
Computational Interaction Analysis of Sirex noctilio Odorant-Binding Protein (SnocOBP7) Combined with Female Sex Pheromones and Symbiotic Fungal Volatiles

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. RNA Extraction and cDNA Synthesis
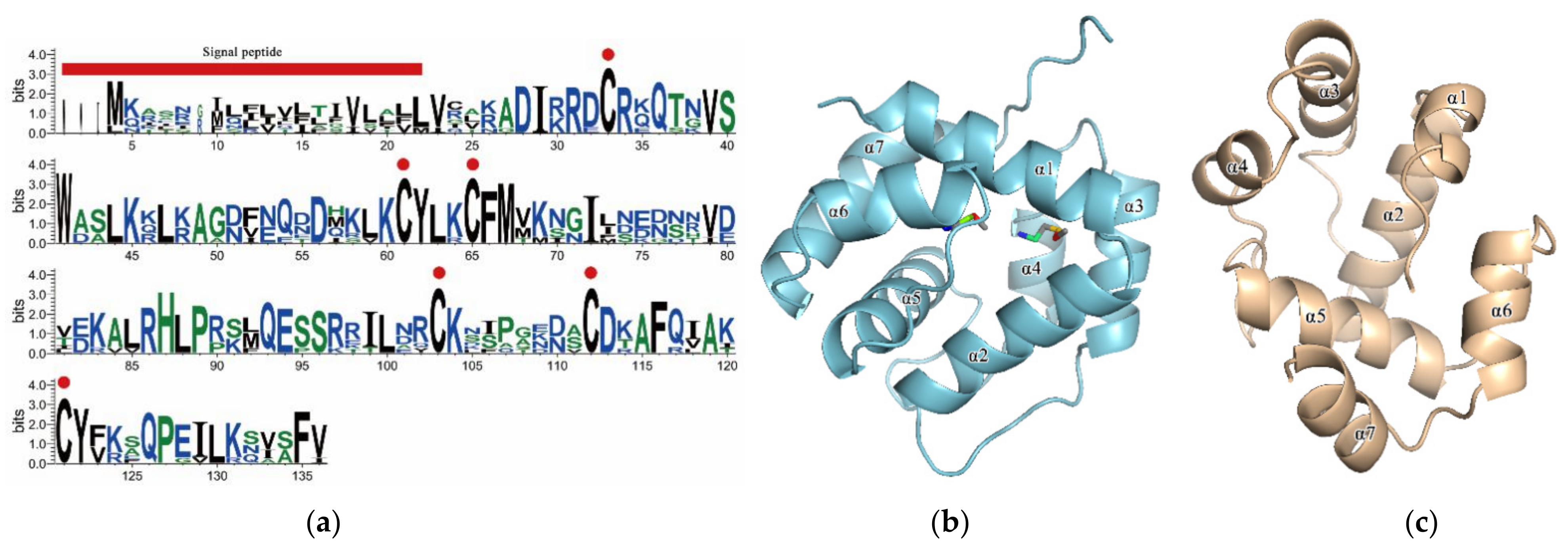
2.2. Sequence Analysis of SnocOBP7
2.3. Gene Cloning and Verification of the SnocOBP7 Sequence
2.4. Homology Modeling and Molecular Docking
2.5. Molecular Dynamics (MD) Simulations of SnocOBP7 and 11 Ligands
2.6. Binding Free Energy Calculation and Per-Residue Free Energy Decomposition
2.7. Computational Alanine Scanning (CAS)
3. Results
3.1. Sequence Analysis and Homology Modeling
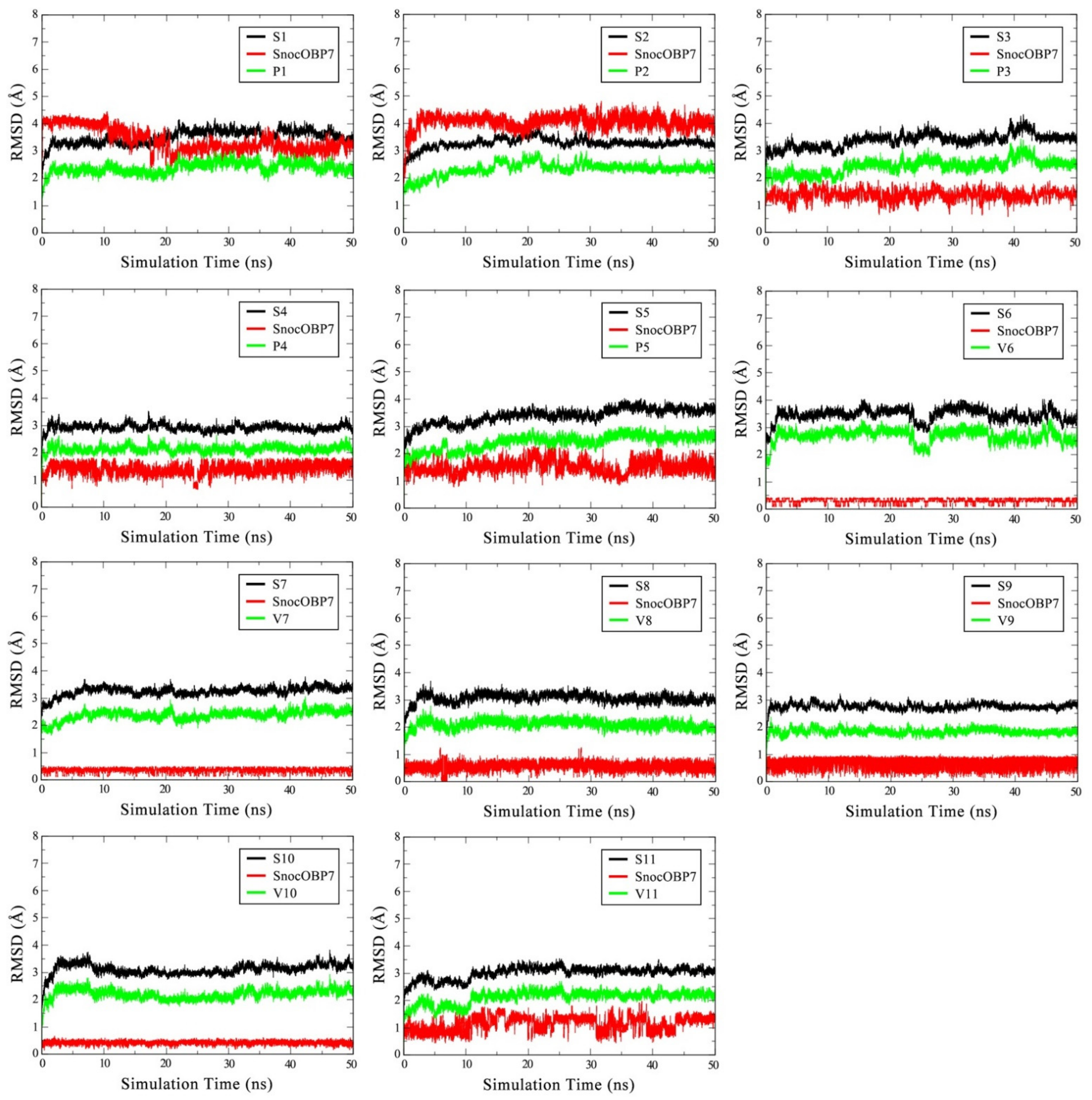
3.2. Stability of SnocOBP7–Ligand Complexes in MD Simulation
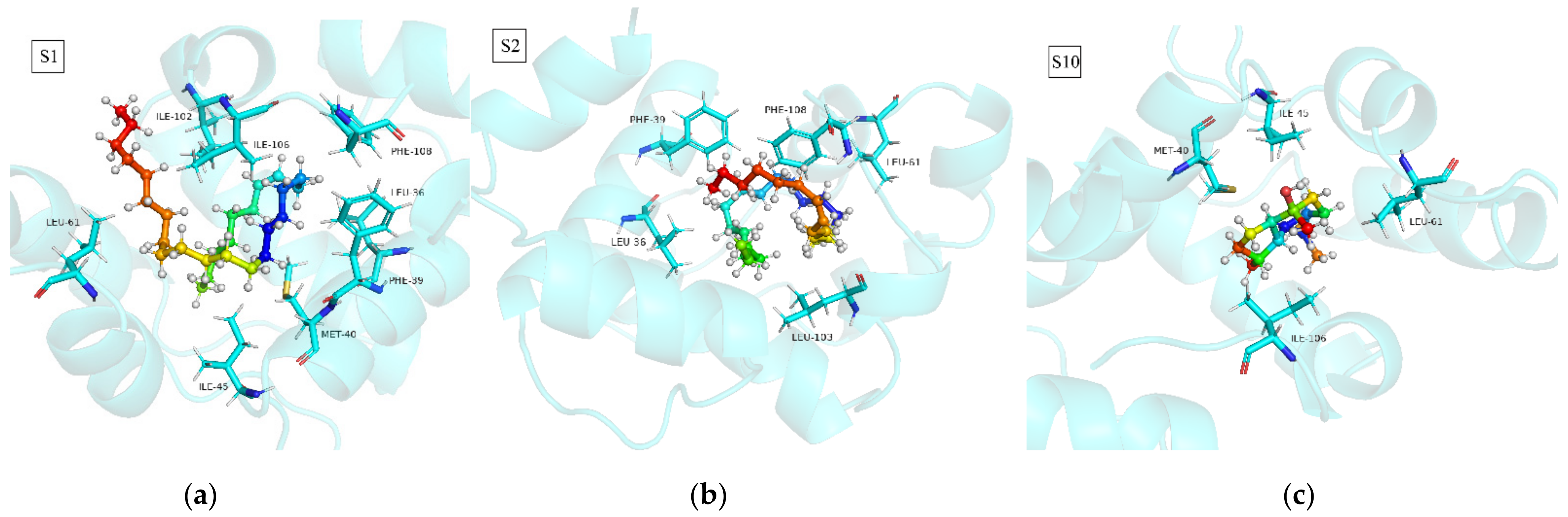
3.3. Binding Mode Analysis of SnocOBP7-Ligand Complexes
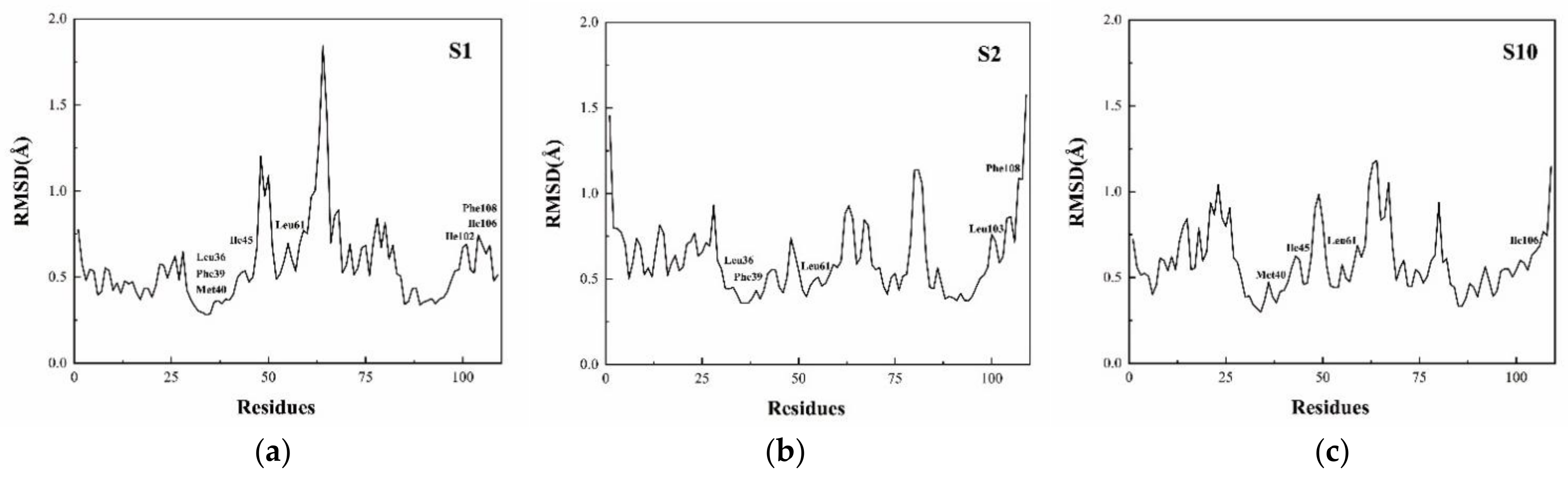
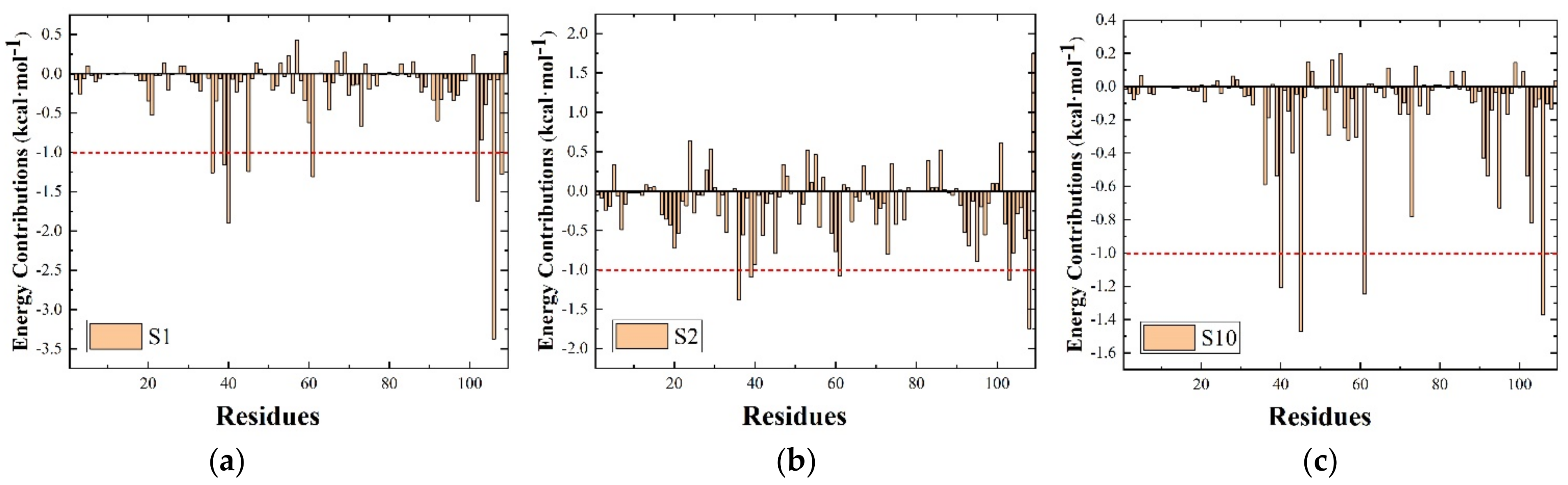
3.4. Per-Residue Free Energy Decomposition
3.5. Computational Alanine Scanning (CAS)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leal, W.S. Odorant Reception in Insects: Roles of Receptors, Binding Proteins, and Degrading Enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Zhou, J.J.; Ban, L.P.; Calvello, M. Soluble proteins in insect chemical communication. Cell. Mol. Life Sci. CMLS 2006, 63, 1658–1676. [Google Scholar] [CrossRef] [PubMed]
- Krieger, J.; Breer, H. Transduction mechanisms of olfactory sensory neurons-20. Insect Pheromone Biochem. Mol. Biol. 2003, 593–607. [Google Scholar]
- Mombaerts, P. Seven-Transmembrane Proteins as Odorant and Chemosensory Receptors. Science 1999, 286, 707–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.; Faisal, Y.; Claudia, S.; Nicolas, D.; Coppin, C.W.; Gunjan, P.; Oakeshott, J.G.; Martine, M. An antennal carboxylesterase from Drosophila melanogaster, esterase 6, is a candidate odorant-degrading enzyme toward food odorants. Front. Physiol. 2015, 6, 315. [Google Scholar]
- Vogt, R.G.; Riddiford, L.M. Pheromone binding and inactivation by moth antennae. Nature 1981, 293, 161–163. [Google Scholar] [CrossRef]
- Briand, L.; Nespoulous, C.; Huet, J.C.; Pernollet, J.C. Disulfide pairing and secondary structure of ASP1, an olfactory-binding protein from honeybee (Apis mellifera L.). J. Pept. Res. 2010, 58, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Maida, R. Odorant-binding proteins in insects. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1995, 111, 503. [Google Scholar] [CrossRef]
- Bino, J.; Andrej, S. Comparative protein structure modeling by iterative alignment, model building and model assessment. Nucleic Acids Res. 2003, 31, 3982–3992. [Google Scholar]
- Ai, H.; Liu, Y.; Long, G.; Yuan, Y.; Chen, Y. Functional characteristics of a novel odorant binding protein in the legume pod borer, Maruca vitrata. Sci. Rep. 2021, 11, 14027. [Google Scholar] [CrossRef]
