
Mild Salinity Stimulates Biochemical Activities and Metabolites Associated with Anticancer Activities in Black Horehound (Ballota nigra L.)

 , and
, and
Abstract
:1. Introduction
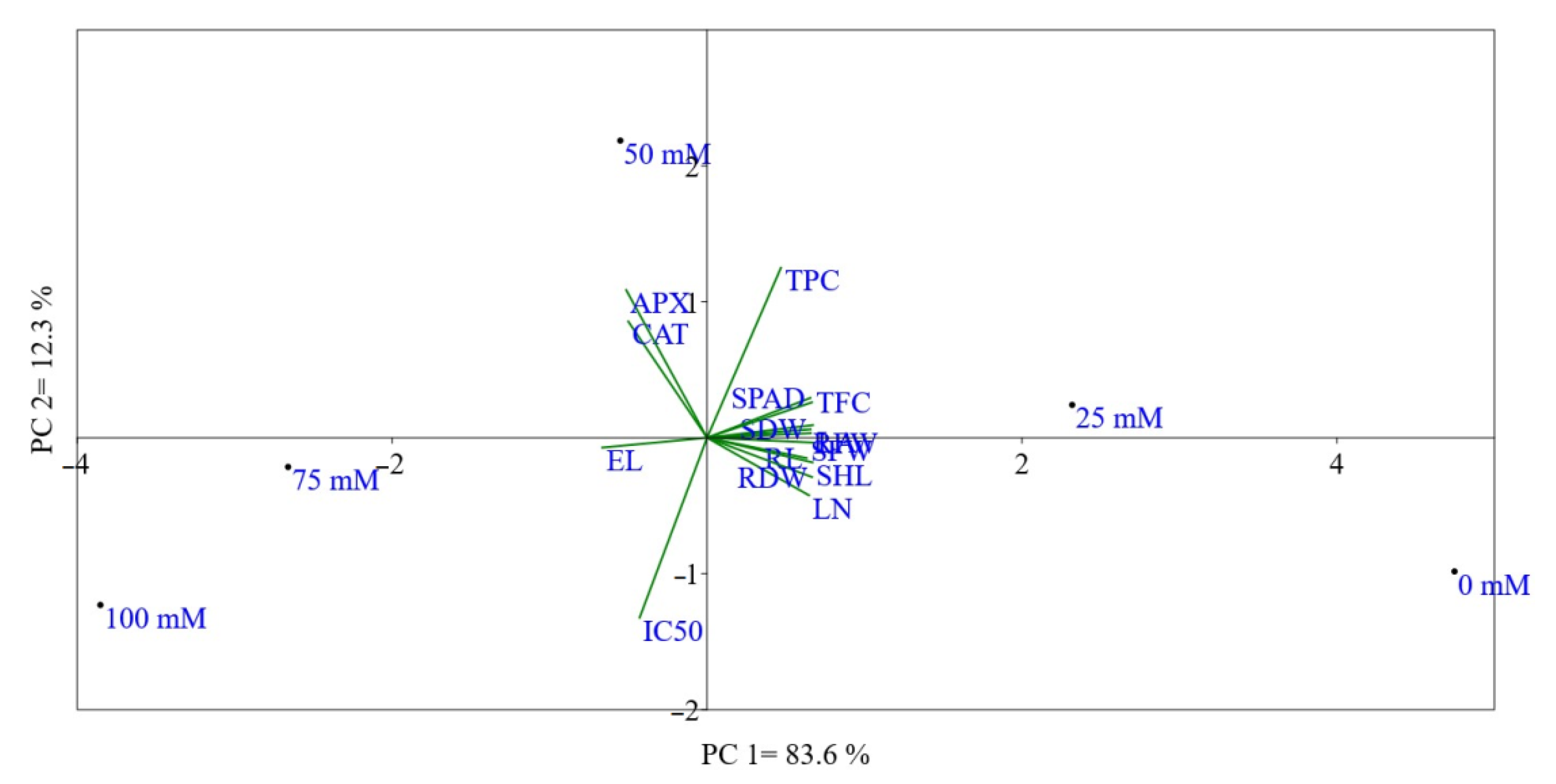
2. Results and Discussion
2.1. Soil Salinity Inhibited Plant Growth and Yield
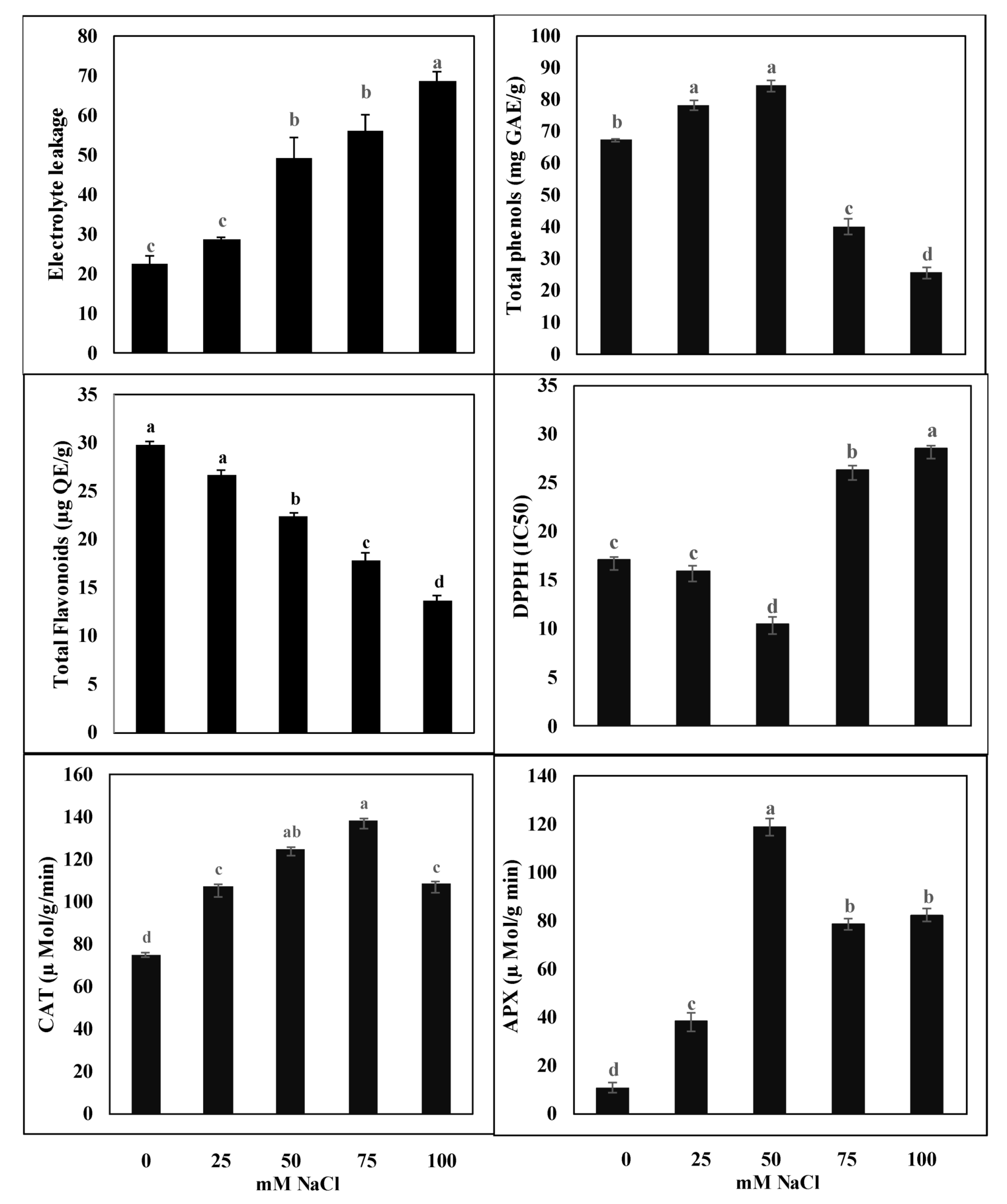
2.2. Effect of Salinity on Cell Membrane Integrity
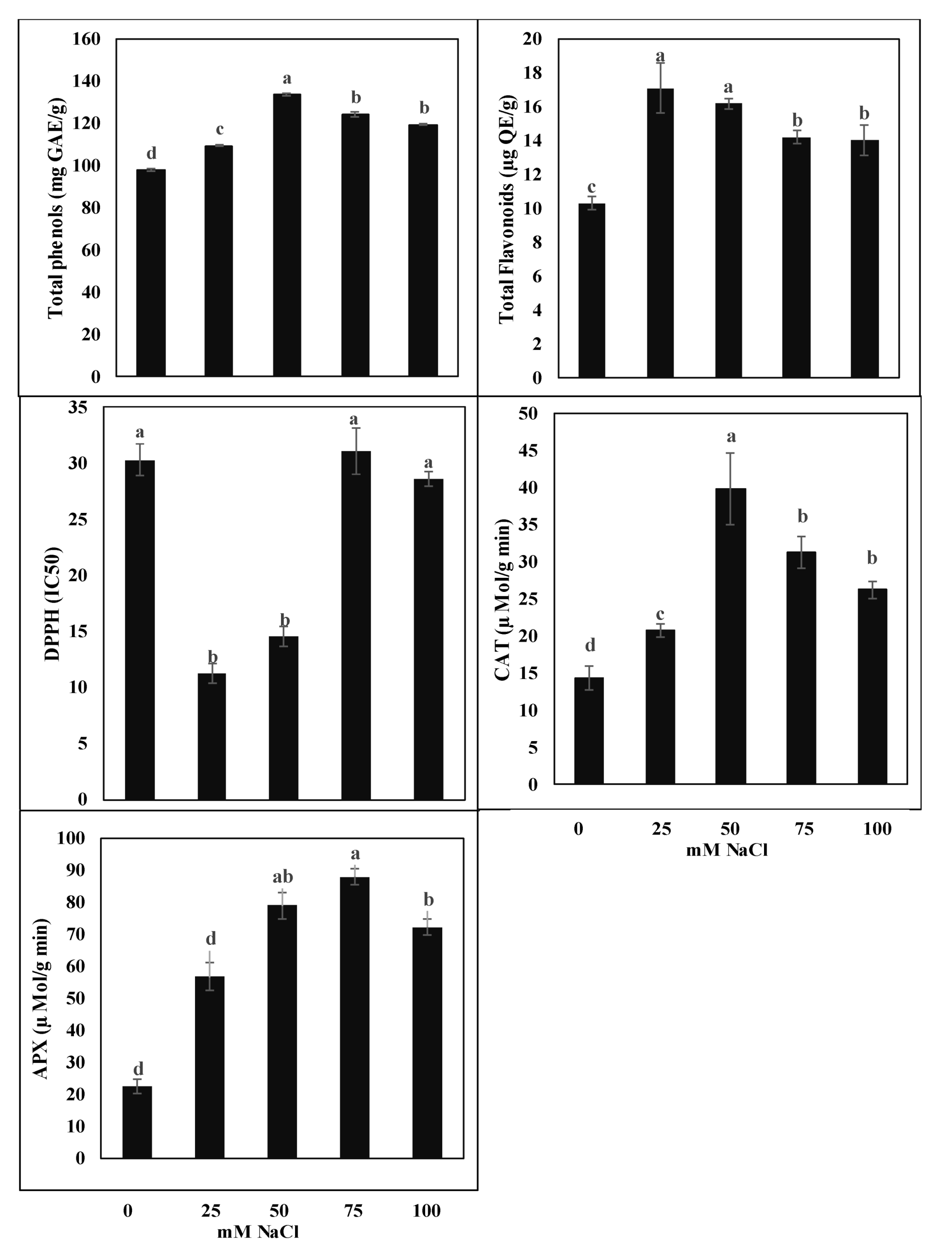
2.3. Effect of Salinity on Total Phenols, Flavonoids, and Antioxidant Enzyme Activities
2.4. Antioxidant Activity
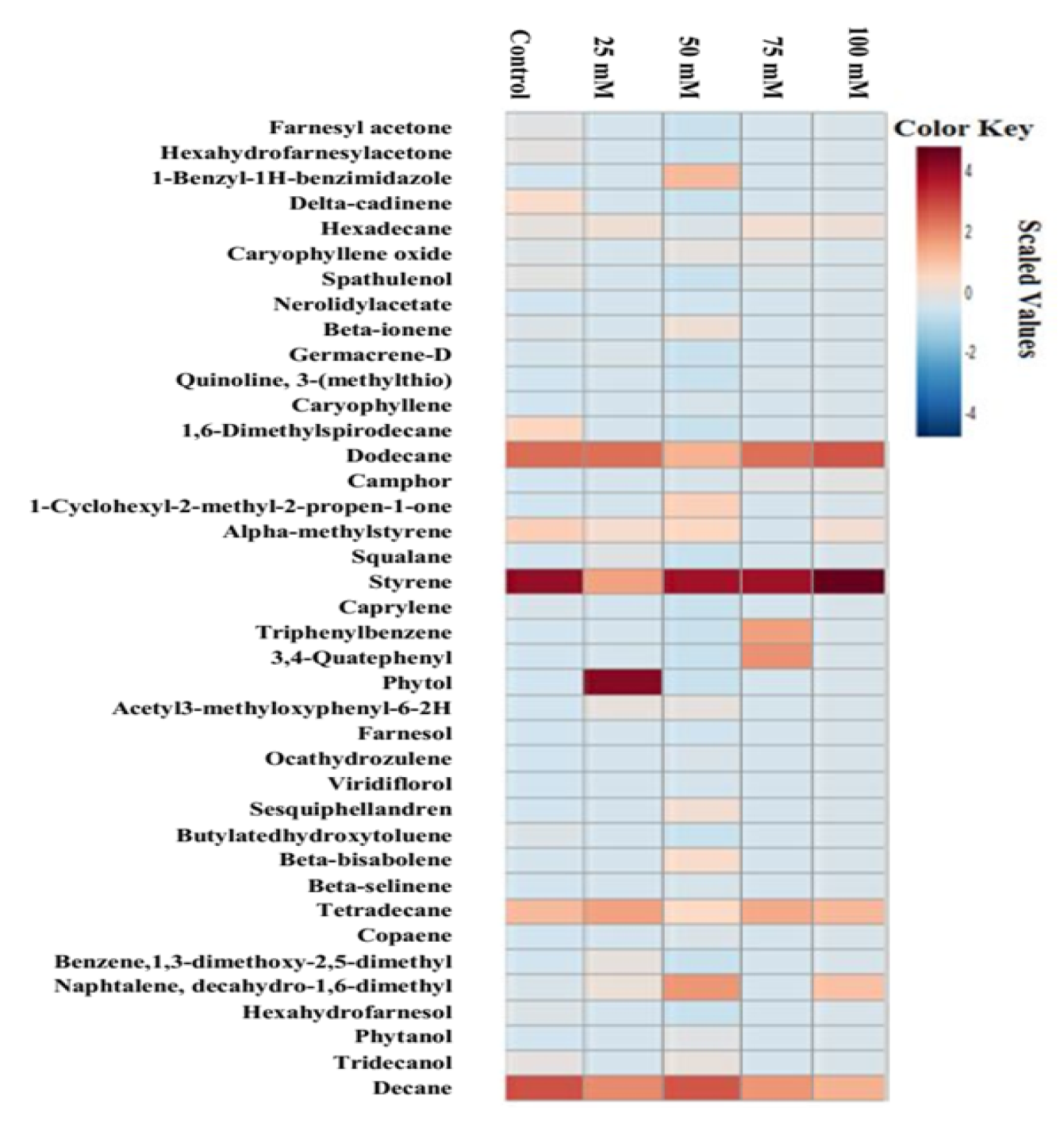
2.5. Salinity Effect on Metabolite Synthesis
3. Conclusions
4. Material and Methods
4.1. Plant and Growth, Maintenance, and Salinity Treatment In Vivo
4.2. Plant Harvest and Sampling
4.3. Plant Growth, Maintenance, and Salinity Treatment In Vitro
4.4. Measurement of Growth and Morphological Characteristics
4.5. Measurement of Electrolyte Leakage (%)
4.6. Antioxidant Activity Assay/Radical Scavenging Assay
4.7. Measurement of Total Flavonoids
4.8. Total Phenol Measurement
4.9. Measurement of Catalase (CAT) and Ascorbic Peroxidase (APX) Activity
4.10. Identification and Characterization of Metabolites
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flowers, T.J.; Gaur, P.M.; Gowda, C.L.; Krishnamurthy, L.; Samineni, S.; Siddique, K.H.; Colmer, T.D. Salt sensitivity in chickpea. Plant Cell Environ. 2010, 33, 490–509. [Google Scholar] [CrossRef] [Green Version]
- Sairam, R.K.; Rao, K.V.; Srivastava, G.C. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Xie, W.; Wu, L.; Zhang, Y.; Wu, T.; Li, X.; Ouyang, Z. Effects of straw application on coastal saline topsoil salinity and wheat yield trend. Soil Tillage Res. 2017, 169, 1–6. [Google Scholar] [CrossRef]
- Fathizad, H.; Ardakani, M.A.H.; Sodaiezadeh, H.; Kerry, R.; Taghizadeh-Mehrjardi, R. Investigation of the spatial and temporal variation of soil salinity using random forests in the central desert of Iran. Geoderma 2020, 365, 114233. [Google Scholar] [CrossRef]
- Jordanov, D.; Velčev, V.; Kožuharov, S.I. Flora na Narodna Republika Bulgariâ; Izd-vo na Bulgarskata Akademiâ na Naukite: Sofia, Bulgaria, 1982. [Google Scholar]
- Sahin, F.P.; Toker, M.C.; Ezer, N. Botanical properties of a Mild Sedative: Ballota nigra L. subsp. nigra. FABAD J. Pharm. Sci. 2005, 30, 94. [Google Scholar]
- Kennedy, J. Herb and supplement use in the US adult population. Clin. Ther. 2005, 27, 1847–1858. [Google Scholar] [CrossRef]
