Acad. Sci. USA 2009, 106, 16428–16433. [Google Scholar] [CrossRef] [Green Version]
- Chen, L. SWEET sugar transporters for phloem transport and pathogen nutrition. New Phytol. 2014, 201, 1150–1155. [Google Scholar] [CrossRef]
- Eom, J.-S.; Chen, L.-Q.; Sosso, D.; Julius, B.T.; Lin, I.; Qu, X.-Q.; Braun, D.M.; Frommer, W.B. SWEETs, transporters for intracellular and intercellular sugar translocation. Curr. Opin. Plant Biol. 2015, 25, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Zhang, Z.; Ren, Z.; Wang, X.; Sun, W.; Feng, H.; Zhao, J.; Zhang, F.; Li, W.; Ma, X.; et al. The GhSWEET42 Glucose Transporter Participates in Verticillium dahliae Infection in Cotton. Front. Plant Sci. 2021, 12, 690754. [Google Scholar] [CrossRef] [PubMed]
- Tjamos, E.C.; Tsitsigiannis, D.I.; Tjamos, S.E.; Antoniou, P.P.; Katinakis, P. Selection and Screening of Endorhizosphere Bacteria from Solarized Soils as Biocontrol Agents Against Verticillium dahliae of Solanaceous Hosts. Eur. J. Plant Pathol. 2004, 110, 35–44. [Google Scholar] [CrossRef]
- Liu, L.; Kloepper, J.W.; Tuzun, S. Induction of Systemic Resistance in Cucumber Against Fusarium Wilt by Plant Growth-Promoting Rhizobacteria. Phytopathology 1995, 85, 695–698. [Google Scholar] [CrossRef]
- Vaneechoutte, M.; Dijkshoorn, L.; Tjernberg, I.; Elaichouni, A.; de Vos, P.; Claeys, G.; Verschraegen, G. Identification of Acinetobacter genomic species by amplified ribosomal DNA restriction analysis. J. Clin. Microbiol. 1995, 33, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Porsby, C.H.; Nielsen, K.F.; Gram, L. Phaeobacter and Ruegeria Species of the Roseobacter Clade Colonize Separate Niches in a Danish Turbot (Scophthalmus maximus)-Rearing Farm and Antagonize Vibrio anguillarum under Different Growth Conditions. Appl. Environ. Microbiol. 2008, 74, 7356–7364. [Google Scholar] [CrossRef] [Green Version]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes De Novo Assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinuesa, P.; Contreras-Moreira, B. Robust Identification of Orthologues and Paralogues for Microbial Pan-Genomics Using GET_HOMOLOGUES: A Case Study of pIncA/C Plasmids. Adv. Struct. Safety Stud. 2015, 1231, 203–232. [Google Scholar] [CrossRef] [Green Version]
- Vinuesa, P.; Ochoa-Sánchez, L.E.; Contreras-Moreira, B. GET_PHYLOMARKERS, a Software Package to Select Optimal Orthologous Clusters for Phylogenomics and Inferring Pan-Genome Phylogenies, Used for a Critical Geno-Taxonomic Revision of the Genus Stenotrophomonas. Front. Microbiol. 2018, 9, 771. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Malandraki, I.; Tjamos, S.E.; Pantelides, I.; Paplomatas, E.J. Thermal inactivation of compost suppressiveness implicates possible biological factors in disease management. Biol. Control 2008, 44, 180–187. [Google Scholar] [CrossRef]
- Bakker, P.A.H.M.; Lamers, J.G.; Bakker, A.W.; Marugg, J.D.; Weisbeek, P.J.; Schippers, B. The role of siderophores in potato tuber yield increase by Pseudomonas putida in a short rotation of potato. Eur. J. Plant Pathol. 1986, 92, 249–256. [Google Scholar] [CrossRef]
- Van Wees, S.; Pieterse, C.; Trijssenaar, A.; Van’t Westende, Y.A.; Hartog, F.; Van Loon, L.C. Differential Induction of Systemic Resistance in Arabidopsis by Biocontrol Bacteria. Mol. Plant-Microbe Interact. 1997, 10, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Bakker, P.A.H.M.; Raaijmakers, J.M.; Schippers, B. Role of iron in the suppression of bacterial plant pathogens by fluorescent pseudomonads. In Iron Chelation in Plants and Soil Microorganisms; Barton, L.L., Hemming, B.C., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 269–278. [Google Scholar]