- Li, D.; Li, C.; Liu, D. Analyses of structural dynamics revealed flexible binding mechanism for the Agrilus mali odorant binding protein 8 towards plant volatiles. Pest Manag. Sci. 2020, 77, 1642–1653. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Zhang, X.C.; Zhu, R.; Yao, W.C.; Wu, X.M. Computational and Experimental Approaches to Decipher the Binding Mechanism of General Odorant-Binding Protein 2 from Athetis lepigone to Chlorpyrifos and Phoxim. J. Agric. Food Chem. 2020, 69, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Bookwalter, J.D.; Riggins, J.J.; Dean, J.F.D.; Mastro, V.C.; Gandhi, K.J.K. Colonization and Development of Sirex noctilio (Hymenoptera: Siricidae) in Bolts of a Native Pine Host and Six Species of Pine Grown in the Southeastern United States. J. Entomol. Sci. 2019, 54, 1–18. [Google Scholar] [CrossRef]
- Chrystal, R.N. The Sirex Wood-wasps and their Importance in Forestry. Bull. Entomol. Res. 1928, 19, 219. [Google Scholar] [CrossRef]
- Martinson, S.J.; Ajó, A.F.; Martínez, A.; Krivak-Tetley, F.E.; Villacide, J.M.; Ayres, M.P.; Corley, J.C. Attack rates of Sirex noctilio and patterns of pine tree defenses and mortality in northern Patagonia. Bull. Entomol. Res. 2018, 109, 141–149. [Google Scholar] [CrossRef]
- Zylstra, K.E.; Mastro, V.C. Common mortality factors of woodwasp larvae in three northeastern United States host species. J. Insect Sci. 2012, 12, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayres, M.P.; Sullivan, J.M.; Harrison, T.; Lombardero, M. Diagnosing the Presence of Sirex noctilio from Examination of Dead and Dying Pine Trees. Available online: http://www.dartmouth.edu/~mpayres/pubs/SirexDiagnostics.pdf (accessed on 29 November 2021).
- Carnegie, A.J.; Eldridge, R.H.; Waterson, D.G. History and Management of Sirex Wood Wasp in Pine Plantations in New South Wales, Australia. N. Z. J. For. Sci. 2005, 35, 3–24. [Google Scholar]
- Da-Peng, L.I.; Shi, J.; Luo, Y.Q. Mutualism between the Eurasian woodwasp, Sirex noctilio (Hymenoptera: Siricidae) and its fungal symbiont Amylostereum areolatum (Russulales: Amylostereaceae). Acta Entomol. Sin. 2015, 58, 1019–1029. [Google Scholar]
- Farji-Brener, A.G.; Corley, J.C. Successful invasions of hymenopteran insects into NW Patagonia. Ecol. Austral 1998, 8, 237–249. [Google Scholar]
- Madden, J.L. Sirex in Australasia. In Dynamics of Forest Insect Populations; Springer: Boston, MA, USA, 1988; pp. 407–429. [Google Scholar]
- Myers, S.W.; Zylstra, K.E.; Francese, J.A.; Borchert, D.M. Phenology and flight periodicity of Sirex noctilio (Hymenoptera: Siricidae) in central New York, U.S.A. Agric. For. Entomol. 2013, 16, 129–135. [Google Scholar] [CrossRef]