- Bandaranayake, W.M. Quality Control, Screening, Toxicity, and Regulation of Herbal Drugs. In Modern Phytomedicine. Turning Medicinal Plants into Drugs; Ahmad, I., Aqil, F., Owais, M., Eds.; Wiley-VCH GmbH & Co. KGa A: Weinheim, Germany, 2007; pp. 25–57. [Google Scholar]
- Belaqziz, R.; Romane, A.; Abbad, A. Salt stress effects on germination, growth, and essential oil content of an endemic thyme species in Morocco (Thymus maroccanus Ball.). J. Appl. Sci. Res. 2009, 5, 858–863. [Google Scholar]
- Said-Al Ahl, H.A.H.; Omer, E.A. Medicinal and aromatic plants production under salt stress. A review. Herba Polonica 2011, 57, 72–87. [Google Scholar]
- Najafian, S.; Khoshkhui, M.; Tavallali, V.; Saharkhiz, M.J. Effect of salicylic acid and salinity in thyme (Thymus vulgaris L.): Investigation on changes in gas exchange, water relations, and membrane stabilization and biomass accumulation. Aust. J. Basic Appl. Sci. 2009, 3, 2620–2626. [Google Scholar]
- Oueslati, S.; Karray-Bouraoui, N.; Attia, H.; Rabhi, M.; Ksouri, R.; Lachaal, M. Physiological and antioxidant responses of Mentha pulegium (Pennyroyal) to salt stress. Acta Physiol. Plant. 2010, 32, 289–296. [Google Scholar] [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Amanifar, S.; Khodabandeloo, M.; Fard, E.M.; Askari, M.S.; Ashrafi, M. Alleviation of salt stress and changes in glycyrrhizin accumulation by arbuscular mycorrhiza in liquorice (Glycyrrhiza glabra) grown under salinity stress. Environ. Exp. Bot. 2019, 160, 25–34. [Google Scholar] [CrossRef]
- Farsaraei, S.; Moghaddam, M.; Pirbalouti, G.A. Changes in growth and essential oil composition of sweet basil in response to salinity stress and super absorbents application. Sci. Hortic. 2020, 271, 109465. [Google Scholar] [CrossRef]
- Jaiswal, D.; Pandey, A.; Mukherjee, A.; Agrawal, M.; Agrawal, S.B. Alterations in growth, antioxidative defense, and medicinally important compounds of Curcuma caesia Roxb. under elevated ultraviolet-B radiation. Environ. Exp. Bot. 2020, 177, 104152. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, A. Seed Priming Technology in the Amelioration of Salinity Stress in Plants. In Advances in Seed Priming; Springer: Singapore, 2018; pp. 81–93. [Google Scholar]
- Bistgani, E.Z.; Hashemi, M.; DaCosta, M.; Craker, L.; Maggi, F.; Morshedloo, R.M. Effect of salinity stress on the physiological characteristics, phenolic compounds, and antioxidant activity of Thymus vulgaris L. and Thymus daenensis Celak. Ind. Crops Prod. 2019, 135, 311–320. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Michailidi, E.; Tzortzakis, N. Physiological and biochemical responses of Lavandula angustifolia to salinity under mineral foliar application. Front. Plant Sci. 2018, 9, 489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neffati, M.; Marzouk, B. Changes in essential oil and fatty acid composition in coriander (Coriandrum sativum L.) leaves under saline conditions. Ind. Crops Prod. 2008, 28, 137–142. [Google Scholar] [CrossRef]
