3.1. Total Polyphenol Content
The combined analysis of variance based on the AMMI 2 model for the total polyphenol content (TPC) in the fruits of bilberry populations during the three years, indicates that all three sources significantly influenced this trait, with a predominant action of population and year–population interaction (
Table 2). Thus, the year–population interaction showed the highest influence on the variability of TPC (58.59%), followed by the population with a contribution of 34.39%, amid the background of lower influence from the climatic conditions during the study (7.02%).
Cumulated, the six populations achieved yearly average values of TPC between 315.20 mg GAE/100 g FW in 2020 and 432.40 mg GAE/100 g FW in 2018, under a middle variability between years (
Table 3). In 2018, the values were significantly higher with 9.77–37.17% compared to those of 2019–2020, with 25% higher accumulation of polyphenols in 2019 compared to 2020.
Regarding the unilateral effect of the bilberry population on TPC, average values from 282.3 mg GAE/100 g FW in the Semenic population to 553.3 mg GAE/100 g FW for the Padis population were registered, amid high inter-population variability. Given the comparisons among the six bilberry populations, it was found that during this period, the Padis population had significantly higher amounts of polyphenols than the other populations, associated with significant increases of 12.17–109.82%. The Cuntu population also stood out, with 32.04–87.09% significantly higher TPC than four other populations. The Muntele Mic, Semenic, and Sadova Noua populations had close TPC values, and were statistically lower compared to the Raul Lung population.
Amid the influence of conditions during 2018–2020 on the TPC in each population, the highest amplitude (475.5 3 mg GAE/100 g FW) was registered in the Padis population, while in the case of the Muntele Mic and Sadova Noua populations, the amplitude was considerably lower.
In 2019, the Muntele Mic, Cuntu, and Padis populations achieved significantly higher TPCs compared to the values for 2018 and 2020, statistically differentiated in favor of those from 2018. The Semenic and Sadova populations recorded significantly higher values in 2018 compared to 2020, higher in turn toward the values from 2019. A special reaction was observed in the case of the Raul Lung population, given that in 2020, it accumulated an amount of TPC significantly higher than in 2018–2019, as opposed to lower values in 2019.
Given the conditions of 2018, a lower inter-population variability of TPC was observed, considering that the populations recorded lower amplitudes, ranging from 266.5 mg GAE/100 g FW in Muntele Mic and 542.7 mg GAE/100 g FW in Padis. The highest values of TPC in 2018 were recorded in the Padis population, which showed significant increases of 2.52–103.6% to most other populations. In the case of the Cuntu and Semenic populations, the TPC was significantly higher than in Sadova Noua and Raul Lung populations, which were statistically undifferentiated.
Regarding the effect of the conditions of 2019 on TPC, it was observed that the populations recorded an amplitude of 659 mg GAE/100 g FW, considerably higher than other years, with values between 144.5 mg GAE/100 g FW in Semenic and 796.3 mg GAE/100 g FW in Padis, amid a very high inter-population variability. Thus, the largest amount of polyphenols was accumulated by the fruit of the Padiş population, associated with significant increases of over 21.63% compared to other populations. In the case of the Cuntu population, the TPC values were significantly higher by 179.8–510.2% compared to the other four populations. In this year, the Semenic, Raul Lung, and Sadova Noua populations accumulated significantly lower amounts of polyphenols in their fruits.
Given the conditions from 2020, the bilberry populations had TPC values ranging from 208.8 mg GAE/100 g FW in Muntele Mic and 579.5 mg GAE/100 g FW in Raul Lung, with an amplitude of 370.7 mg GAE/100 g FW and a high inter-population variability. Thus, the Raul Lung population showed a significantly higher TPC, over 80.64% greater than the other populations. In the case of the Cuntu and Padis populations, the values were 24.31–53.64% significantly higher compared to the Sadova Noua, Semenic, and Muntele Mic populations, which were statistically undifferentiated.
Depending on the values of the ASV index, it is noted that the Semenic, Sadova Noua, and Muntele Mic populations had the highest TPC stability, on a background of a qualitative (different signs of IPCA) type of population–year interaction (
Table 4). In the case of the Padis and Raul Lung populations, the amounts of polyphenols displayed large deviations during the study, associated with a qualitative interaction in the Padis population and a quantitative interaction in the Raul Lung population. In this regard, the low stability of the Raul Lung population was associated with a TPC value below the overall mean, while in the Padis population, the low stability of TPC was correlated with values above the mean.