- Tribe, G.D. The woodwasp Sirex noctilio Fabricius (Hymenoptera: Siricidae), a pest of Pinus species, now established in South Africa: Short communication. Afr. Entomol. 1995, 3, 215–217. [Google Scholar]
- Sun, X.; Xu, Q.; Luo, Y. A Maximum Entropy Model Predicts the Potential Geographic Distribution of Sirex noctilio. Forests 2020, 11, 175. [Google Scholar] [CrossRef] [Green Version]
- Hurley, B.P.; Slippers, B.; Wingfield, M.J. A comparison of control results for the alien invasive woodwasp, Sirex noctilio, in the southern hemisphere. Agric. For. Entomol. 2010, 9, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Carnegiea, A.J.; Matsukib, M.; Haugenc, D.; Hurley, D.B.P.; Ahumadae, R. Predicting the potential distribution of Sirex noctilio (Hymenoptera: Siricidae), a significant exotic pest of Pinus plantations. Ann. For. Sci. 2006, 63, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Mlonyeni, X.O.; Wingfield, B.D.; Greeff, J.M.; Hurley, B.P.; Slippers, B. Population variation in traits of Deladenus siricidicola that could influence the biocontrol of Sirex noctilio in South Africa. Int. J. Pest Manag. 2018, 64, 324–332. [Google Scholar] [CrossRef] [Green Version]
- Rui, L. Identification of Pheromone Components of Sirex noctilio Fabricius and Trapping Technology in Forest; Beijing Forestry University: Beijing, China, 2019. [Google Scholar]
- Fazila, Y.; Gurr, G.M.; Carnegie, A.J.; Bedding, R.A.; Richard, B.; Gitau, C.W.; Nicol, H.I. The bark beetle, Ips grandicollis, disrupts biological control of the woodwasp, Sirex noctilio, via fungal symbiont interactions. FEMS Microbiol. Ecol. 2014, 88, 38–47. [Google Scholar]
- Böröczky, K.; Zylstra, K.E.; Mccartney, N.B.; Mastro, V.C.; Tumlinson, J.H. Volatile Profile Differences and the Associated Sirex noctilio Activity in Two Host Tree Species in the Northeastern United States. J. Chem. Ecol. 2012, 38, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Simpson, R.F.; Mcquilkin, R.M. Identification of volatiles from felled Pinus radiata and the electroantennograms they elicit from Sirex noctilio. Entomol. Exp. Appl. 2011, 19, 205–213. [Google Scholar] [CrossRef]
- Sarvary, M.A.; Cooperband, M.F.; Hajek, A.E. The importance of olfactory and visual cues in developing better monitoring tools for Sirex noctilio (Hymenoptera: Siricidae). Agric. For. Entomol. 2014, 17, 29–35. [Google Scholar] [CrossRef]
- Katalin, B.; Damon, J.C.; Tappey, H.J.; Joshua, C.K.; Kelley, E.Z.; Victor, C.M.; James, H.T. Monoalkenes as Contact Sex Pheromone Components of the Woodwasp Sirex noctilio. J. Chem. Ecol. 2009, 35, 1202–1211. [Google Scholar]
- Taylor, K.L. The Sirex woodwasp: Ecology and Control of an Introduced Forest Insect; Csiro: Melbourne, Australia, 1981; pp. 231–248. [Google Scholar]
