
Pathogenicity of Cordyceps javanica (Hypocreales: Cordycipitaceae) to Diaphorina citri (Hemiptera: Liviidae) Adults, with Ultrastructural Observations on the Fungal Infection Process

 ,
,  ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Plants and Fungus
2.3. Molecular Characterization of Entomopathogenic Fungi
2.4. Preparation of Conidia
2.5. Inoculation of D. citri with Entomopathogenic Fungi
2.6. Psyllid Mortality Bioassays
2.7. Light Microscopy (LM) and Electron Microscopy (EM)
2.8. Statistical Analysis
3. Results
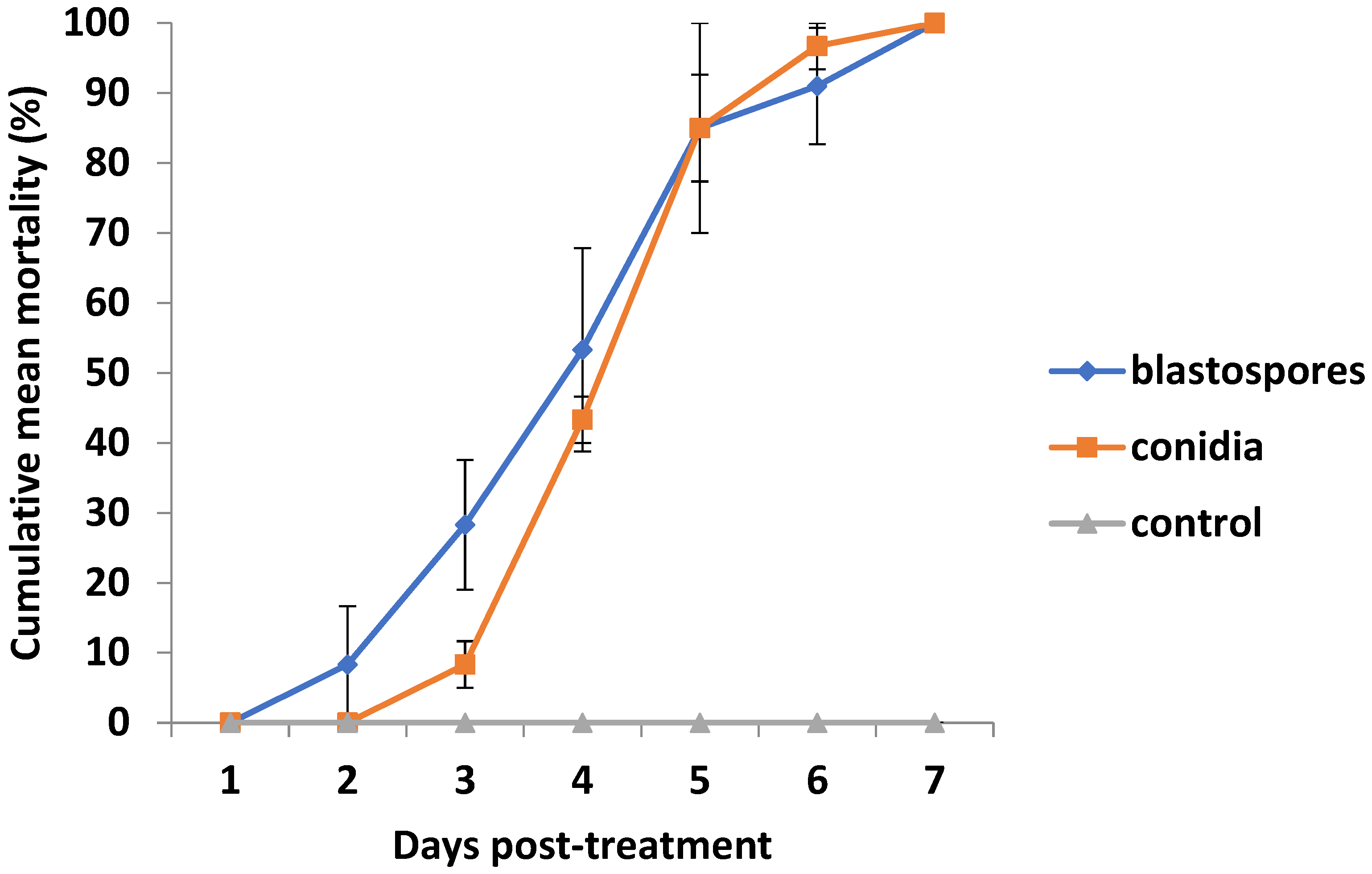
3.1. Percent Psyllid Mortality
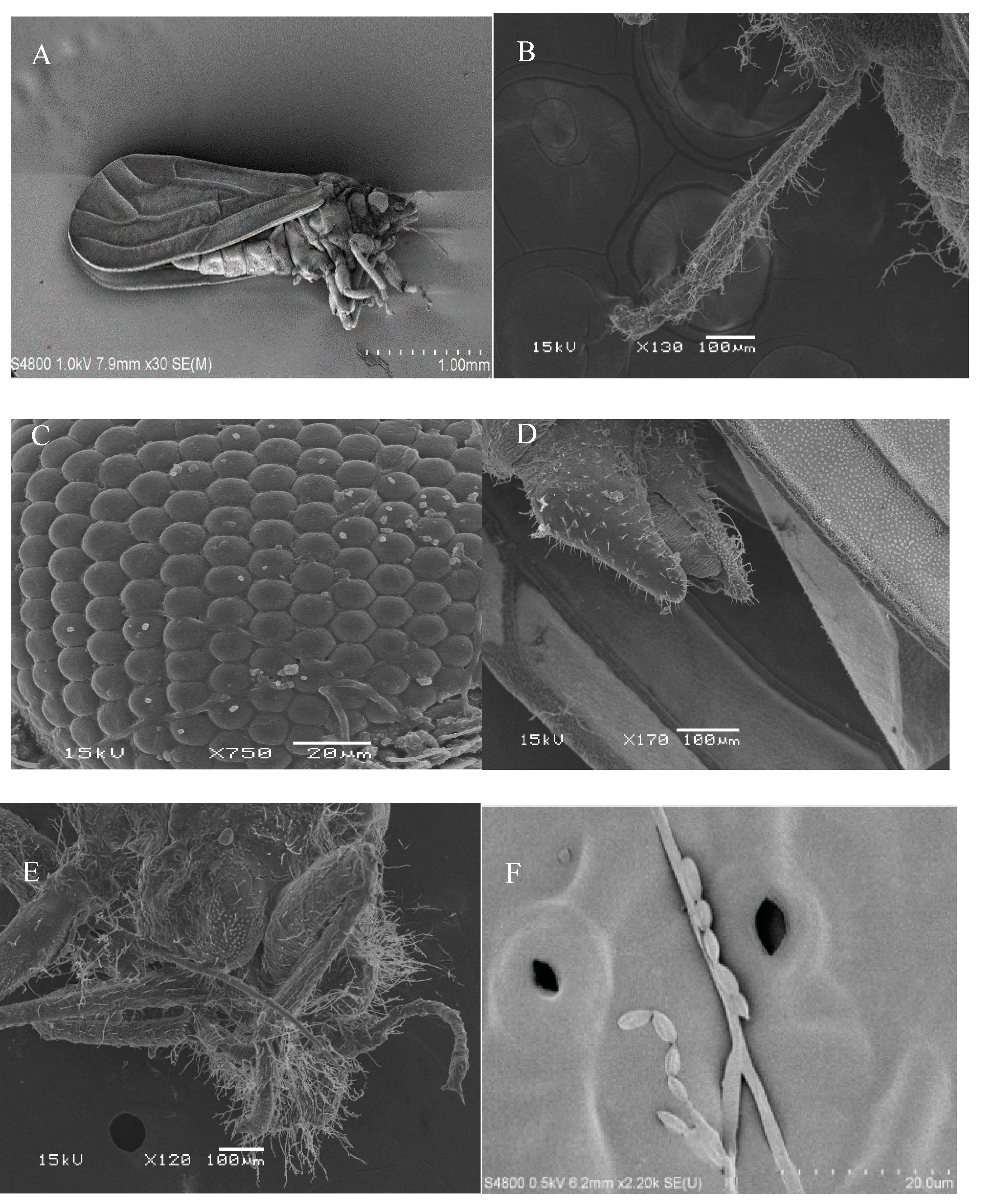
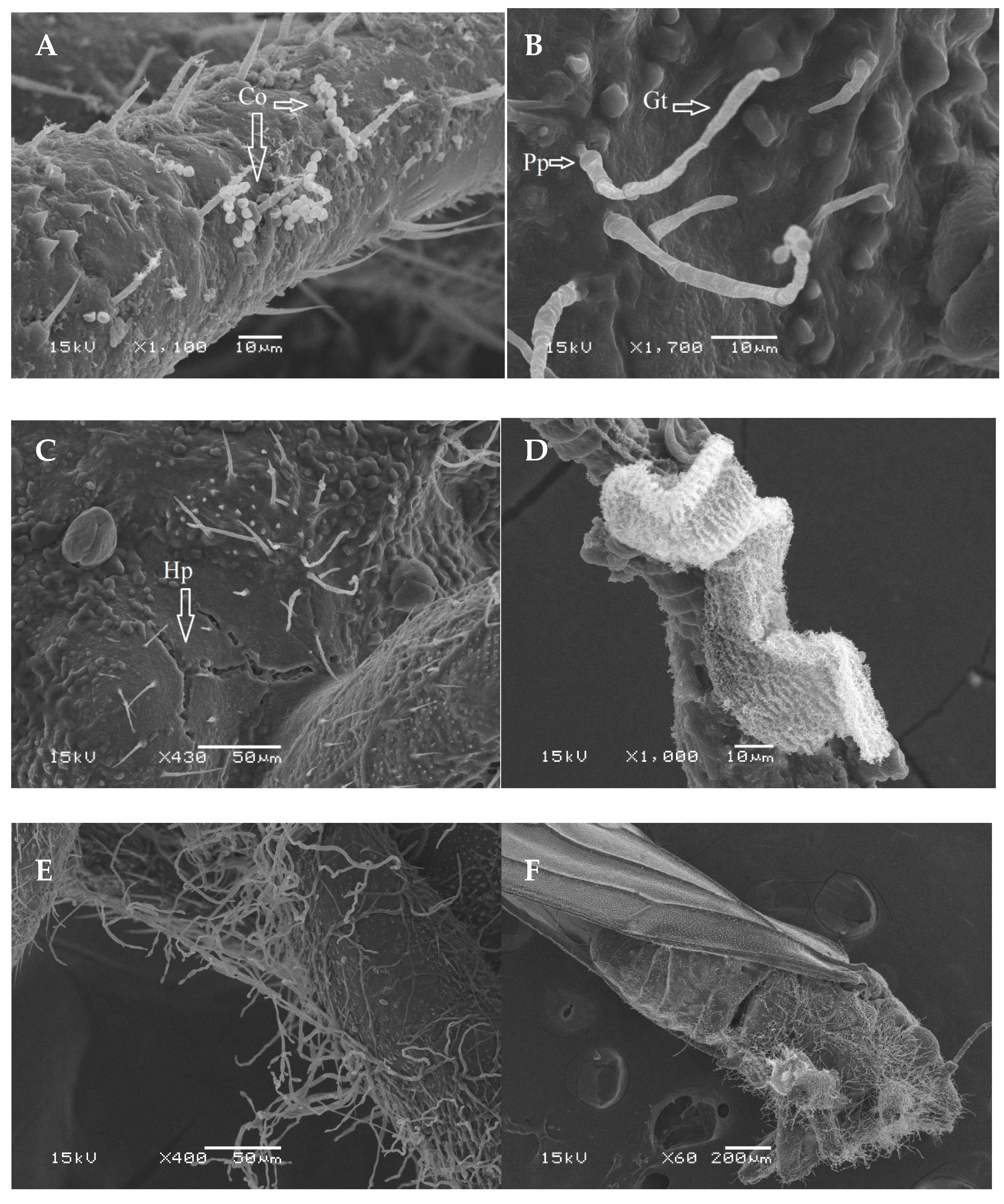
3.2. Scanning Electron Microscopy
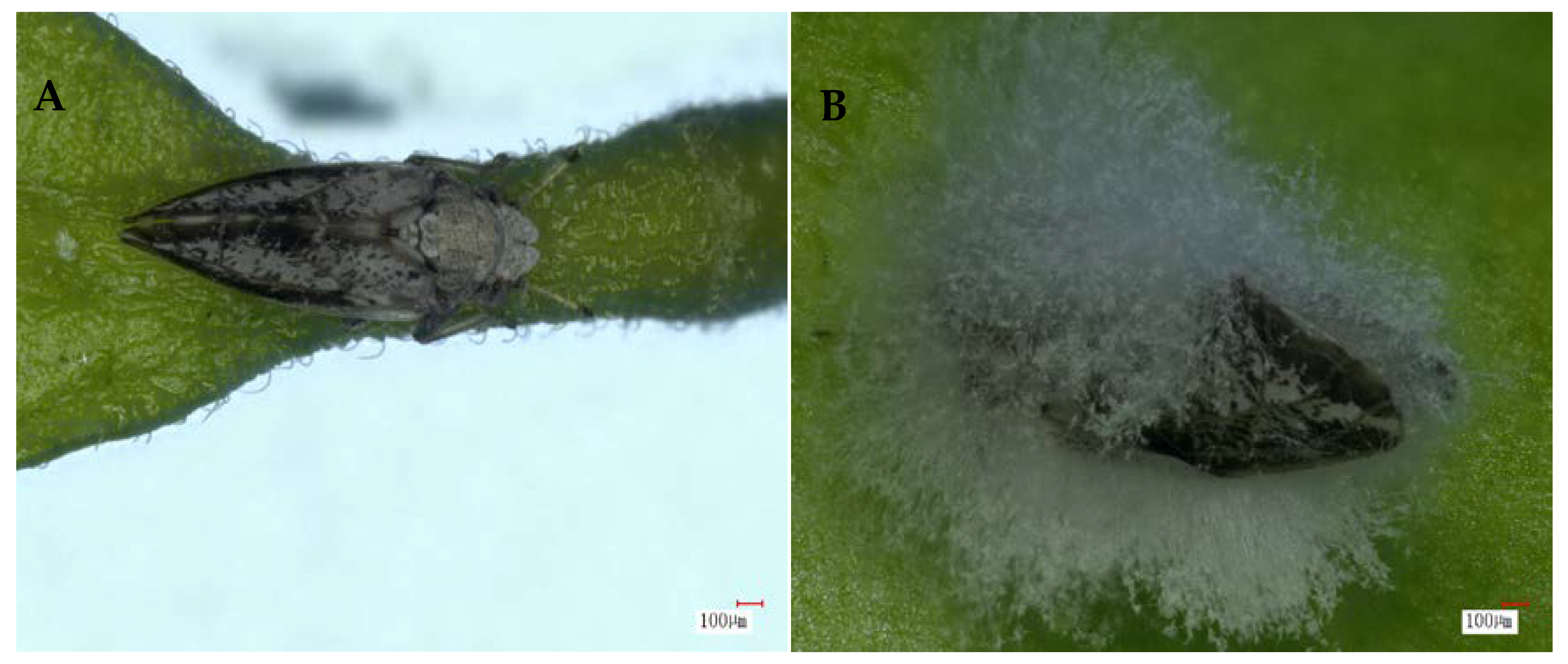
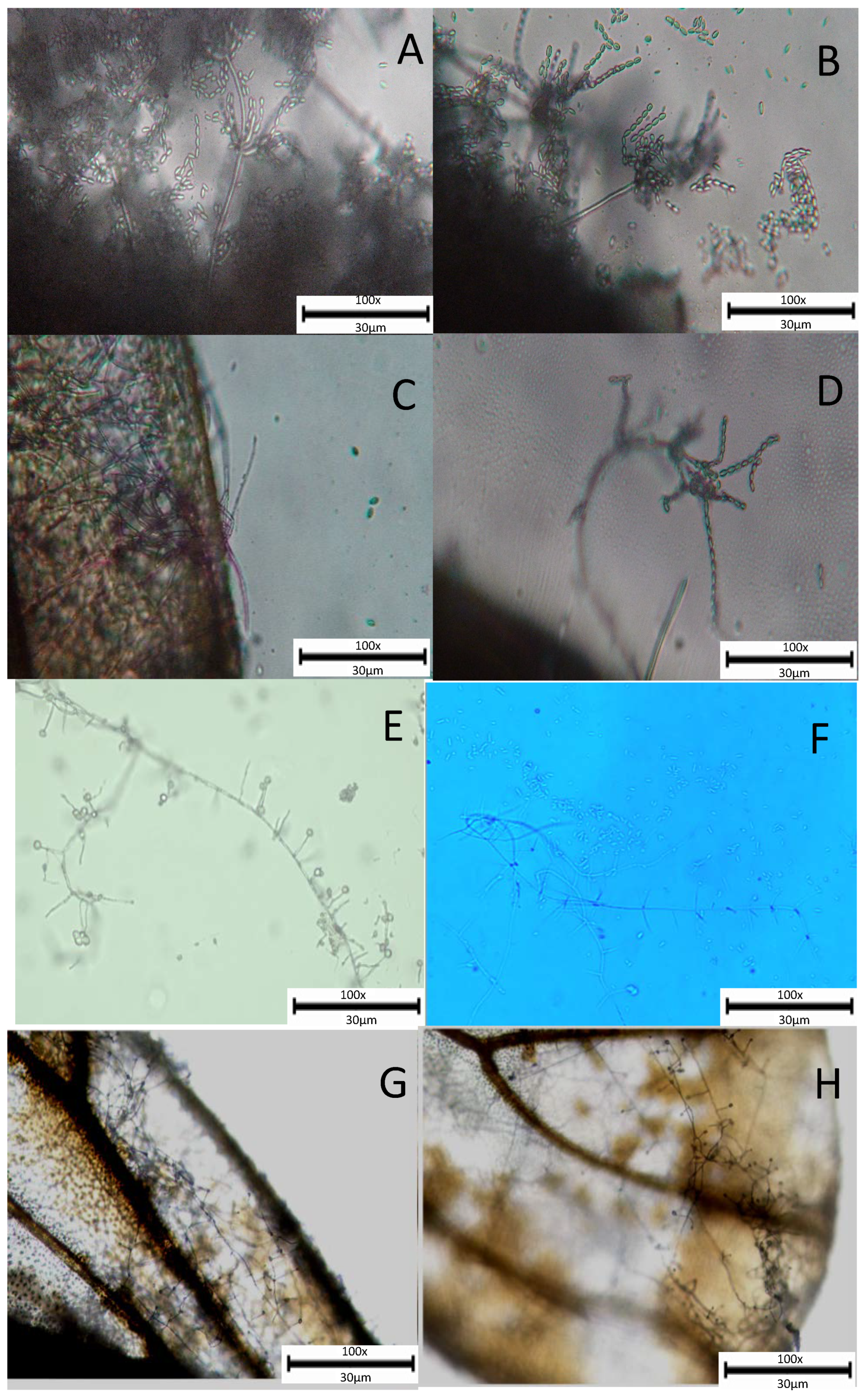
3.3. Light Microscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hussain, M.; Akutse, K.S.; Ravindran, K.; Lin, Y.; Bamisile, B.S.; Qasim, M.; Dash, C.K.; Wang, L. Effects of different temperature regimes on survival of Diaphorina citri and its endosymbiotic bacterial communities. Environ. Microbiol. 2017, 19, 3439–3449. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Cortés, K.; Guzmán-Franco, A.; González-Hernández, H.; Ortega-Arenas, L.; Villanueva-Jiménez, J.; Robles-Bermúdez, A. Susceptibility of Diaphorina citri (Hemiptera: Liviidae) and its parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) to entomopathogenic fungi under laboratory conditions. Neotrop. Entomol. 2018, 47, 131–138. [Google Scholar] [CrossRef]
