
The Impact of Forest Fungi on Promoting Growth and Development of Brassica napus L.

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Inoculum from Fungi
2.2. In Vitro Experiment
2.3. Pot Experiment
2.4. Analysis of Microorganism Abundance in Soil
2.5. Analysis of IAA Production by Fungi
2.6. Isolation of Total RNA and Reverse Transcription Reaction
2.7. Semi-Quantitative RT-PCR (sqRT-PCR)
2.8. Bioinformatic Analysis of B. napus Metallothionein Gene Promoters
2.9. Statistical Analysis
3. Results
3.1. Analysis of the Effect of Ectomycorrhizal Fungi on the Growth of Rapeseed Seedlings In Vitro
3.2. Analysis of the Effect of Fungi on the Growth and Development of Rapeseed Shoots in a Pot Experiment
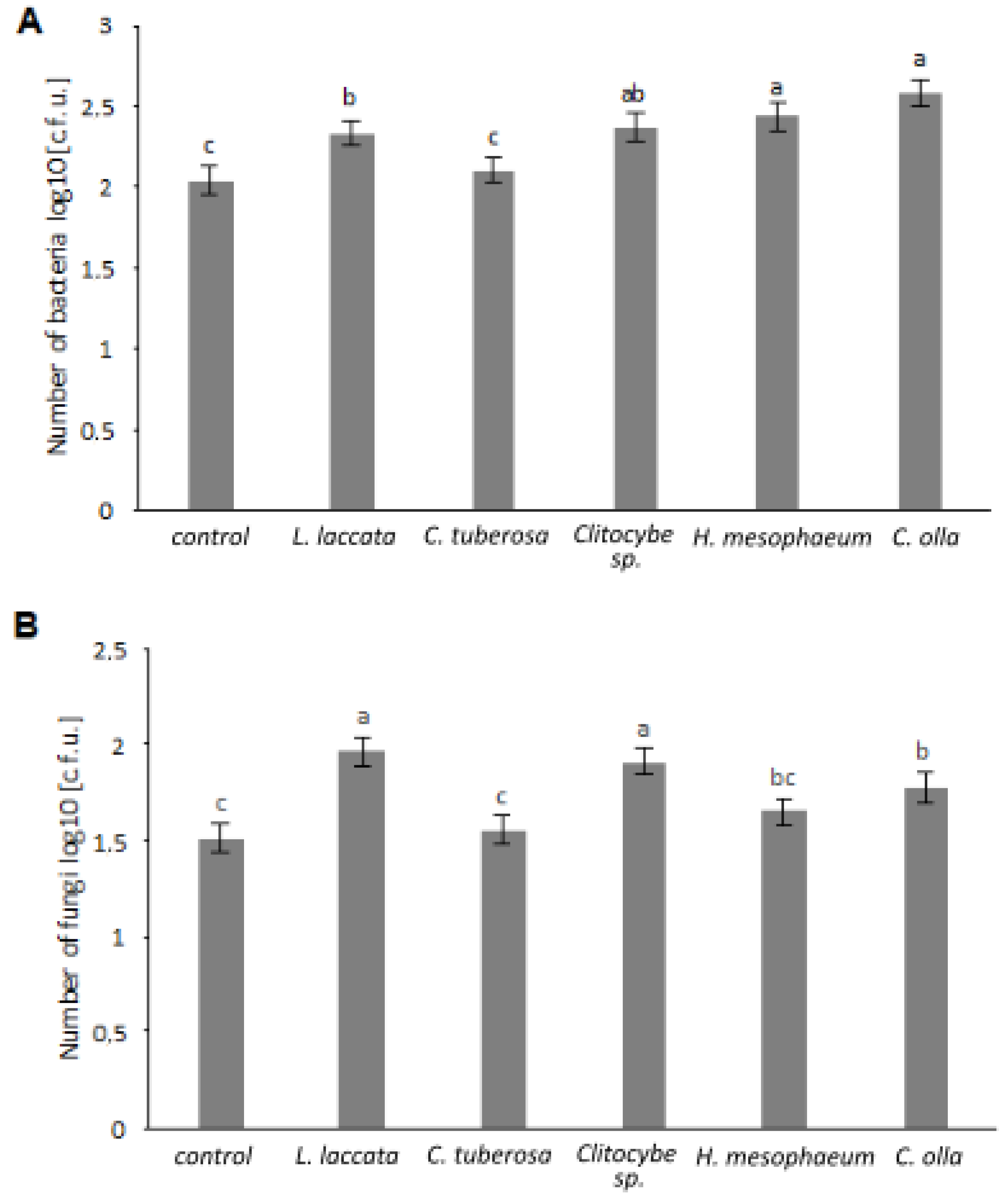
3.3. Analysis of the Microorganisms Populations in the Soil after the Pot Experiment
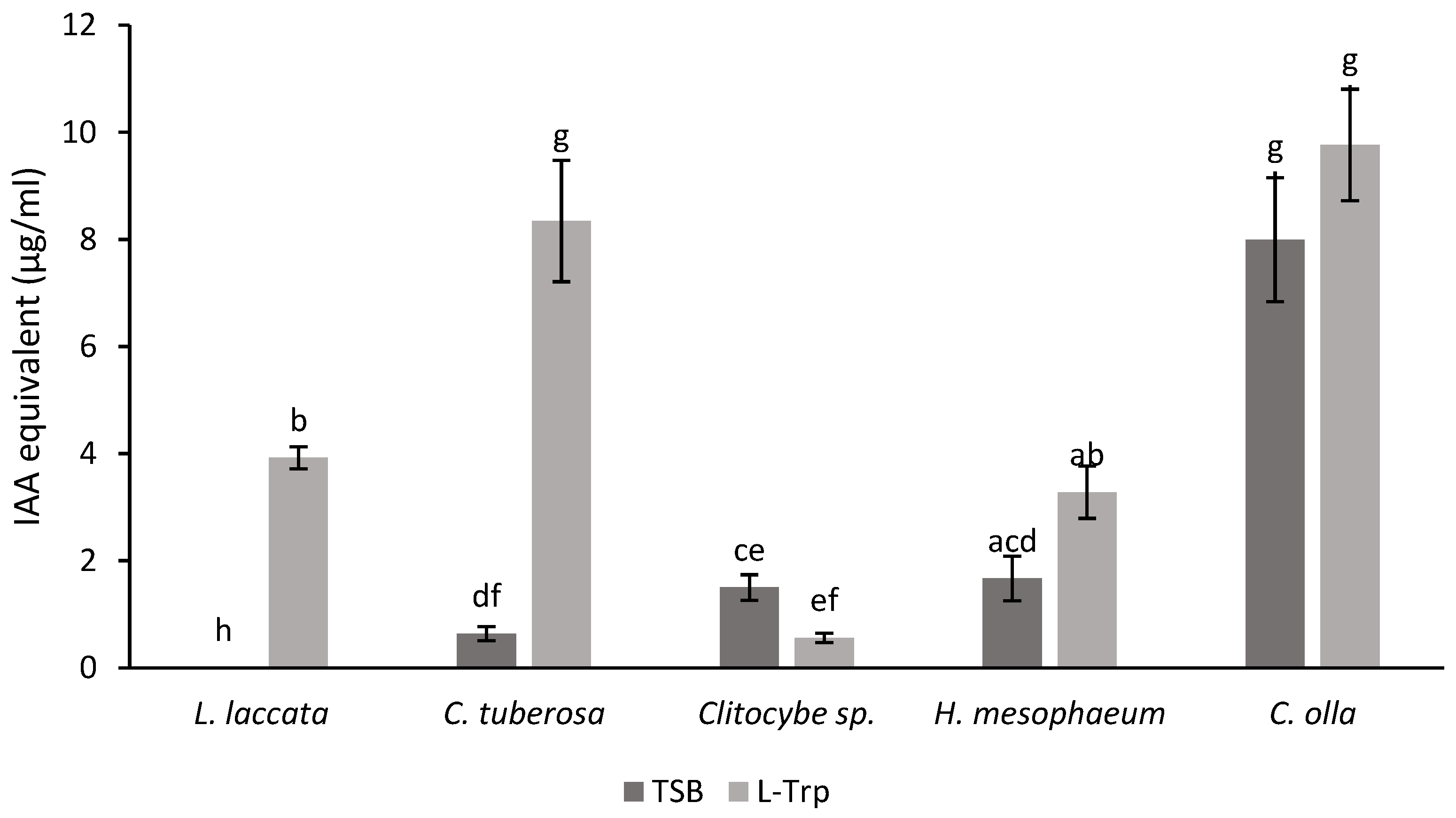
3.4. Analysis of IAA Production by Fungi
3.5. In Silico Analysis of BnMT1-BnMT3 Sequences and Expression of These Genes in the Presence of Ectomycorrhizal Fungi
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Korelewski, K. Review and systematics of works increasing utility value of non-urban lands. Przegl. Nauk. Inz. Ksztalt. Sr. 2006, 2, 27–34. [Google Scholar]
- Baldi, E. Soil–Plant Interaction: Effects on Plant Growth and Soil Biodiversity. Agronomy 2021, 11, 2378. [Google Scholar] [CrossRef]
- Bashan, Y.; Holguin, G.; De-Bashan, L.E. Azospirillum-plant relationships: Physiological, molecular, agricultural, and environmental advances (1997–2003). Can. J. Microbiol. 2004, 50, 521–577. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowska, G.; Zdziechowska, E. The role of rhizobacteria in the stimulation of the growth and development processes and protection of plants against environmental factors. Prog. Plant Prot. 2015, 55, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Hrynkiewicz, K.; Szymańska, S.; Piernik, A.; Thiem, D. Ectomycorrhizal Community Structure of Salix and Betula spp. at a Saline Site in Central Poland in Relation to the Seasons and Soil Parameters. Water Air Soil Pollut. 2015, 226, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Paredes, A.; Valdés, G.; Nuti, M. Ecosystem Functions of Microbial Consortia in Sustainable Agriculture. Agronomy 2020, 10, 1902. [Google Scholar] [CrossRef]