- Guo, B.; Hao, E.; Qiao, H.; Wang, J.; Lu, P. Antennal transcriptome analysis of olfactory genes and characterizations of odorant binding proteins in two woodwasps, Sirex noctilio and Sirex nitobei (Hymenoptera: Siricidae). BMC Genom. 2021, 22, 172. [Google Scholar] [CrossRef]
- Alder, B.J.; Wainwright, T.E. Studies in Molecular Dynamics. I. General Method. J. Chem. Phys. 1959, 31, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Russell, R.B.; Alber, F.; Aloy, P.; Davis, F.P.; Korkin, D.; Pichaud, M.; Topf, M.; Sali, A. A structural perspective on protein–protein interactions. Curr. Opin. Struct. Biol. 2004, 14, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; Mcwilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Schwede, T.; Kopp, J.; Guex, N.; Peitsch, M.C. SWISS-MODEL: An automated protein homology-modeling server. Nucleic Acids Res. 2003, 31, 3381–3385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiser, A.; Do, R.K.G. Modeling of loops in protein structures. Protein Sci. 2010, 9, 1753–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Blundell, T.L. Comparative Protein Modelling by Satisfaction of Spatial Restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar]
- Martí-Renom, M.A.; Stuart, A.C.; Fiser, A.; Sánchez, R.; Sali, A. Comparative Protein Structure Modeling of Genes and Genomes. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 291–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Lasker, K.; Schneidman-Duhovny, D.; Webb, B.; Huang, C.C.; Pettersen, E.F.; Goddard, T.D.; Meng, E.C.; Sali, A.; Ferrin, T.E. UCSF Chimera, MODELLER, and IMP: An integrated modeling system. J. Struct. Biol. 2012, 179, 269–278. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Llc, S. The PyMOL Molecular Graphics System, Version 1.3r1. Proteins Struct. Funct. Bioinform. 2010, 30, 442–454. [Google Scholar]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers—ScienceDirect. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Yuan, X.; Li, J.; Lin, S.; Yang, B.; Chen, C.; Zhao, J.; Zheng, W.; Liao, H.; Yang, Z.; et al. Assessing the performance of the g_mmpbsa tools to simulate the inhibition of oseltamivir to influenza virus neuraminidase by molecular mechanics Poisson–Boltzmann surface area methods. J. Chin. Chem. Soc. 2020, 67, 46–53. [Google Scholar] [CrossRef]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and Testing of a General AMBER Force Field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Vranken, W.F. ACPYPE-Antechamber python parser interface. BMC Res. Notes 2012, 5, 367. [Google Scholar]
- Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Berendsen, H. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Kumar, R.; Lynn, A. G_mmpbsa—A GROMACS tool for high-throughput MM-PBSA calculations. J. Chem. Inf. Model. 2014, 54, 1951–1962. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wu, F.; Yuan, X.; Zhang, L.; Zhang, S. Novel binding patterns between ganoderic acids and neuraminidase: Insights from Docking, Molecular dynamics and MM/PBSA studies. J. Mol. Graph. Model. 2016, 65, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Li, Y.; Xing, Y.; Li, R.; Liu, J. Structural insights into two representative conformations of the complex formed by Grapholita molesta (Busck) pheromone binding protein 2 and Z-8-dodecenyl acetate. J. Agric. Food Chem. 2019, 67, 4425–4434. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.F.; Wang, F.; Chen, Y.Z.; Hao, G.F.; Yang, G.F. LARMD: Integration of bioinformatic resources to profile ligand-driven protein dynamics with a case on the activation of estrogen receptor. Brief. Bioinform. 2019, 21, 6. [Google Scholar]
- Navriti, C.; Om, S. Active site fingerprinting and pharmacophore screening strategies for the identification of dual inhibitors of protein kinase C (ΡΚCβ) and poly (ADP-ribose) polymerase-1 (PARP-1). Mol. Divers. 2016, 20, 747–761. [Google Scholar]
- Zheng, J.; Li, J.; Han, L.; Wang, Y.; Wu, W.; Qi, X.; Tao, Y.; Zhang, L.; Zhang, Z.; Chen, Z. Crystal structure of the Locusta migratoria odorant binding protein. Biochem. Biophys. Res. Commun. 2015, 456, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Broto, C.; Nita, P. NAPS: Network Analysis of Protein Structures. Nucleic Acids Res. 2016, 44, W375–W382. [Google Scholar]
- Zhang, F.; Chen, N.; Wu, R. Molecular Dynamics Simulations Elucidate Conformational Dynamics Responsible for the Cyclization Reaction in TEAS. J. Chem. Inf. Model. 2016, 56, 877. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Ren, J.; Liu, T.; Lin, S.; Liao, H. Molecular Docking and Molecular Dynamics (MD) Simulation of Human Anti Complement Factor H (CFH) Antibody Ab42 and CFH Polypeptide (pCFH). Int. J. Mol. Sci. 2018, 20, 2568. [Google Scholar] [CrossRef]
- Kortemme, T.; Kim, D.E.; Baker, D. Computational Alanine Scanning of Protein-Protein Interfaces. Sci. Signal. 2004, 2004, pl2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spassov, V.Z.; Yan, L. pH-selective mutagenesis of protein-protein interfaces: In silico design of therapeutic antibodies with prolonged half-life. Proteins Struct. Funct. Genet. 2013, 81, 704–714. [Google Scholar] [CrossRef] [Green Version]
- Spassov, V.Z.; Yan, L. A pH-dependent computational approach to the effect of mutations on protein stability. J. Comput. Chem. 2016, 37, 2573–2587. [Google Scholar] [CrossRef] [PubMed]
- Yipeng, L.; Lixiao, D.; Yun, Z.; Shiyong, Y.; Qiong, Z.; Guirong, W.; Yang, L. Identification and sex-biased profiles of candidate olfactory genes in the antennal transcriptome of the parasitoid wasp Cotesia vestalis. Comp. Biochem. Physiol. D Genom. Proteom. 2020, 34, 100657. [Google Scholar]