- Boina, D.R.; Rogers, M.E.; Wang, N.; Stelinski, L.L. Effect of pyriproxyfen, a juvenile hormone mimic, on egg hatch, nymph development, adult emergence and reproduction of the Asian citrus psyllid, Diaphorina citri Kuwayama. Pest Manag. Sci. Former. Pestic. Sci. 2010, 66, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, J.A.; Stansly, P.A. Rate, placement and timing of aldicarb applications to control Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae), in oranges. Pest Manag. Sci. Former. Pestic. Sci. 2008, 64, 1159–1169. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, S.; Karimi, J.; Kamali, S.; Moghadam, E.M. Biocontrol of Planococcus citri (Hemiptera: Pseudococcidae) by Lecanicillium longisporum and Lecanicillium lecanii under laboratory and greenhouse conditions. J. Asia-Paci. Entomol. 2017, 20, 605–612. [Google Scholar] [CrossRef]
- Beloti, V.; Santos, F.; Alves, G.; Bento, J.; Yamamoto, P. Curry leaf smells better than citrus to females of Diaphorina citri (Hemiptera: Liviidae). Arth. Plant Interac. 2017, 11, 709–716. [Google Scholar] [CrossRef]
- Gallou, A.; Serna-Domínguez, M.G.; Berlanga-Padilla, A.M.; Ayala-Zermeño, M.A.; Mellín-Rosas, M.A.; Montesinos-Matías, R.; Arredondo-Bernal, H.C. Species clarification of Isaria isolates used as biocontrol agents against Diaphorina citri (Hemiptera: Liviidae) in Mexico. Fungal Bioloy 2016, 120, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Boopathi, T.; Karuppuchamy, P.; Singh, S.B.; Kalyanasundaram, M.; Mohankumar, S.; Ravi, M. Microbial control of the invasive spiraling whitefly on cassava with entomopathogenic fungi. Braz. J. Microbiol. 