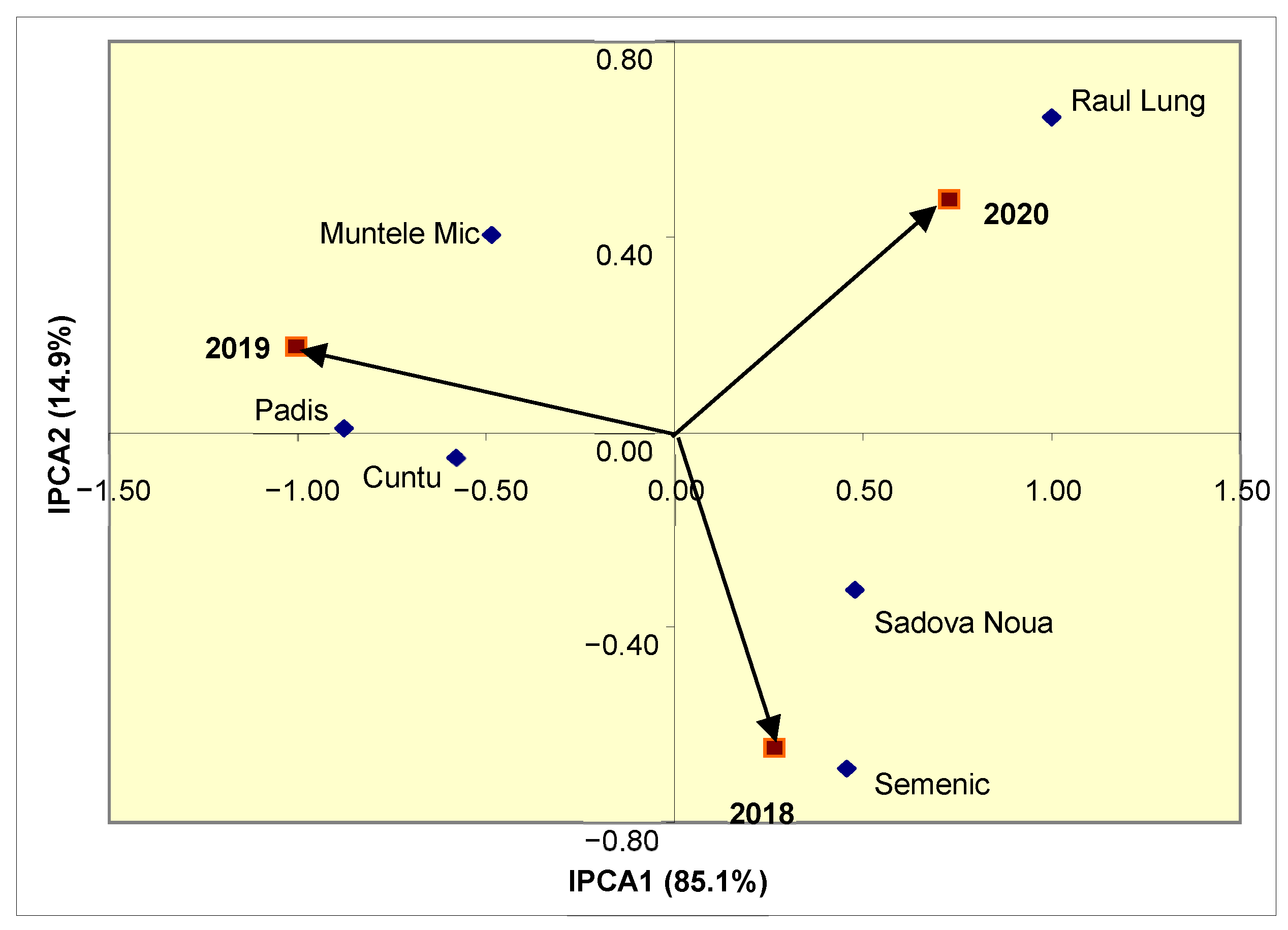
Taking into account the fact that IPCA1 displays a considerable contribution (85.1%) to total variability, depending on the position in the biplot from
Figure 1, it was noted that the Raul Lung and Padis populations had the lowest stability of TPC during the study. Based on the length of the associated vectors, it was found that the conditions of 2019–2020 had high contributions to the population–year interaction for this trait. Depending on the position towards the year vectors, it was observed that the Raul Lung population had a specific adaptation to the conditions of 2020, while the Semenic population efficiently used the conditions of 2018. In the case of the Padis and Cuntu populations, specific adaptations to the conditions of 2019 were observed.