- Zhou, C.-X.; Min, S.-F.; Yan-Long, T.; Wang, M.-Q. Analysis of antennal transcriptome and odorant binding protein expression profiles of the recently identified parasitoid wasp, Sclerodermus sp. Comp. Biochem. Physiol. Part D Genom. Proteom. 2015, 16, 10–19. [Google Scholar] [CrossRef]
- Simpson, R.F. Bioassay of pine oil components as attractants for Sirex noctilio (Hymenoptera: Siricidae) using electroantennogram techniques. Entomol. Exp. Appl. 2011, 19, 11–18. [Google Scholar] [CrossRef]
- Morgan, F.; Stewart, N.C. The biology and behaviour of the woodwasp Sirex noctilio F. in New Zealand. Trans. Roy. Soc. N. Z. 1966, 7, 195–204. [Google Scholar]
- Bao, M.; Qiao, H.; Shi, J.; Luo, Y.; Lu, P. Research progress in reproductive behavior and chemical ecological regulation of the European woodwasp (Sirex noctilio), a severe invasive pest. Sci. Silvae Sin. 2020, 56, 127–141. [Google Scholar]
- Buchanan, P.K. Aphyllophorales in Australasia. Aust. Syst. Bot. 2001, 14, 417–437. [Google Scholar] [CrossRef]
- Wang, L.X.; Ren, L.L.; Liu, X.B.; Shi, J.; Wang, J.Z.; Luo, Y.Q. Effects of endophytic fungi in Mongolian pine on the selection behavior of woodwasp (Sirex noctilio) and the growth of its fungal symbiont. Pest Manag. Sci. 2018, 75, 492–505. [Google Scholar] [CrossRef]
- Parkin, E.A. Symbiosis in Larval Siricidæ (Hymenoptera). Nature 1941, 147, 329. [Google Scholar] [CrossRef]
- Nest, M.A.; Wingfield, B.D.; Wingfield, M.J.; Stenlid, J.; Slippers, B. Genetics of Amylostereum Species Associated with Siricidae Woodwasps. In The Sirex Woodwasp and Its Fungal Symbiont; Springer: Dordrecht, The Netherlands, 2012; pp. 81–94. [Google Scholar]
- Madden, J.L. Avian predation of the woodwasp, Sirex noctilio F., and its parasitoid complex in Tasmania. Wildl. Res. 1982, 9, 135–144. [Google Scholar] [CrossRef]
- Yang, B.; Lin, S.-J.; Ren, J.-Y.; Liu, T.; Wang, Y.-M.; Li, C.-M.; Xu, W.-W.; He, Y.-W.; Zheng, W.-H.; Zhao, J.; et al. Molecular Docking and Molecular Dynamics (MD) Simulation of Human Anti-Complement Factor H (CFH) Antibody Ab42 and CFH Polypeptide. Int. J. Mol. Sci. 2019, 20, 2568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.H.; Zhang, X.Q.; Zhang, X.C.; Xu, J.W.; Li, L.L.; Zhu, X.Y.; Wang, J.J.; Wei, J.Y.; Mang, D.Z.; Zhang, F.; et al. Key Amino Acid Residues Influencing Binding Affinities of Pheromone-Binding Protein from Athetis lepigone to Two Sex Pheromones. J. Agric. Food Chem. 2020, 68, 6092–6103. [Google Scholar] [CrossRef] [PubMed]
- Zhen, T.; Yue, L.; Tong, Z.; Xuan, Y.; Ruichi, L.; Jiyuan, L. Structure dynamics reveal key residues essential for the sense of 1-dodecanol by Cydia pomonella pheromone binding protein 2 (CpomPBP2). Pest Manag. Sci. 2020, 76, 3667–3675. [Google Scholar]
- Affonso, R.; Guimaraes, A.P.; Oliveira, A.A.; Slana, G.; Franca, T. Applications of molecular modeling in the design of new insect repellents targeting the odorant binding protein of Anopheles gambiae. J Brazil Chem. Soc. 2013, 24, 473–482. [Google Scholar] [CrossRef]