- Mercado-Blanco, J.; Jurado, R.; Hervás, A.; Jiménez-Dıaz, R.M. Suppression of Verticillium wilt in olive planting stocks by root-associated fluorescent Pseudomonas spp. Biol. Control. 2004, 30, 474–486. [Google Scholar] [CrossRef]
- Raaijmakers, J.; Vlami, M.; De Souza, J.T. Antibiotic production by bacterial biocontrol agents. Antonie Leeuwenhoek 2002, 81, 537–547. [Google Scholar] [CrossRef]
- Validov, S.Z.; Kamilova, F.; Lugtenberg, B.J. Pseudomonas putida strain PCL1760 controls tomato foot and root rot in stonewool under industrial conditions in a certified greenhouse. Biol. Control. 2009, 48, 6–11. [Google Scholar] [CrossRef]
- Sun, D.; Zhuo, T.; Hu, X.; Fan, X.; Zou, H. Identification of a Pseudomonas putida as biocontrol agent for tomato bacterial wilt disease. Biol. Control. 2017, 114, 45–50. [Google Scholar] [CrossRef]
- Pantelides, I.; Tjamos, S.; Striglis, I.A.; Chatzipavlidis, I.; Paplomatas, E.J. Mode of action of a non-pathogenic Fusarium oxysporum strain against Verticillium dahliae using Real Time QPCR analysis and biomarker transformation. Biol. Control. 2009, 50, 30–36. [Google Scholar] [CrossRef]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Kemen, E. Microbe–microbe interactions determine oomycete and fungal host colonization. Curr. Opin. Plant Biol. 2014, 20, 75–81. [Google Scholar] [CrossRef]
- Kwak, Y.-S.; Weller, D.M. Take-all of Wheat and Natural Disease Suppression: A Review. Plant Pathol. J. 2013, 29, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Isaac, I. A comparative study of pathogenic isolates of Verticillium. Trans. Br. Mycol. Soc. 1949, 32, 137–157. [Google Scholar] [CrossRef]
- Malca, I.; Erwin, D.C.; Moje, W.; Jones, B. Effects of pH and carbon and nitrogen sources on the growth of Verticillium alboatrum. Phytopathology 1966, 56, 401–406. [Google Scholar]
- Cooper, R.; Wood, R. Regulation of synthesis of cell wall degrading enzymes by Veticillium alboatrum and Fusarium oxsporum f. sp. lycopersici. Physiol. Plant Pathol. 1975, 5, 135–156. [Google Scholar] [CrossRef]
- Sarmiento-Villamil, J.L.; García-Pedrajas, N.E.; Cañizares, M.C.; García-Pedrajas, M.D. Molecular Mechanisms Controlling the Disease Cycle in the Vascular Pathogen Verticillium dahliae Characterized Through Forward Genetics and Transcriptomics. Mol. Plant-Microbe Interact. 2020, 33, 825–841. [Google Scholar] [CrossRef] [PubMed]
- Naglik, J.; Albrecht, A.; Bader, O.; Hube, B. Candida albicans proteinases and host/pathogen interactions. Cell. Microbiol. 2004, 6, 915–926. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziazia, P.; Poulaki, E.G.; Gkizi, D.; Lozano, L.; Serrano, M.; Tjamos, S.E. Feeding the Microbes: A Strategy to Control Verticillium Wilt. Agronomy 2021, 11, 1946. https://doi.org/10.3390/agronomy11101946
Ziazia P, Poulaki EG, Gkizi D, Lozano L, Serrano M, Tjamos SE. Feeding the Microbes: A Strategy to Control Verticillium Wilt. Agronomy. 2021; 11(10):1946. https://doi.org/10.3390/agronomy11101946
Chicago/Turabian StyleZiazia, Paraskevi, Eirini G. Poulaki, Danai Gkizi, Luis Lozano, Mario Serrano, and Sotirios E. Tjamos. 2021. "Feeding the Microbes: A Strategy to Control Verticillium Wilt" Agronomy 11, no. 10: 1946. https://doi.org/10.3390/agronomy11101946
APA StyleZiazia, P., Poulaki, E. G., Gkizi, D., Lozano, L., Serrano, M., & Tjamos, S. E. (2021). Feeding the Microbes: A Strategy to Control Verticillium Wilt. Agronomy, 11(10), 1946. https://doi.org/10.3390/agronomy11101946