3.2. DPPH Radical Scavenging Activity
For DPPH radical scavenging activity, the results of combined analysis of variance according to the AMMI 2 model, indicates that all three main sources of variation have a significant influence (
Table 5). The contributions of variation sources are differentiated, thus the variability of this indicator was more influenced by the conditions during the three years (63.51%), while the effects of population (14.58%) and year–population interaction (21.90%) were considerably lower. Given that the first two IPCA fully express the effect of population–year interaction, it follows that the AMMI2 model is suitable for assessing the antioxidant activity of fruit extracts in these six populations.
Considering the cumulative effect of the years, during this period (
Table 6), average values of inhibition ranging from 72.09% in 2020 to 81.94% in 2018 were found, with low variation between the three years. Generally, the bilberry populations recorded during 2018–2019 had significant increases of radical scavenging activity, equivalent to values about 8–9% higher than the results of 2020. Additionally, the conditions of 2018 favored the manifestation of a significantly higher inhibition compared to the conditions of 2019.
The average values of radical scavenging activity for the six populations had an amplitude of 5.83%, with limits from 75.58% for the Cuntu population, up to 81.41% for the Padiş population, with low inter-population variability. Compared to the overall mean, the Padis and Raul Lung populations recorded significant increases of 1.97–3.37%, while for other populations, the inhibition was significantly lower than the mean. Based on comparisons between populations, it was found that the Padis population had a radical scavenging activity over 1.40% significantly higher than the other populations, followed by the Raul Lung population, which recorded significant deviations of 2.38–4.43% compared to the rest of the statistically differentiated populations.
Given the population–year interaction, it was observed that under the conditions of 2018, the inhibition recorded values between 78.90% in the Cuntu and 85.72% in the Padis populations, with an amplitude of 6.82% and low variability between populations. Given this year’s conditions, the Padis population had a radical scavenging activity 3.51% significantly greater than the other populations. Additionally, the Muntele Mic, Sadova Noua, and Semenic populations registered inhibitions 1.55–3.31% higher than the Raul Lung and Cuntu populations.
Regarding the radical scavenging activity in 2019, amid an inter-population variability close to the previous year, values between 78.34% in Padis and 81.78% in Muntele Mic were recorded, with an amplitude of 3.44% and a grouping of populations in five classes. Thus, the Muntele Mic population used this year’s conditions at a higher level, showing significantly higher inhibition of over 0.54% compared to the other populations. In the case of the Sadova Noua and Padis populations, close values were observed, associated with significantly lower radical scavenging activity than the Cuntu, Semenic, and Raul Lung populations.
Given the conditions of 2020, the populations registered a high amplitude of 12.50%, ranging between 67.67% in Sadova Noua and 80.17% in Padis. As such, this year the Padis population showed the highest radical scavenging activity, associated with significant increases of over 2.01% compared to other populations. Additionally, the Raul Lung population recorded a significantly higher inhibition of 9.15–10.39% compared to four other populations. The Muntele Mic and Semenic populations had similar antioxidant activities amid an inhibition 1.15–1.67% higher than the Cuntu and Sadova Noua populations.
Considering the effect of the variability in yearly conditions during the study on the radical scavenging activity in each population, it was observed that the Semenic and Sadova Noua populations capitalized the conditions of 2018 at a higher level, registering a significantly higher inhibition compared to the statistically differentiated values of 2019–2020. In the period 2018–2019, the Muntele Mic and Raul Lung populations presented close and significantly higher values of inhibition by 3.08–13.1% compared to 2020. The fruits of the Cuntu population showed significantly higher radical scavenging by 0.74–11.45% in 2019 compared to the values of 2018 and 2020, respectively. Additionally, in the case of the Padis population, the conditions from 2018 favored a significantly higher expression of inhibition compared to the statistically differentiated values from 2019–2020.