2015, 46, 1077–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, T.; Takatsuka, J.; Shimazu, M. Characterization of Paecilomyces cinnamomeus from the camellia whitefly, Aleurocanthus camelliae (Hemiptera: Aleyrodidae), infesting tea in Japan. J. Invert. Pathol. 2012, 110, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Diao, H.; Liang, L.; Hao, C.; Arthurs, S.; Ma, R. Pathogenicity of Isaria fumosorosea to Bemisia tabaci, with some observations on the fungal infection process and host immune response. J. Invert. Pathol. 2015, 130, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Chow, A.; Dunlap, C.A.; Jackson, M.A.; Flores, D.; Patt, J.M.; Sétamou, M. Oviposition behavior and survival of Tamarixia radiata (Hymenoptera: Eulophidae), an ectoparasitoid of the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Liviidae), on hosts exposed to an entomopathogenic fungus, Isaria fumosorosea (Hypocreales: Cordycipitaceae), under laboratory conditions. J. Econ. Entomol. 2016, 109, 1995–2005. [Google Scholar]
- Kumar, V.; Avery, P.B.; Ahmed, J.; Cave, R.D.; McKenzie, C.L.; Osborne, L.S. Compatibility and efficacy of Isaria fumosorosea with horticultural oils for mitigation of the Asian citrus psyllid, Diaphorina citri (Hemiptera: Liviidae). Insects 2017, 8, 119. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Cheng, B.; Du, D.; Hu, X.; Peng, A.; Pu, Z.; Zhang, X.; Huang, Z.; Chen, G. Morphological, molecular and virulence characterization of three Lencanicillium species infecting Asian citrus psyllids in Huangyan citrus groves. J. Invert. Pathol. 2015, 125, 45–55. [Google Scholar] [CrossRef]
- Milosavljević, I.; Amrich, R.; Strode, V.; Hoddle, M.S. Modeling the phenology of Asian citrus psyllid (Hemiptera: Liviidae) in urban southern california: Effects of environment, habitat, and natural enemies. Environ. Entomol. 2018, 47, 233–243. [Google Scholar] [CrossRef]
- Lopez-Llorca, L.; Carbonell, T.; Salinas, J. Colonization of plant waste substrates by entomopathogenic and mycoparasitic fungi—a SEM study. Micron 1999, 30, 325–333. [Google Scholar] [CrossRef]
- Lopez-Llorca, L.V.; Carbonell, T.; Gomez-Vidal, S. Degradation of insect cuticle by Paecilomyces farinosus proteases. Mycol. Progress 2002, 1, 249–256. [Google Scholar] [CrossRef]
- Meyer, J.M.; Hoy, M.A.; Boucias, D.G. Isolation and characterization of an Isaria fumosorosea isolate infecting the Asian citrus psyllid in Florida. J. Invert. Pathol. 2008, 99, 96–102. [Google Scholar] [CrossRef]
- Kwiatkowski, N.P.; Babiker, W.M.; Merz, W.G.; Carroll, K.C.; Zhang, S.X. Evaluation of nucleic acid sequencing of the D1/D2 region of the large subunit of the 28S rDNA and the internal transcribed spacer region using SmartGene IDNS software for