- Znajewska, Z.; Dąbrowska, G.B.; Narbutt, O. Trichoderma viride strains stimulating the growth and development of winter rapeseed (Brassica napus L.). Prog. Plant Prot. 2018, 58, 264–269. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Cortés-Penagos, C.; López-Bucio, J. Trichoderma virens, a Plant Beneficial Fungus, Enhances Biomass Production and Promotes Lateral Root Growth through an Auxin-Dependent Mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, Y.O.; Kim, S.H.; Lee, J.; Kim, H.; Lee, H.-S.; Kwak, S.-S. Three Brassica rapa metallothionein genes are differentially regulated under various stress conditions. Mol. Biol. Rep. 2011, 39, 2059–2067. [Google Scholar] [CrossRef]
- Bonfante, P.; Genre, A. Mechanisms underlying beneficial plant–fungus interactions in mycorrhizal symbiosis. Nat. Commun. 2010, 1, 48. [Google Scholar] [CrossRef] [Green Version]
- Landeweert, R.; Hoffland, E.; Finlay, R.; Kuyper, T.W.; van Breemen, N. Linking plants to rocks: Ectomycorrhizal fungi mobilize nutrients from minerals. Trends Ecol. Evol. 2001, 16, 248–254. [Google Scholar] [CrossRef]
- Jentschke, G.; Godbold, D.L. Metal toxicity and ectomycorrhizas. Physiol. Plant. 2000, 109, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Wallander, H.; Wickman, T. Biotite and microcline as potassium sources in ectomycorrhizal and non-mycorrhizal Pinus sylvestris seedlings. Mycorrhiza 1999, 9, 25–32. [Google Scholar] [CrossRef]
- Wallander, H. Use of strontium isotopes and foliar K content to estimate weathering of biotite induced by pine seedlings colonised by ectomycorrhizal fungi from two different soils. Plant Soil 2000, 222, 215–229. [Google Scholar] [CrossRef]
- Wallander, H. Uptake of P from apatite by Pinus sylvestris seedlings colonised by different ectomycorrhizal fungi. Plant Soil 2000, 218/2, 249–256. [Google Scholar] [CrossRef]
- Luo, Z.-B.; Janz, D.; Jiang, X.; Göbel, C.; Wildhagen, H.; Tan, Y.; Rennenberg, H.; Feussner, I.; Polle, A. Upgrading Root Physiology for Stress Tolerance by Ectomycorrhizas: Insights from Metabolite and Transcriptional Profiling into Reprogramming for Stress Anticipation. Plant Physiol. 2009, 151, 1902–1917. [Google Scholar] [CrossRef] [Green Version]
- Peay, K.G.; Bruns, T.D. Spore dispersal of basidiomycete fungi at the landscape scale is driven by stochastic and deterministic processes and generates variability in plant–fungal interactions. New Phytol. 2014, 204, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Pontes, M.V.A.; Hainaut, M.; Henrissat, B.; Hilden, K.; Mäkelä, M.; de Vries, R.P. Comparative analysis of basidiomycete transcriptomes reveals a core set of expressed genes encoding plant biomass degrading enzymes. Fungal Genet. Biol. 2017, 112, 40–46. [Google Scholar] [CrossRef]