Regarding the unilateral effect of extract concentration (
Table S1), the inhibition showed an amplitude of 12.90%, with mean values ranging from 71.66% for 0.67 mg/mL concentration to 84.57% in the case of the maximum concentration. Thus, at the level of the whole study, there is a significant reduction of the radical scavenging activity proportional to the decrease of extract concentration. The rate of inhibition decrease was between 0.67%/mg/mL when the concentration was changed from 10 to 5 mg/mL, and 5.43%/mg/mL when the concentration was reduced from 1.25 to 0.67 mg/mL, respectively.
Regarding the combined effect of the extract concentration and the population on the radical scavenging activity, it is observed that the fruits of the Padis population showed the highest antioxidant activity regardless of extract concentration. For the 10 mg/mL concentration, inhibition registered values between 81.09% in Cuntu and 89.22% in Padis populations, amid significant deviations among most of the populations except for Semenic and Sadova Noua, which showed close values. Under the effect of 5 mg/mL concentration, the radical scavenging activity varied from 78.54% in Cuntu to 85.62% in Padis populations, amid higher values of 81.4–83.08% in Muntele Mic and Raul Lung populations and significantly lower values of 78.73–79.89% in Sadova Noua and Semenic populations. The extracts with a concentration of 2.5 mg/mL showed inhibitions from 75.07% in Cuntu to 81.31% in Padis, associated with a 6.24% amplitude and significant differentiation between populations.
For the 1.25 mg/mL concentration, inhibition registered values between 72.74% in Cuntu and 77.81% in Padiş, with significant deviations among most of the populations, with the exception of Cuntu and Sadova Noua, which recorded close values. Under the effect of 0.67 mg/mL concentration, the radical scavenging activity varied from 70.46% in Cuntu to 73.51% in Raul Lung, associated with values of 71.52–73.09% in Semenic and Padiş, and significantly lower values of 70.47–70.94% in the Sadova Noua and Muntele Mic populations. Thus, the antioxidant activity of the extracts of the Cuntu, Sadova Noua, and Muntele Mic populations at a concentration of 0.67 mg/mL were equivalent to the antioxidant activity of ascorbic acid at a concentration of 0.08 mg/mL (
Table S2). In the case of Padis and Raul Lung populations, during the period of study the antioxidant activity at the concentration of 0.67mg/mL was higher than that of ascorbic acid at a concentration of 0.1 mg/mL.
Based on ASV ranks (
Table 7), it was observed that the highest stability of radical scavenging activity manifested in the Cuntu population on a background of a qualitative population–year interaction, and a value below the overall mean. In the case of the Raul Lung and Padis populations, antioxidant activity registered considerable deviations during the study in the presence of mixed interactions at Padis and a quantitative one at the Raul Lung population, respectively. The Semenic and Sadova Noua populations had a cross interaction, registering values of antioxidant activity in accordance with the variation of the year’s mean.