- Aziz, E.E.; Al-Amier, H.; Craker, L.E. Influence of salt stress on growth and essential oil production in peppermint, pennyroyal, and apple mint. J. Herbs Spices Med. Plants 2008, 14, 77–87. [Google Scholar] [CrossRef]
- Said-Al Ahl, H.A.H.; Meawad, A.A.; Abou-Zeid, E.N.; Ali, M.S. Response of different basil varieties to soil salinity. Int. Agrophys. 2010, 24, 183–188. [Google Scholar]
- Gebauer, J.; El-Siddig, K.; Salih, A.A.; Ebert, G. Tamarindus indica L. seedlings are moderately salt tolerant when exposed to NaCl-Induced salinity. Sci. Hortic. 2004, 103, 1–8. [Google Scholar] [CrossRef]
- Tavakkoli, E.; Rengasamy, P.; McDonald, G.K. High concentrations of Na+ and Cl–ions in soil solution have simultaneous detrimental effects on growth of faba bean under salinity stress. J. Exp. Bot. 2010, 61, 4449–4459. [Google Scholar] [CrossRef]
- Cramer, G.R. Sodium-Calcium Interactions Under Salinity Stress. In Salinity: Environment-Plants-Molecules; Springer: Dordrecht, The Netherlands, 2008; pp. 205–227. [Google Scholar]
- Bhatt, R.M.; Rao, N.S. Influence of pod load on the response of okra to water stress. Indian J. Plant Physiol. 2005, 10, 54. [Google Scholar]
- Rezgui, M.; Majdoub, N.; Ben-Kaab, S.; Marzouk, B.; Gouia, H.; Araújo, M.E.M.; Ben-Kaab, L.B. How salt stress represses the biosynthesis of marrubiin and disturbs the antioxidant activity of Marrubium vulgare L. Pol. J. Environ. Stud. 2017, 26, 267–277. [Google Scholar] [CrossRef]
- Naeem, M.S.; Jin, Z.L.; Wan, G.L.; Liu, D.; Liu, H.B.; Yoneyama, K.; Zhou, W.J. 5–Aminolevulinic acid improves photosynthetic gas exchange capacity and ion uptake under salinity stress in oilseed rape (Brassica napus L.). Plant Soil 2010, 332, 405–415. [Google Scholar] [CrossRef]
- Harrathi, J.; Attia, H.; Neffati, M.; Hosni, K.; Marzouk, B.; Lachâal, M.; Karray-Bouraoui, N. Salt effects on shoot growth and essential oil yield and composition in safflower (Carthamus tinctorius L.). J. Essent. Oil Res. 2013, 25, 482–487. [Google Scholar] [CrossRef]
- Venkateswarlu, B.; Ramesh, K. Cell membrane stability and biochemical response of cultured cells of groundnut under polyethylene glycol induced water stress. Plant Sci. 1992, 90, 179–185. [Google Scholar] [CrossRef]