- Caesar-TonThat, T.-C.; Cochran, V.L. Soil aggregate stabilization by a saprophytic lignin-decomposing basidiomycete fungus I. Microbiological aspects. Biol. Fertil. Soils 2000, 32, 374–380. [Google Scholar] [CrossRef]
- Caesar-Tonthat, T.C.; Cochran, V.L. Role of a Saprophytic Basidiomycete Soil Fungus in Aggregate Stabilization. In Proceedings of the 10th International Soil Conservation Organization Meeeting, West Lafayette, IN, USA, 24–29 May 1999; pp. 575–579. [Google Scholar]
- Freisinger, E. Spectroscopic characterization of a fruit-specific metallothionein: M. acuminata MT3. Inorg. Chim. Acta 2007, 360, 369–380. [Google Scholar] [CrossRef]
- Capdevila, M.; Atrian, S. Metallothionein protein evolution: A miniassay. J. Biol. Inorg. Chem. 2011, 16, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Leszczyszyn, O.I.; Imam, H.T.; Blindauer, C.A. Diversity and distribution of plant metallothioneins: A review of structure, properties and functions. Metallomics 2013, 5, 1146–1169. [Google Scholar] [CrossRef] [PubMed]
- Koszucka, A.M.; Dąbrowska, G. Plant metallothionein. Adv. Biol. Cell 2006, 33, 285–302. [Google Scholar]
- Usha, B.; Venkataraman, G.; Parida, A. Heavy metal and abiotic stress inducible metallothionein isoforms from Prosopis juliflora (SW) D.C. show differences in binding to heavy metals in vitro. Mol. Genet. Genom. 2008, 281, 99–108. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Sáez, C.A.; Moenne, A. Copper-induced activation of TRPs and VDCCs triggers a calcium signature response regulating gene expression in Ectocarpus siliculosus. Peer J. 2018, 6, e4556. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowska, G.; Hrynkiewicz, K.; Trejgell, A. Do Arbuscular Mycorrhizal Fungi Affect Metallothionein MT2 Expression In Brassica Napus L. Roots? Acta Biol. Cracov. Bot. 2012, 54, 34–39. [Google Scholar] [CrossRef]
- Guo, W.; Bundithya, W.; Goldsbrough, P.B. Characterization of the Arabidopsis metallothionein gene family: Tissue-specific expression and induction during senescence and in response to copper. New Phytol. 2003, 159, 369–381. [Google Scholar] [CrossRef]