- Li, H.-L.; Song, X.-M.; Wu, F.; Qiu, Y.-L.; Fu, X.-B.; Zhang, L.-Y.; Tan, J. Chemical structure of semiochemicals and key binding sites together determine the olfactory functional modes of odorant-binding protein 2 in Eastern honey bee, Apis cerana. Int. J. Biol. Macromol. 2020, 145, 876–884. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence |
|---|---|
| SnocOBP7F (5′ to 3′) | GGCGGACATTAGAAGAGACTGT |
| SnocOBP7R (3′ to 5′) | TGCTTCAAGATCTCGGGCTG |
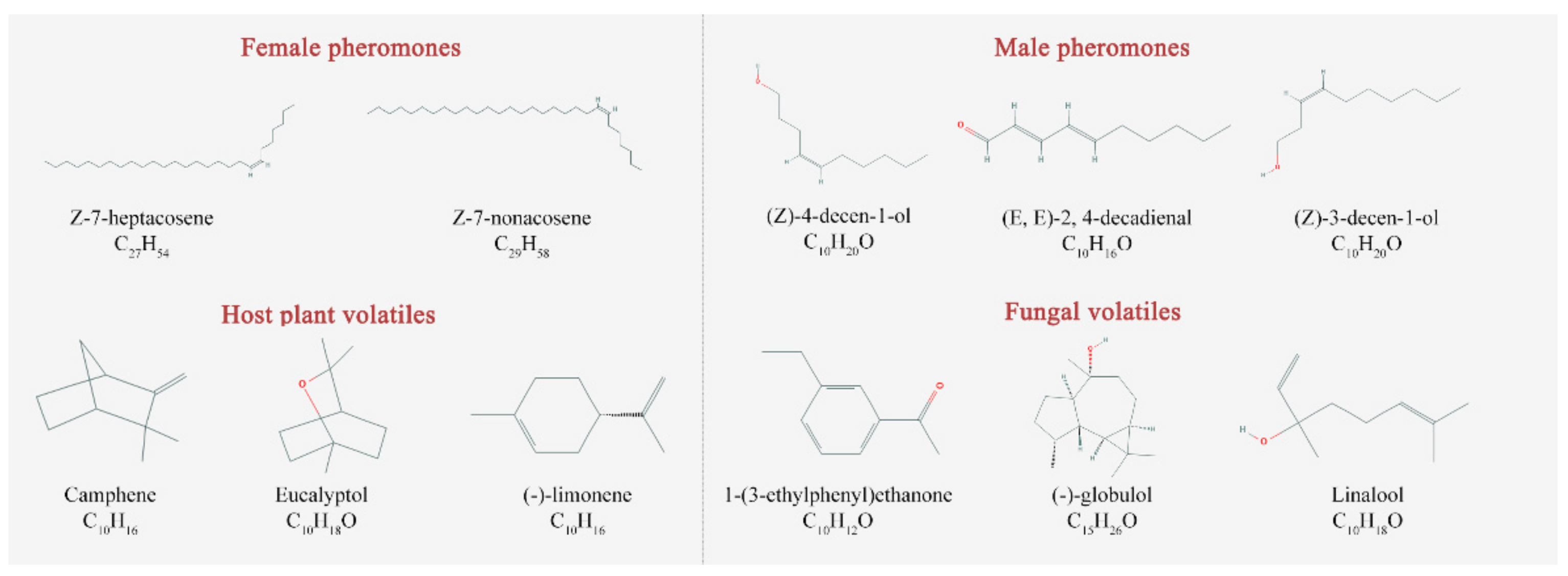
| ID | Ligand Name | Molecular Formula | PubChem ID | Molecular Weight (g/mol) | Complex |
|---|---|---|---|---|---|
| Female pheromones | |||||
| P1 A | (Z)-7-heptacosene | C27H54 | 56936088 | 378.7 | S1 C |
| P2 | (Z)-7-nonacosene | C29H58 | 56936089 | 406.8 | S2 |
| Male pheromones | |||||
| P3 | (Z)-4-decen-1-ol | C10H20O | 5362798 | 156.26 | S3 |
| P4 | (E,E)-2,4-decadienal | C10H16O | 5283349 | 152.23 | S4 |
| P5 | (Z)-3-decen-1-ol | C10H20O | 5352846 | 156.26 | S5 |
| Host plant volatiles | |||||
| V6 B | Camphene | C10H16 | 6616 | 136.23 | S6 |
| V7 | Eucalyptol | C10H18O | 2758 | 154.25 | S7 |
| V8 | (−)-Limonene | C10H16 | 439250 | 136.23 | S8 |
| Fungal volatiles | |||||
| V9 | 1-(3-ethylphenyl)ethanone | C10H12O | 31493 | 148.2 | S9 |
| V10 | (−)-Globulol | C15H26O | 12304985 | 222.37 | S10 |
| V11 | Linalool | C10H18O | 6549 | 154.25 | S11 |
| Complex | Cluster (ns) | Van der Waals Energy | Electrostatic Energy | Polar Solvation Energy | SASA Energy | Binding Energy | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (ΔGvdw) | (ΔGele) | (ΔGPB) | (ΔGSA) | (ΔGbind) | ||||||||||||
| S1 | 21–50 | −59.900 | ± | 0.341 | −0.300 | ± | 0.018 | 10.975 | ± | 0.188 | −6.432 | ± | 0.034 | −55.656 | ± | 0.351 |
| S2 | 23–50 | −62.877 | ± | 0.220 | −0.385 | ± | 0.027 | 13.013 | ± | 0.196 | −6.533 | ± | 0.018 | −56.783 | ± | 0.260 |
| S3 | 25–50 | −26.014 | ± | 0.162 | −1.547 | ± | 0.097 | 9.895 | ± | 0.146 | −3.071 | ± | 0.010 | −20.740 | ± | 0.150 |
| S4 | 25–50 | −30.342 | ± | 0.130 | −2.300 | ± | 0.068 | 15.082 | ± | 0.081 | −3.121 | ± | 0.010 | −20.668 | ± | 0.140 |
| S5 | 33–50 | −29.697 | ± | 0.123 | −5.350 | ± | 0.140 | 14.269 | ± | 0.141 | −3.372 | ± | 0.010 | −24.143 | ± | 0.184 |
| S6 | 27–48 | −24.627 | ± | 0.099 | −0.108 | ± | 0.012 | 4.518 | ± | 0.051 | −2.540 | ± | 0.008 | −22.755 | ± | 0.111 |
| S7 | 25–40 | −26.805 | ± | 0.105 | −0.339 | ± | 0.031 | 5.513 | ± | 0.045 | −2.721 | ± | 0.009 | −24.347 | ± | 0.111 |
| S8 | 25–50 | −24.903 | ± | 0.092 | −0.095 | ± | 0.016 | 7.564 | ± | 0.057 | −2.795 | ± | 0.008 | −20.228 | ± | 0.107 |
| S9 | 25–49 | −25.514 | ± | 0.104 | −4.347 | ± | 0.094 | 13.277 | ± | 0.065 | −2.776 | ± | 0.009 | −19.366 | ± | 0.112 |