- Demidchik, V.; Straltsova, D.; Medvedev, S.S.; Pozhvanov, G.A.; Sokolik, A.; Yurin, V. Stress-Induced electrolyte leakage: The role of K+-Permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp. Bot. 2014, 65, 1259–1270. [Google Scholar] [CrossRef]
- Kumar, S.A.; Kumari, P.H.; Jawahar, G.; Prashanth, S.; Suravajhala, P.; Katam, R.; Sivan, P.; Rao, K.S.; Kirti, P.B.; Kishor, P.K. Beyond just being foot soldiers–osmotin like protein (OLP) and chitinase (Chi11) genes act as sentinels to confront salt, drought, and fungal stress tolerance in tomato. Environ. Exp. Bot. 2016, 132, 53–65. [Google Scholar] [CrossRef]
- Manian, R.; Anusuya, N.; Siddhuraju, P.; Manian, S. The antioxidant activity and free radical scavenging potential of two different solvent extracts of Camellia sinensis (L.) O. Kuntz, Ficus bengalensis L. and Ficus racemosa L. Food Chem. 2008, 107, 1000–1007. [Google Scholar] [CrossRef]
- Makowczynska, J.; Grzegorczyk, K.I.; Wysokinska, H. Antioxidant activity of tissue culture-Raised Ballota nigra L. plants grown ex vitro. Acta Pol. Pharm. 2015, 72, 769–775. [Google Scholar]
- Kanatt, S.R.; Chander, R.; Sharma, A. Antioxidant potential of mint (Mentha spicata L.) in radiation-Processed lamb meat. Food Chem. 2007, 100, 451–458. [Google Scholar] [CrossRef]
- Zhou, Y.; Tang, N.; Huang, L.; Zhao, Y.; Tang, X.; Wang, K. Effects of Salt Stress on Plant Growth, Antioxidant Capacity, Glandular Trichome Density, and Volatile Exudates of Schizonepeta tenuifolia Briq. Int. J. Mol. Sci. 2018, 19, 252. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants, and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-Scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Guan, L.; Sun, Y.; Zhu, Y.; Liu, L.; Lu, R.; Zhang, A. Calcium and ZmCCaMK are involved in brassinosteroid-Induced antioxidant defense in maize leaves. Plant Cell Physiol. 2015, 56, 883–896. [Google Scholar] [CrossRef] [Green Version]
- Aghaei, K.; Ehsanpour, A.A.; Komatsu, S. Potato responds to salt stress by increased activity of antioxidant enzymes. J. Integr. Plant Biol. 2009, 51, 1095–1103. [Google Scholar] [CrossRef]
- Vafadar, F.; Amooaghaie, R.; Ehsanzadeh, P.; Ghanadian, M. Salinity stress alters ion homeostasis, antioxidant activities, and the production of rosmarinic acid, luteolin, and apigenin in Dracocephalum kotschyi Boiss. Biologia 2020, 75, 2147–2158. [Google Scholar] [CrossRef]
- Dvorak, P.; Krasylenko, Y.; Zeiner, A.; Samaj, J.; Takac, T. Signaling toward ROS-Scavenging enzymes in plants. Front. Plant Sci. 2020, 11, 2178. [Google Scholar]