- Ren, Y.; Liu, Y.; Chen, H.; Li, G.; Zhang, X.; Zhao, J. Type 4 metallothionein genes are involved in regulating Zn ion accumulation in late embryo and in controlling early seedling growth in Arabidopsis. Plant Cell Environ. 2011, 35, 770–789. [Google Scholar] [CrossRef]
- Mierek-Adamska, A.; Kotowicz, K.; Goc, A.; Boniecka, J.; Berdychowska, J.; Dąbrowska, G.B. Potential involvement of rapeseed (Brassica napus L.) metallothioneins in the hydrogen peroxide-induced regulation of seed vigour. J. Agron. Crop. Sci. 2019, 205, 598–607. [Google Scholar] [CrossRef]
- Dabrowska, G.; Mierek-Adamska, A.; Goc, A. Characterisation of Brassica napus L. metallothionein genes (BnMTs) expression in organs and during seed germination. Aust. J. Crop Sci. 2013, 7, 1324–1332. [Google Scholar]
- Glenn, M.G.; Chew, F.S.; Williams, P. Hyphal penetration of Brassica (Cruciferae) roots by a vesicular-arbuscular mycorrizal fungus. New Phytol. 1985, 99, 463–472. [Google Scholar] [CrossRef]
- Tommerup, I. Effect of soil water potential on spore germination by vesicular-arbuscular mycorrhizal fungi. Trans. Br. Mycol. Soc. 1984, 83, 193–202. [Google Scholar] [CrossRef]
- Ishimoto, H.; Fukushi, Y.; Yoshida, T.; Tahara, S. Rhizopus and Fusarium are Selected as Dominant Fungal Genera in Rhizospheres of Brassicaceae. J. Chem. Ecol. 2000, 26, 2387–2399. [Google Scholar] [CrossRef]
- Qiang, X.; Weiss, M.; Kogel, K.H.; Schäfer, P. Piriformospora indica—A mutualistic basidiomycete with an exceptionally large plant host range. Mol. Plant Pathol. 2011, 13, 508–518. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.; Yang, L.; Zhang, L.; Jiang, D.; Chen, W.; Li, G. Diversity and biocontrol potential of endophytic fungi in Brassica napus. Biol. Control. 2014, 72, 98–108. [Google Scholar] [CrossRef]
- Dąbrowska, G.; Baum, C.; Trejgell, A.; Hrynkiewicz, K. Impact of arbuscular mycorrhizal fungi on the growth and expression of gene encoding stress protein—Metallothionein BnMT2 in the non-host crop Brassica napus L. J. Plant Nutr. Soil Sci. 2014, 177, 459–467. [Google Scholar] [CrossRef]
- Dąbrowska, G. Plant Metallothioneins: Putative Functions Identified by Promoter Analysis in silico. Acta Biol. Cracov. Bot. 2012, 54, 109–120. [Google Scholar] [CrossRef]