identification of filamentous fungi in a clinical laboratory. J. Mol. Diag. 2012, 14, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Petti, C.; Bosshard, P.; Brandt, M.; Clarridge, J.; Feldblyum, T.; Foxall, P.; Furtado, M.; Pace, N.; Procop, G. Interpretive Criteria for Identification of Bacteria and Fungi by DNA Target Sequencing; Approved Guideline; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2008; Volume 28. [Google Scholar]
- Gurulingappa, P.; Mc Gee, P.; Sword, G.A. In vitro and in planta compatibility of insecticides and the endophytic entomopathogen, Lecanicillium lecanii. Mycopathologia 2011, 172, 161–168. [Google Scholar] [CrossRef]
- Avery, P.B.; Wekesa, V.W.; Hunter, W.B.; Hall, D.G.; McKenzie, C.L.; Osborne, L.S.; Powell, C.A.; Rogers, M.E. Effects of the fungus Isaria fumosorosea (Hypocreales: Cordycipitaceae) on reduced feeding and mortality of the Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Bio. Sci. Tech. 2011, 21, 1065–1078. [Google Scholar] [CrossRef]
- Lu, L.; Cheng, B.; Hu, X.; Pu, Z.; Chen, G. Genetic diversity of Lecanicillium lecanii and its pathogenicity against Diaphorina citri. J. Zhejiang Univ. (Agric. Life Sci.) 2015, 41, 34–43. [Google Scholar]
- Anderson, C.M.; McGee, P.A.; Nehl, D.B.; Mensah, R.K. The fungus Lecanicillium lecanii colonises the plant Gossypium hirsutum and the aphid Aphis gossypii. Aust. Mycol. 2007, 26, 65–70. [Google Scholar]