- Green, T.; Lee, R.; Toghill, A.; Meadowcroft, S.; Lund, V. FosterThe toxicity of styrene to the nasal epithelium of mice and rats: Studies on the mode of action and relevance to humans. J. Chem. Biol. Interact. 2001, 137, 185–202. [Google Scholar] [CrossRef]
- Singh, S.; Singh, J. Percutaneous absorption, biophysical, macroscopic barrier properties of porcine skin exposed to major components of JP-8 jet fuel. Environ. Toxicol. Pharm. 2003, 14, 77–85. [Google Scholar] [CrossRef]
- Yang, J.H.; Lee, C.H.; Monteiro-Riviere, N.A.; Riviere, J.E.; Tsang, C.; Chou, C. Toxicity of jet fuel aliphatic and aromatic hydrocarbon mixtures on human epidermal keratinocytes: Evaluation based on in vitro cytotoxicity and interleukin-8 release. Arch Toxicol. 2006, 80, 508–523. [Google Scholar] [CrossRef]
- Khadhri, A.; Neffati, M.; Smiti, S.; Nogueira, J.M.F.; Araújo, M.E.M. Influence of salt stress on essential oil yield and composition of lemon grass (Cymbopogon schoenanthus L. Spreng. ssp. Laniger (Hook) Maire et Weil). Nat. Prod. Res. 2011, 25, 108–117. [Google Scholar] [CrossRef]
- Yeo, S.K.; Ali, A.Y.; Hayward, O.A.; Turnham, D.; Jackson, T.; Bowen, I.D.; Clarkson, R. β-Bisabolene, a Sesquiterpene from the Essential Oil Extract of Opoponax (Commiphora guidottii), Exhibits Cytotoxicity in Breast Cancer Cell Lines. Phytother. Res. 2016, 30, 418–425. [Google Scholar] [CrossRef]
- Park, K.R.; Nam, D.; Yun, H.M.; Lee, S.G.; Jang, H.J.; Sethi, G.; Cho, S.K.; Ahn, K.S. β-Caryophyllene oxide inhibits growth and induces apoptosis through the suppression of PI3K/AKT/mTOR/S6K1 pathways and ROS-mediated MAPKs activation. Cancer Lett. 2011, 312, 178–188. [Google Scholar] [CrossRef]
- Shin, Y.; Lee, Y. Enhanced biodegradation of n-Hexadecane in solid-Phase of soil by employing immobilized Pseudomonas aeruginosa on size-Optimized coconut fibers. Toxicol Res. 2013, 29, 257–261. [Google Scholar] [CrossRef] [Green Version]
- Lemes, S.R.; Ribeiro e Silva, C.; Jefferson, H.V.; Chen-Chen, L.; Silva Lima, R.; Caridad, N.P.; Montes de Sousa, M.A.; de Melo Reis, P.R.; da Silva, N.J. Presence of antigenotoxic and anticytotoxic effects of the chalcone 1E,4E-1-(4-Chlorophenyl)-5-(2,6,6-Trimethylcyclohexen-1-Yl)Penta-1,4-Dien-3-One using in vitro and in vivo assays. Drug Chem. Toxicol. 2020, 43, 383–390. [Google Scholar] [CrossRef]