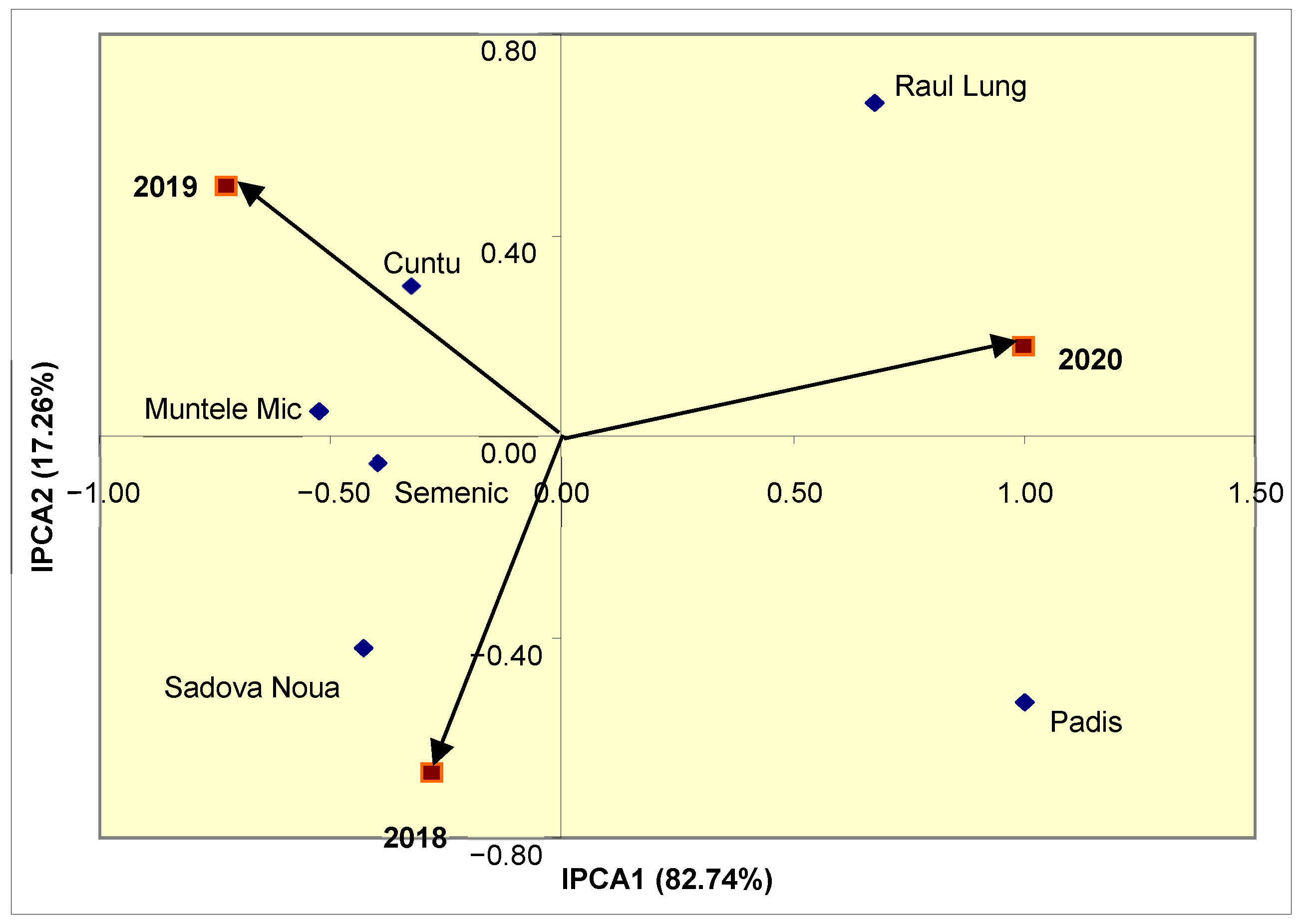
Given that IPCA1 expresses approximately 83% of the variability of the population–year interaction (
Figure 2), and depending on the coordinates of each year, it was found that the conditions of 2020 had a higher contribution to this interaction, while the conditions of 2018 had a lesser influence on the variation of radical scavenging activity in the six bilberry populations.
Depending on the distance from the origin, it was observed that the antioxidant activity of the Padis and Raul Lung populations was strongly influenced by the population–year interaction, while in the Cuntu and Semenic populations, this trait had greater stability. The close position of the Cuntu population towards the 2019 vector indicates a specific adaptation to this year’s conditions, registering the highest value of this trait, while in the Sadova Noua population, the conditions from 2018 favored superior radical scavenging activity.
3.3. Half Maximal Inhibitory Concentration (IC50)
The combined analysis of variance based on the AMMI 2 model for the half maximal inhibitory concentrations (IC
50) of extracts for bilberry populations during three years, indicates that all three sources significantly influenced this trait, with predominant action from population and a lower contribution from year (
Table 8). Thus, population had the highest influence on the variability of IC
50 values (51.86%), followed by the population–year interaction with a contribution of 34.36%, on a background of lower influence of year conditions during the study (13.96%).
Overall, during the study, the half maximal inhibitory concentration (IC
50) had low variability, associated with an amplitude of 0.009 mg/mL, ranging from 0.04 mg/mL in 2020 to 0.049 mg/mL in 2019 (
Table 9). The conditions of 2020 had a positive influence on the antioxidant activity evaluated through IC
50, registering significant increases compared to the results of 2018–2019. Additionally, under the conditions of 2018–2019, the antioxidant activity based on IC
50 registered close values.