- Gravel, V.; Antoun, H.; Tweddell, R.J. Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation with Pseudomonas putida or Trichoderma atroviride: Possible role of indole acetic acid (IAA). Soil Biol. Biochem. 2007, 39, 1968–1977. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate–phenol–chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Dąbrowska, G.; Hrynkiewicz, K.; Trejgell, A.; Baum, C. The effect of plant growth-promoting rhizobacteria on the phytoextraction of Cd and Zn by Brassica napus L. Int. J. Phytoremediat. 2017, 19, 597–604. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Ülker, B.; E Somssich, I. WRKY transcription factors: From DNA binding towards biological function. Curr. Opin. Plant Biol. 2004, 7, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Matarasso, N.; Schuster, S.; Avni, A. A Novel Plant Cysteine Protease Has a Dual Function as a Regulator of 1-Aminocyclopropane-1-Carboxylic Acid Synthase Gene Expression. Plant Cell 2005, 17, 1205–1216. [Google Scholar] [CrossRef] [Green Version]
- Chakravarty, P.; Hwang, S.F. Effect of an ectomycorrhizal fungus, Laccaria laccata, on Fusarium damping-off in Pinus banksiana seedlings. For. Pathol. 1991, 21, 97–106. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, X. Effects of several ectomycorrhizal fungi on the chlorophyll content and chlorophyll fluorescence parameters in different pine seedlings. J. Najing For. Univ. 2010, 34, 9–12. [Google Scholar]
- Hrynkiewicz, K.; Dabrowska, G.; Baum, C.; Niedojadło, K.; Leinweber, P. Interactive and Single Effects of Ectomycorrhiza Formation and Bacillus cereus on Metallothionein MT1 Expression and Phytoextraction of Cd and Zn by Willows. Water Air Soil Pollut. 2011, 223, 957–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalski, T. Occurrence and associated symptoms of Sirococcus conigenus on Picea abies. Phytopathologia 2010, 58, 53–61. [Google Scholar]
- Hossain, M.M.; Sultana, F.; Islam, S. Plant Growth-Promoting Fungi (PGPF): Phytostimulation and Induced Systemic Resistance; Springer: Singapore, 2017; Volume 2, ISBN 9789811065934. [Google Scholar]
- Schäfer, P.; Pfiffi, S.; Voll, L.M.; Zajic, D.; Chandler, P.M.; Waller, F.; Scholz, U.; Pons-Kühnemann, J.; Sonnewald, S.; Sonnewald, U.; et al. Manipulation of plant innate immunity and gibberellin as factor of compatibility in the mutualistic association of barley roots with Piriformospora indica. Plant J. 2009, 59, 461–474. [Google Scholar] [CrossRef]
- Sukumar, P.; Maloney, G.S.; Muday, G.K. Localized Induction of the ATP-Binding Cassette B19 Auxin Transporter Enhances Adventitious Root Formation in Arabidopsis. Plant Physiol. 2013, 162, 1392–1405. [Google Scholar] [CrossRef]