- Rivero-Cruz, B.; Rojas, M.A.; Rodríguez-Sotres, R.; Cerda-García-Rojas, C.M.; Mata, R. Smooth muscle relaxant action of benzyl benzoates and salicylic acid derivatives from Brickellia veronicaefolia on isolated guinea-pig ileum. Planta Med. 2005, 71, 320–325. [Google Scholar] [CrossRef]
- Alexandra, T.; Marina, I.M.; Daniela, M.; Ioana, S.I.; Maria, B.; Radu, R.; Maria, T.A.; Tudor, S.; Maria, G. Autophagy—A Hidden but Important Actor on Oral Cancer Scene. Int. J. Mol. Sci. 2020, 21, 9325. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Sui, X.; Wang, Y.; Wang, W.; Haibo, W.; Zhang, F.; Yuna, T.; Feng, Z. Sesquiterpene-Neolignans from Manglietia hookeri. Nat. Prod. Res. 2016, 30, 1477–1483. [Google Scholar] [CrossRef]
- Said-Al Ahl, H.A.; Abou-Ellail, M.; Omer, E.A. Harvest date and genotype influences growth characters and essential oil production and composition of Petroselinum crispum plants. J. Chem. Pharm. Res. 2016, 8, 992–1003. [Google Scholar]
- Taarit, M.B.; Msaada, K.; Hosni, K.; Hammami, M.; Kchouk, M.E.; Marzouk, B. Plant growth, essential oil yield, and composition of sage (Salvia officinalis L.) fruits cultivated under salt stress conditions. Ind. Crops Prod. 2009, 30, 333–337. [Google Scholar] [CrossRef]
- Hajieghrari, M.; Hejazi, P. Analysis of heartsease (Viola tricolor L.) flavonoid glycosides by micro-Liquid chromatography coupled to multistage mass spectrometry. J. Hazard. Mater. 2020, 389, 122134. [Google Scholar] [CrossRef]
- Akter, K.; Barnes, E.C.; Loa-Kum-Cheung, W.L.; Yin, P.; Kichu, M.; Brophy, J.J.; Barrow, R.A.; Imchen, I.; Vemulpad, S.R.; Jamie, J.F. Antimicrobial and antioxidant activity and chemical characterization of Erythrina stricta Roxb. (Fabaceae). J. Ethnopharmacol. 2016, 185, 171–181. [Google Scholar] [CrossRef]
- Pavithra, P.S.; Sreevidya, N.; Verma, R.S. Antibacterial activity and chemical composition of essential oil of Pamburus missionis. J. Ethnopharmacol. 2009, 124, 151–153. [Google Scholar] [CrossRef]
- Yin, X.; Liu, Y.; Pan, J.; Ye, H.L.; Sun, Y.; Zhao, D.Y.; Kuang, H.X.; Yang, B.Y. Melongenaterpenes A–L, Vetispirane-Type Sesquiterpenoids from the Roots of Solanum melongena. J. Nat. Prod. 2019, 82, 3242–3248. [Google Scholar] [CrossRef]
- Vukic, M.D.; Vukovic, N.L.; Djelic, G.T.; Obradovic, A.; Kacaniova, M.M.; Markovic, S.; Popović, S.; Baskić, D. Phytochemical analysis, antioxidant, antibacterial and cytotoxic activity of different plant organs of Eryngium serbicum L. Ind. Crops Prod. 2018, 115, 88–97. [Google Scholar] [CrossRef]