In this period, the Muntele Mic and Semenic populations had antioxidant activity significantly higher than the other populations except for Raul Lung, achieving the lowest average values (0.038–0.039 mg/mL) of IC50. The Sadova Noua and Cuntu populations recorded the lowest antioxidant activities, associated with high average values (0.049–0.059 mg/mL) of IC50.
Considering the combined effect of years and populations on the IC50, it was observed that given the conditions of 2018, the populations registered an amplitude of 0.024 mg/mL, with values between 0.037 mg/mL in Muntele Mic and 0.061 mg/mL in Sadova Noua. The highest antioxidant activity was seen in the Muntele Mic, Raul Lung, and Padis populations, being associated with significantly lower IC50 values than the other populations.
Amid the conditions of 2019, the bilberry populations recorded IC50 values from 0.037 mg/mL in Semenic and Padis to 0.076 mg/mL in Sadova Noua, with an amplitude of 0.039 mg/mL, higher than the previous year. This year, the Semenic, Padis, Muntele Mic, and Raul Lung populations showed the highest antioxidant activity, associated with significantly lower IC50 values than the Sadova Noua and Cuntu populations.
In the conditions of 2020, the IC50 value had lower variability between populations than in previous years, with values from 0.03 mg/mL in Semenic to 0.049 mg/mL in Padis, on a background of lower antioxidant activity compared to the results of 2018–2019. The Semenic population had a higher antioxidant activity and a significantly lower IC50 than the Padis, Raul Lung, and Sadova Noua populations.
In the case of the Muntele Mic and Raul Lung populations, the IC50 values had small and no significant variations during the study. In the Sadova Noua population, significant variations of antioxidant activity from one year to another were observed, associated with higher IC50 values in 2019 and lower in 2020. The Semenic and Cuntu populations registered significantly higher antioxidant activities in 2020 than in 2018, while in the Padis population, the IC50 value in 2019 was significantly lower than that of 2020.