| S10 | 21–50 | −34.180 | ± | 0.130 | −3.947 | ± | 0.127 | 10.632 | ± | 0.087 | −3.562 | ± | 0.009 | −31.057 | ± | 0.154 |
| S11 | 25–40 | −26.293 | ± | 0.132 | −3.541 | ± | 0.115 | 12.040 | ± | 0.114 | −3.072 | ± | 0.008 | −20.867 | ± | 0.163 |
| Complex | Residue | ΔGMM | ΔGPB | ΔGSA | ΔGbind |
|---|---|---|---|---|---|
| S1 (21–50 ns) | Leu36 | −1.673 | 0.534 | −0.121 | −1.260 |
| Phe39 | −1.427 | 0.374 | −0.106 | −1.161 | |
| Met40 | −2.041 | 0.302 | −0.159 | −1.898 | |
| Ile45 | −1.173 | 0.037 | −0.108 | −1.244 | |
| Leu61 | −1.183 | 0.058 | −0.182 | −1.309 | |
| Ile102 | −1.816 | 0.344 | −0.156 | −1.626 | |
| Ile106 | −3.076 | 0.137 | −0.435 | −3.374 | |
| Phe108 | −1.561 | 0.385 | −0.098 | −1.275 | |
| S2 (23–50 ns) | Leu36 | −1.535 | 0.313 | −0.160 | −1.382 |
| Phe39 | −1.832 | 0.893 | −0.153 | −1.092 | |
| Leu61 | −0.949 | −0.068 | −0.064 | −1.081 | |
| Leu103 | −2.020 | 1.116 | −0.226 | −1.131 | |
| Phe108 | −3.534 | 2.188 | −0.395 | −1.744 | |
| S10 (21–50 ns) | Met40 | −1.690 | 0.612 | −0.129 | −1.208 |
| Ile45 | −1.455 | 0.078 | −0.094 | −1.470 | |
| Leu61 | −1.121 | −0.007 | −0.115 | −1.243 | |
| Ile106 | −1.218 | 0.020 | −0.173 | −1.371 |
| Complex | Mutation | ΔΔGmut |
|---|---|---|
| S1 | Leu36>Ala | 0.77 |
| Phe39>Ala | 0.46 | |
| Met40>Ala | 1.6 | |
| Ile45>Ala | 0.32 | |
| Leu61>Ala | 1.26 | |
| Ile102>Ala | 0.24 | |
| Ile106>Ala | 0.38 | |
| Phe108>Ala | 0.85 | |
| S2 | Leu36>Ala | 0.88 |
| Phe39>Ala | 1.55 | |
| Leu61>Ala | 1.55 | |
| Leu103>Ala | 1.75 | |
| Phe108>Ala | 2.63 | |
| S10 | Met40>Ala | 0.75 |
| Ile45>Ala | 0.47 | |
| Leu61>Ala | 0.84 | |
| Ile106>Ala | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.-N.; Hao, E.-H.; Li, H.; Yuan, X.-H.; Lu, P.-F.; Qiao, H.-L. Computational Interaction Analysis of Sirex noctilio Odorant-Binding Protein (SnocOBP7) Combined with Female Sex Pheromones and Symbiotic Fungal Volatiles. Agronomy 2021, 11, 2461. https://doi.org/10.3390/agronomy11122461
Li Y-N, Hao E-H, Li H, Yuan X-H, Lu P-F, Qiao H-L. Computational Interaction Analysis of Sirex noctilio Odorant-Binding Protein (SnocOBP7) Combined with Female Sex Pheromones and Symbiotic Fungal Volatiles. Agronomy. 2021; 11(12):2461. https://doi.org/10.3390/agronomy11122461
Chicago/Turabian StyleLi, Yi-Ni, En-Hua Hao, Han Li, Xiao-Hui Yuan, Peng-Fei Lu, and Hai-Li Qiao. 2021. "Computational Interaction Analysis of Sirex noctilio Odorant-Binding Protein (SnocOBP7) Combined with Female Sex Pheromones and Symbiotic Fungal Volatiles" Agronomy 11, no. 12: 2461. https://doi.org/10.3390/agronomy11122461
APA StyleLi, Y. -N., Hao, E. -H., Li, H., Yuan, X. -H., Lu, P. -F., & Qiao, H. -L. (2021). Computational Interaction Analysis of Sirex noctilio Odorant-Binding Protein (SnocOBP7) Combined with Female Sex Pheromones and Symbiotic Fungal Volatiles. Agronomy, 11(12), 2461. https://doi.org/10.3390/agronomy11122461