- Tounekti, T.; Khemira, H. NaCl stress-Induced changes in the essential oil quality and abietane diterpene yield and composition in common sage. J. Intercult. Ethnopharmacol. 2015, 4, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Tung, L.D.; Serrano, E.P. Effects of warm water in breaking dormancy of rice seed. Omonrice 2011, 18, 129–136. [Google Scholar]
- Huang, D.J.; Chun-Der, L.I.N.; Hsien-Jung, C.H.E.N.; Yaw-Huei, L.I.N. Antioxidant and antiproliferative activities of sweet potato (Ipomoea batatas [L.] LamTainong 57’) constituents. Bot. Bull. Acad. Sin. 2004, 45, 179–186. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2017; Volume 456. [Google Scholar]
- Linstrom, P.J.; Mallard, W.G. The NIST Chemistry WebBook: A chemical data resource on the internet. J. Chem. Eng. Data 2001, 46, 1059–1063. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean of Squares (Greenhouse) | Mean of Squares | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S.V | d.f | LN | LA | RL | RFW | RDW | SHL | SFW | SDW | SPAD | d.f | LN | SHE | NB |
| Repeat | 2 | 45.3 | 1431.9 | 0.3 | 40.7 | 0.8 | 0.1 | 158.5 | 7.05 | 9.2 | - | - | - | - |
| Salinity | 4 | 52.7 ** | 6690.9 ** | 20.4 ** | 519.8 ** | 6.0 ** | 20.4 ** | 47.0 ** | 12.2 ** | 106.2 ** | 4 | 729.1 ** | 4.5 ** | 19.7 ** |
| Error | 8 | 21.0 | 257.1 | 0.4 | 8.8 | 0.2 | 0.4 | 24.32 | 0.7 | 1.4 | 10 | 4.3 | 0.3 | 0.6 |
| C.V | 10.5 | 3.8 | 2.1 | 8.1 | 5.7 | 2.1 | 8.2 | 6.9 | 3.7 | 10.2 | 18.7 | 20.6 | ||
| NaCl (mM) | ||||||
|---|---|---|---|---|---|---|
| Condition | Characteristics | 0 | 25 | 50 | 75 | 100 |
| Greenhouse | Leaf Number | 66.5 ± 2.2 a | 48.3 ± 2.1 b | 37.2 ± 1.2 c | 33 ± 1.2 c | 32 ± 0.8 c |
| Leaf Area | 476.3 ± 7.2 a | 446 ± 14.5 a | 396.5 ± 8.5 b | 372.5 ± 10.2 c | 351.2 ± 15.6 c | |
| Root Length | 33.2 ± 1.2 a | 33.6 ± 1.5 a | 29.3 ± 0.8 b | 28.3 ± 0.4 b | 28.2 ± 0.3 b | |
| Root Fresh Weight | 52.9 ± 1.9 a | 46.5 ± 1.6 b | 34.8 ± 2.5 c | 27.7 ± 1.2 d | 20.8 ± 1.1 e | |
| Root Dry Weight | 9.2 ± 0.4 a | 8.1 ± 0.3 b | 6.6 ± 0.4 c | 6.2 ± 0.4 c | 5.9 ±0. 5 c | |
| Shoot Length | 31.5 ±0.5 a | 28.6 ±0.6 b | 24.2 ±0.6 c | 23.2 ±0.4 c | 21.3 ±0.3 d | |
| Shoot Fresh Weight | 78. ±1.1 a | 64.2 ± 3.6 b | 60.8 ± 3.5 bc | 49.5 ±2.2 cd | 48 ± 2.8 d | |
| Shoot dry Weight | 15.2 ± 1.5 a | 12.9 ± 1.4 b | 12.2 ± 1.8 bc | 11.2 ± 1.4 cd | 9.8 ± 1.3 d | |
| In vitro | SPAD | 40.4 ± 1.5 a | 35.3 ± 0.3 b | 33.5 ± 0.3 b | 29.3 ± 0.2 c | 24.7 ± 1.2 d |
| Leaf Number | 45± 0.6 a | 23.6 ± 1.1 b | 18 ± 1.2 c | 12 ± 1.1 d | 3.7 ± 0.7 e | |
| Shoot Length | 4.6 ± 0.3 a | 3.6 ± 0.6 ab | 2.7 ± 0.5 bc | 2.5 ± 0.4 c | 1.4 ± 0.5 d | |
| Number of branches | 7.8 ± 0.7 a | 4 ± 0.5 b | 3 ± 0.5 b | 3 ± 0.5 b | 1 ± 0.3 d |