Depending on the values of the ASV index (
Table 10), it was noted that the Semenic and Muntele Mic populations had the highest stability of IC
50, on a background of a quantitative (same signs of IPCA) type of population–year interaction. In the case of the Sadova Noua population, the IC
50 value recorded large deviations during the study, associated with a qualitative interaction.
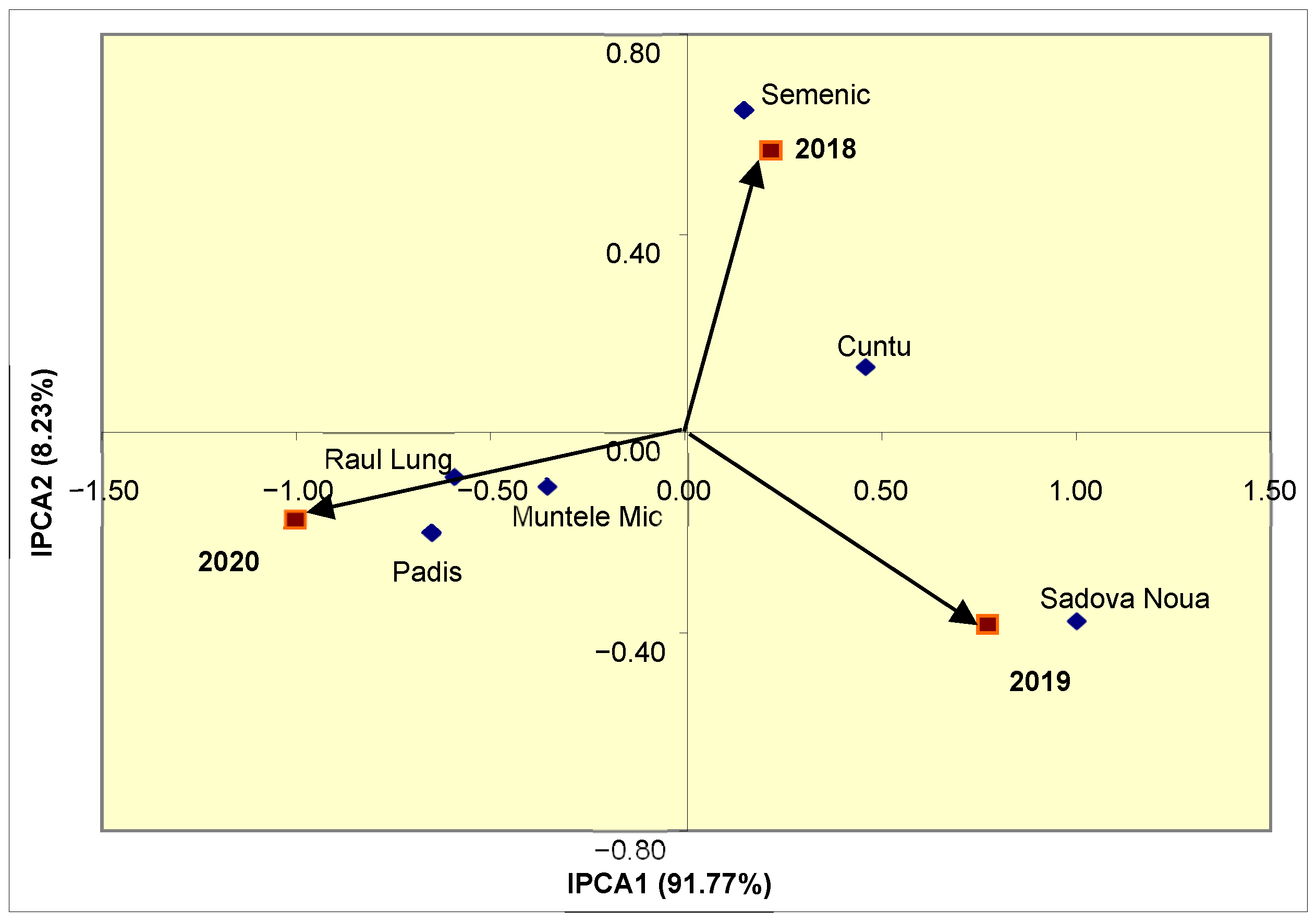
Considering that IPCA1 has a considerable contribution (91.77%) to total variability, based on the position in the biplot from
Figure 3, it was noted that the Semenic and Muntele Mic populations expressed the highest stability of IC
50 during the study. Based on the year’s associated vectors, it was found that the conditions from 2018–2019 had close contributions to the population–year interaction. Depending on the position towards the year vectors, it was observed that the Sadova Noua population had the lowest stability of IC
50 amid specific adaptation to the conditions of 2019. In the case of the Padis and Raul Lung populations, specific adaptations to the conditions of 2020 were observed.
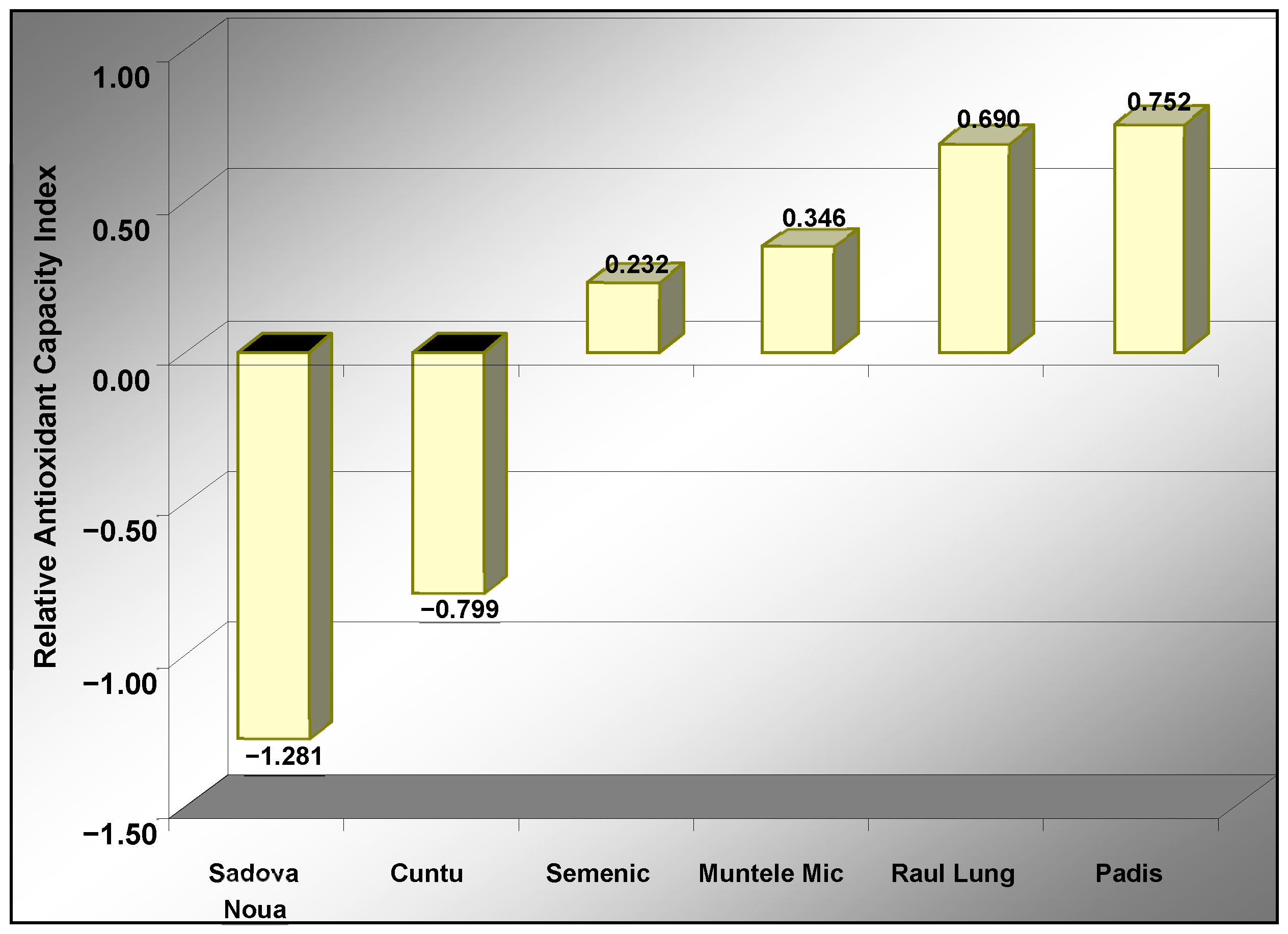
Based on the relative antioxidant capacity index (RACI) value of the six bilberry populations (
Figure 4), it was noted that the Padis and Raul Lung populations showed the highest antioxidant activity, while the Sadova Noua and Cuntu populations had significantly lower values of this index and relatively week antioxidant activity, respectively.
3.4. Variance Components and Correlations
In the case of polyphenol content (