- Sirrenberg, A.; Göbel, C.; Grond, S.; Czempinski, N.; Ratzinger, A.; Karlovsky, P.; Santos, P.; Feussner, I.; Pawlowski, K. Piriformospora indica affects plant growth by auxin production. Physiol. Plant. 2007, 131, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Steenhoudt, O.; Vanderleyden, J. Azospirillum, a free-living nitrogen-fixing bacterium closely associated with grasses: Genetic, biochemical and ecological aspects. FEMS Microbiol. Rev. 2000, 24, 487–506. [Google Scholar] [CrossRef] [PubMed]
- Dauch, A.L.; Jabaji, S. Metallothionein and bZIP Transcription Factor Genes from Velvetleaf and Their Differential Expression Following Colletotrichum coccodes Infection. Phytopathology 2006, 96, 1116–1123. [Google Scholar] [CrossRef] [Green Version]
- Miles, T.D.; Day, B.; Schilder, A.C. Identification of differentially expressed genes in a resistant versus a susceptible blueberry cultivar after infection by Colletotrichum acutatum. Mol. Plant Pathol. 2011, 12, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Voiblet, C.; Duplessis, S.; Encelot, N.; Martin, F. Identification of symbiosis-regulated genes in Eucalyptus globulus-Pisolithus tinctorius ectomycorrhiza by differential hybridization of arrayed cDNAs. Plant J. 2001, 25, 181–191. [Google Scholar] [CrossRef]
- Rivera-Becerril, F.; Van Tuinen, D.; Martin-Laurent, F.; Metwally, A.; Dietz, K.-J.; Gianinazzi, S.; Gianinazzi-Pearson, V. Molecular changes in Pisum sativum L. roots during arbuscular mycorrhiza buffering of cadmium stress. Mycorrhiza 2005, 16, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Mortier, F.; Le Tacon, F.; Garbaye, J. Effect of inoculum type and inoculation dose on ectomycorrhizal development, root necrosis and growth of Douglas fir seedlings inoculated with Laccaria laccata in a nursery. Ann. Sci. For. 1988, 45, 301–310. [Google Scholar] [CrossRef]
- Ouziad, F.; Hildebrandt, U.; Schmelzer, E.; Bothe, H. Differential gene expressions in arbuscular mycorrhizal-colonized tomato grown under heavy metal stress. J. Plant Physiol. 2005, 162, 634–649. [Google Scholar] [CrossRef] [Green Version]
- Cicatelli, A.; Lingua, G.; Todeschini, V.; Biondi, S.; Torrigiani, P.; Castiglione, S. Arbuscular mycorrhizal fungi restore normal growth in a white poplar clone grown on heavy metal-contaminated soil, and this is associated with upregulation of foliar metallothionein and polyamine biosynthetic gene expression. Ann. Bot. 2010, 106, 791–802. [Google Scholar] [CrossRef]