| Mean of Squares | ||||||||
|---|---|---|---|---|---|---|---|---|
| Greenhouse | Source of variation | d.f | TPh | Flv | DPP | APX | CAT | EL |
| Repeat | 2 | 4.5 | 0.2 | 0.5 | 2391.3 | 0.8 | 4.63 | |
| Salinity | 4 | 1916.5 ** | 126.5 ** | 171.2 ** | 132,662.0 ** | 1685.1 ** | 1104.0 ** | |
| Error | 8 | 28.7 | 27.8 | 0.6 | 4178.9 | 335.3 | 54.8 | |
| C.V% | 9.1 | 4.1 | 4.1 | 19.6 | 16.5 | 16.4 | ||
| In vitro | Salinity | 4 | 563.6 * | 20.7 ** | 245.9 ** | 1965.1 ** | 284.4 ** | - |
| Error | 8 | 119.6 | 2.8 | 5.6 | 63.0 | 46.9 | - | |
| C.V | 9.3 | 11.6 | 10 | 12.5 | 25.8 | - | ||
| Variables | Principle Components | |
|---|---|---|
| PC1 | PC 2 | |
| Leaf Number | 0.27 | −0.18 |
| Leaf Area | 0.28 | −0.01 |
| Root Length | 0.26 | −0.06 |
| Root Fresh Weight | 0.28 | 0.04 |
| Root Dry Weight | 0.28 | −0.12 |
| Shoot Length | 0.28 | −0.08 |
| Shoot Fresh Weight | 0.28 | 0.01 |
| Shoot Dry Weight | 0.27 | 0.03 |
| SPAD | 0.27 | 0.12 |
| Electrolyte leakage | −0.28 | −0.03 |
| Total phenols (mg GAE/g) | 0.20 | 0.52 |
| Total Flavonoids (µg QE/g) | 0.28 | 0.11 |
| DPPH (IC50) | −0.18 | −0.55 |
| CAT (µ Mol/g/min) | −0.21 | 0.36 |
| APX (µ Mol/g/min) | −0.21 | 0.45 |
| Eigenvalues | 12.5 | 1.8 |
| % of variance | 83.6 | 12.3 |
| Cumulative % | 83.6 | 95.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Younessi-Hamzekhanlu, M.; Dibazarnia, Z.; Oustan, S.; Vinson, T.; Katam, R.; Mahna, N. Mild Salinity Stimulates Biochemical Activities and Metabolites Associated with Anticancer Activities in Black Horehound (Ballota nigra L.). Agronomy 2021, 11, 2538. https://doi.org/10.3390/agronomy11122538
Younessi-Hamzekhanlu M, Dibazarnia Z, Oustan S, Vinson T, Katam R, Mahna N. Mild Salinity Stimulates Biochemical Activities and Metabolites Associated with Anticancer Activities in Black Horehound (Ballota nigra L.). Agronomy. 2021; 11(12):2538. https://doi.org/10.3390/agronomy11122538
Chicago/Turabian StyleYounessi-Hamzekhanlu, Mehdi, Zahra Dibazarnia, Shahin Oustan, Teniyah Vinson, Ramesh Katam, and Nasser Mahna. 2021. "Mild Salinity Stimulates Biochemical Activities and Metabolites Associated with Anticancer Activities in Black Horehound (Ballota nigra L.)" Agronomy 11, no. 12: 2538. https://doi.org/10.3390/agronomy11122538
APA StyleYounessi-Hamzekhanlu, M., Dibazarnia, Z., Oustan, S., Vinson, T., Katam, R., & Mahna, N. (2021). Mild Salinity Stimulates Biochemical Activities and Metabolites Associated with Anticancer Activities in Black Horehound (Ballota nigra L.). Agronomy, 11(12), 2538. https://doi.org/10.3390/agronomy11122538