Table 11), major differences were observed among phenotypic and genotypic variances, and among the two coefficients of variation, respectively. This fact indicates that the phenotypic expression of this trait has been considerably influenced by the population–year interaction. The low value of broad sense heritability (14.83%), associated with the fact that the genotypic variance was significantly lower than other variances, suggests that in these six bilberry populations, the polyphenol content is mainly controlled by environmental conditions and non-additive genes.
In the case of radical scavenging activity on the background of variation coefficients that attest a very low variability, it was noted that genotypic variance has a low contribution to phenotypic variance, having at the same time a significantly lower value compared to the interaction variance. Considering the low value of broad sense heritability (24.89%), it turns out that this trait is strongly influenced by non-additive genes in interaction with environmental conditions.
In the case of IC50 on the background of close values for the coefficients of variation, it was noted that phenotypic variance has a similar contribution as the interaction variance to the total variability. Considering the heritability value (56.50%) as well, it can be stated that in the inheritance of this trait, genes with additive effect also act.
Considering the correlations between the analyzed traits of bilberry fruits and environmental characteristics, it was found that the TPC had a positive relationship with soil pH and phosphorus content, and weak correlations with soil nitrogen and potassium contents (
Table 12). The DPPH radical scavenging activity had a moderate positive correlation with pH and phosphorus content, and negative correlations with nitrogen and potassium contents in soil. IC
50 values are significantly negatively correlated with altitude and positively correlated with potassium content, suggesting that the inhibitory concentration of fruit extracts decreases with increasing altitude and decreasing potassium content.



 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}