- Hoy, M.A.; Singh, R.; Rogers, M.E. Evaluations of a novel isolate of Isaria fumosorosea for control of the Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Florida Entomol. 2010, 93, 24–32. [Google Scholar] [CrossRef]
- Sewify, G.; Mabrouk, A. The susceptibility of different stages of citrus brown mite Eutetranychus orientalis Oudemans (family: Tetranychidae) to entomopathogenic fungus Verticillium lecanii. In Proceedings of the Vth International Colloquium on Invertebrate Pathology and Microbial Control, Adelaide, Australia, 20–24 August 1990. [Google Scholar]
- Liu, W.; Xie, Y.; Xue, J.; Zhang, Y.; Zhang, X. Ultrastructural and cytochemical characterization of brown soft scale Coccus hesperidum (Hemiptera: Coccidae) infected by the Lecanicillium lecanii (Ascomycota: Hypocreales). Micron 2011, 42, 71–79. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.; Walters, K.F. Pathogenicity of the entomopathogenic fungus, Lecanicillium muscarium, against the sweetpotato whitefly Bemisia tabaci under laboratory and glasshouse conditions. Mycopathologia 2005, 160, 315–319. [Google Scholar] [CrossRef]
- Avery, P.B.; Pick, D.A.; Aristizábal, L.F.; Kerrigan, J.; Powell, C.A.; Rogers, M.E.; Arthurs, S.P. Compatibility of Isaria fumosorosea (Hypocreales: Cordycipitaceae) blastospores with agricultural chemicals used for management of the Asian citrus psyllid, Diaphorina citri (Hemiptera: Liviidae). Insects 2013, 4, 694–711. [Google Scholar] [CrossRef]