- Berta, M.; Giovannelli, A.; Potenza, E.; Traversi, M.L.; Racchi, M.L. Type 3 metallothioneins respond to water deficit in leaf and in the cambial zone of white poplar (Populus alba). J. Plant Physiol. 2009, 166, 521–530. [Google Scholar] [CrossRef]
- Gu, L.; Pallardy, S.G.; Hosman, K.P.; Sun, Y. Drought-influenced mortality of tree species with different predawn leaf water dynamics in a decade-long study of a central US forest. Biogeosciences 2015, 12, 2831–2845. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Anandhan, S.; Singh, S.; Patade, V.Y.; Ahmed, Z.; Pande, V. Metallothionein-like gene from Cicer microphyllum is regulated by multiple abiotic stresses. Protoplasma 2010, 248, 839–847. [Google Scholar] [CrossRef]
- Ren, Y.; Zhao, J. Functional analysis of the rice metallothionein gene OsMT2b promoter in transgenic Arabidopsis plants and rice germinated embryos. Plant Sci. 2009, 176, 528–538. [Google Scholar] [CrossRef]
- Dong, C.-J.; Wang, Y.; Yu, S.-S.; Liu, J.-Y. Characterization of a Novel Rice Metallothionein Gene Promoter: Its Tissue Specificity and Heavy Metal Responsiveness. J. Integr. Plant Biol. 2010, 52, 914–924. [Google Scholar] [CrossRef]
- Xue, T.; Li, X.; Zhu, W.; Wu, C.; Yang, G.; Zheng, C. Cotton metallothionein GhMT3a, a reactive oxygen species scavenger, increased tolerance against abiotic stress in transgenic tobacco and yeast. J. Exp. Bot. 2008, 60, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Hassinen, V.H.; Tervahauta, A.I.; Schat, H.; Kärenlampi, S.O. Plant metallothioneins—Metal chelators with ROS scavenging activity? Plant Biol. 2010, 13, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, G.; Kata, A.; Goc, A.; Szechyńska-Hebda, M.; Skrzypek, E. Characteristics of the plant ascorbate peroxidase family. Acta Biol. Crac. Ser. Bot. 2007, 49, 7–17. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence 5′–3′ | Application |
|---|---|---|
| BnMT1_for BnMT1_rev | TGGCAGGTTCTAACTGTGGA CAAATGAAAACATTATACACCACACA | expression of BnMT1 gene |
| BnMT2_for BnMT2_rev | TCAATTTGATTAACATTCTCTGCT AAGCCTGCAGCCATTATTACA | expression of BnMT2 gene |
| BnMT3_for BnMT3_rev | GCAAAACAACAAAACACACACA CCATTACATCACACACCATGC | expression of BnMT3 gene |
| Bn5S_for Bn5S_rev | AGTCGCACAAATCGTGTCTG TCCATGCTCTCAGCATCAAC | expression of Bn5S reference gene |
| Variants of Experience | Length of Roots (mm) | Number of Lateral Roots | Length of Shoots (mm) | Number of Leaves |
|---|---|---|---|---|
| control | 114.6 ± 16.9 a | 40.2 ± 7.3 ab | 54.0 ± 10.7 ab | 3.0 ± 1.1 ab |
| L. laccata | 131.4 ± 25.0 a | 35.0 ± 19.6 ab | 58.6 ± 18.9 ab | 3.6 ± 0.9 a |
| C. tuberosa | 127.2 ± 12.6 a | 47.7 ± 9.3 ab | 62.8 ± 10.7 a | 3.1 ± 0.3 ab |
| Clitocybe sp. | 81.5 ± 7.4 b | 43.1 ± 7.8 b | 54.3 ± 13.6 ab | 3.0 ± 0.0 ab |
| H. mesophaeum | 112.5 ± 34.1 a | 38.7 ± 7.7 b | 59.8 ± 12.2 ab | 3.0 ± 0.5 ab |
| C. olla | 117.0 ± 32.3 a | 61.0 ± 26.6 a | 63.4 ± 13.3 a | 3.6 ± 0.8 a |
| Variants of Experience | Chlorophyll (mg cm3) | Length of Shoots (mm) | Number of Leaves | Fresh Biomass (g) | Dry Biomass (g) |
|---|---|---|---|---|---|
| control | 0.815 ± 0.412 a | 665.3 ± 121.9 a | 7.3 ± 1.4 bc | 209.40 ± 1.77 b | 17.54 ± 0.67 b |
| L. laccata | 0.718 ± 0.412 abc | 609.2 ± 169.4 ab | 8.4 ± 3.5 abc | 264.26 ± 11.87 a | 21.58 ± 1.23 a |
| C. tuberosa | 0.387 ± 0.222 d | 473.8 ± 91.2 bcd | 8.4 ± 2.6 abc | 46.69 ± 1.57 d | 3.68 ± 0.55 e |
| Clitocybe sp. | 0.579 ± 0.237 c | 530.0 ± 122.1 bcd | 9.1 ± 1.9 ab | 172.56 ± 3.42 c | 11.29 ± 1.13 d |
| H. mesophaeum | 0.624 ± 0.290 abc | 646.60 ± 137.53 abc | 6.8 ± 1.3 c | 166.75 ± 3.98 c | 11.64 ± 0.96 d |
| C. olla | 0.636 ± 0.307 abc | 507.00 ± 126.67 bcd | 9.8 ± 2.0 a | 195.42 ± 5.06 b | 14.54 ± 1.42 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dąbrowska, G.B.; Garstecka, Z.; Trejgell, A.; Dąbrowski, H.P.; Konieczna, W.; Szyp-Borowska, I. The Impact of Forest Fungi on Promoting Growth and Development of Brassica napus L. Agronomy 2021, 11, 2475. https://doi.org/10.3390/agronomy11122475
Dąbrowska GB, Garstecka Z, Trejgell A, Dąbrowski HP, Konieczna W, Szyp-Borowska I. The Impact of Forest Fungi on Promoting Growth and Development of Brassica napus L. Agronomy. 2021; 11(12):2475. https://doi.org/10.3390/agronomy11122475
Chicago/Turabian StyleDąbrowska, Grażyna B., Zuzanna Garstecka, Alina Trejgell, Henryk P. Dąbrowski, Wiktoria Konieczna, and Iwona Szyp-Borowska. 2021. "The Impact of Forest Fungi on Promoting Growth and Development of Brassica napus L." Agronomy 11, no. 12: 2475. https://doi.org/10.3390/agronomy11122475
APA StyleDąbrowska, G. B., Garstecka, Z., Trejgell, A., Dąbrowski, H. P., Konieczna, W., & Szyp-Borowska, I. (2021). The Impact of Forest Fungi on Promoting Growth and Development of Brassica napus L. Agronomy, 11(12), 2475. https://doi.org/10.3390/agronomy11122475