- Pick, D.A.; Avery, P.B.; Qureshi, J.A.; Arthurs, S.P.; Powell, C.A. Field persistence and pathogenicity of Cordyceps fumosorosea for management of Diaphorina citri. Biocontrol Sci. Technol. 2021, 1–12. [Google Scholar] [CrossRef]
- Ammar, E.-D.; Alessandro, R.; Shatters, R.G., Jr.; Hall, D.G. Behavioral, ultrastructural and chemical studies on the honeydew and waxy secretions by nymphs and adults of the Asian citrus psyllid Diaphorina citri (Hemiptera: Psyllidae). PLoS ONE 2013, 8, e64938. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.M.; Hoy, M.A.; Boucias, D.G. Morphological and molecular characterization of a Hirsutella species infecting the Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae), in Florida. J. Invert. Pathol. 2007, 95, 101–109. [Google Scholar] [CrossRef]
- Hussain, M.; Akutse, K.S.; Lin, Y.; Chen, S.; Huang, W.; Zhang, J.; Idrees, A.; Qiu, D.; Wang, L. Susceptibilities of Candidatus Liberibacter asiaticus—Infected and noninfected Diaphorina citri to entomopathogenic fungi and their detoxification enzyme activities under different temperatures. MicrobiologyOpen 2018, 7, e00607. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xie, Y.; Xue, J.; Gao, Y.; Zhang, Y.; Zhang, X.; Tan, J. Histopathological changes of Ceroplastes japonicus infected by Lecanicillium lecanii. J. Invert. Pathol. 2009, 101, 96–105. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.A.; de Souza, W. An electron microscopic study of penetration by Trypanosoma rangeli into midgut cells of Rhodnius prolixus. J. Invert. Pathol. 2001, 77, 22–26. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, M.; Avery, P.B.; Zhu, W.; Pitino, M.; Arthurs, S.P.; Wang, L.; Qiu, D.; Mao, R. Pathogenicity of Cordyceps javanica (Hypocreales: Cordycipitaceae) to Diaphorina citri (Hemiptera: Liviidae) Adults, with Ultrastructural Observations on the Fungal Infection Process. Agronomy 2021, 11, 2476. https://doi.org/10.3390/agronomy11122476
Hussain M, Avery PB, Zhu W, Pitino M, Arthurs SP, Wang L, Qiu D, Mao R. Pathogenicity of Cordyceps javanica (Hypocreales: Cordycipitaceae) to Diaphorina citri (Hemiptera: Liviidae) Adults, with Ultrastructural Observations on the Fungal Infection Process. Agronomy. 2021; 11(12):2476. https://doi.org/10.3390/agronomy11122476
Chicago/Turabian StyleHussain, Mubasher, Pasco B. Avery, Wenchao Zhu, Marco Pitino, Steven P. Arthurs, Liande Wang, Dongliang Qiu, and Runqian Mao. 2021. "Pathogenicity of Cordyceps javanica (Hypocreales: Cordycipitaceae) to Diaphorina citri (Hemiptera: Liviidae) Adults, with Ultrastructural Observations on the Fungal Infection Process" Agronomy 11, no. 12: 2476. https://doi.org/10.3390/agronomy11122476
APA StyleHussain, M., Avery, P. B., Zhu, W., Pitino, M., Arthurs, S. P., Wang, L., Qiu, D., & Mao, R. (2021). Pathogenicity of Cordyceps javanica (Hypocreales: Cordycipitaceae) to Diaphorina citri (Hemiptera: Liviidae) Adults, with Ultrastructural Observations on the Fungal Infection Process. Agronomy, 11(12), 2476. https://doi.org/10.3390/agronomy11122476