
Selection of Elite dura-Type Parents to Produce Dwarf Progenies of Elaeis guineensis Using Genetic Parameters


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Data Collection
2.3. Statistical Analysis
3. Results and Discussion
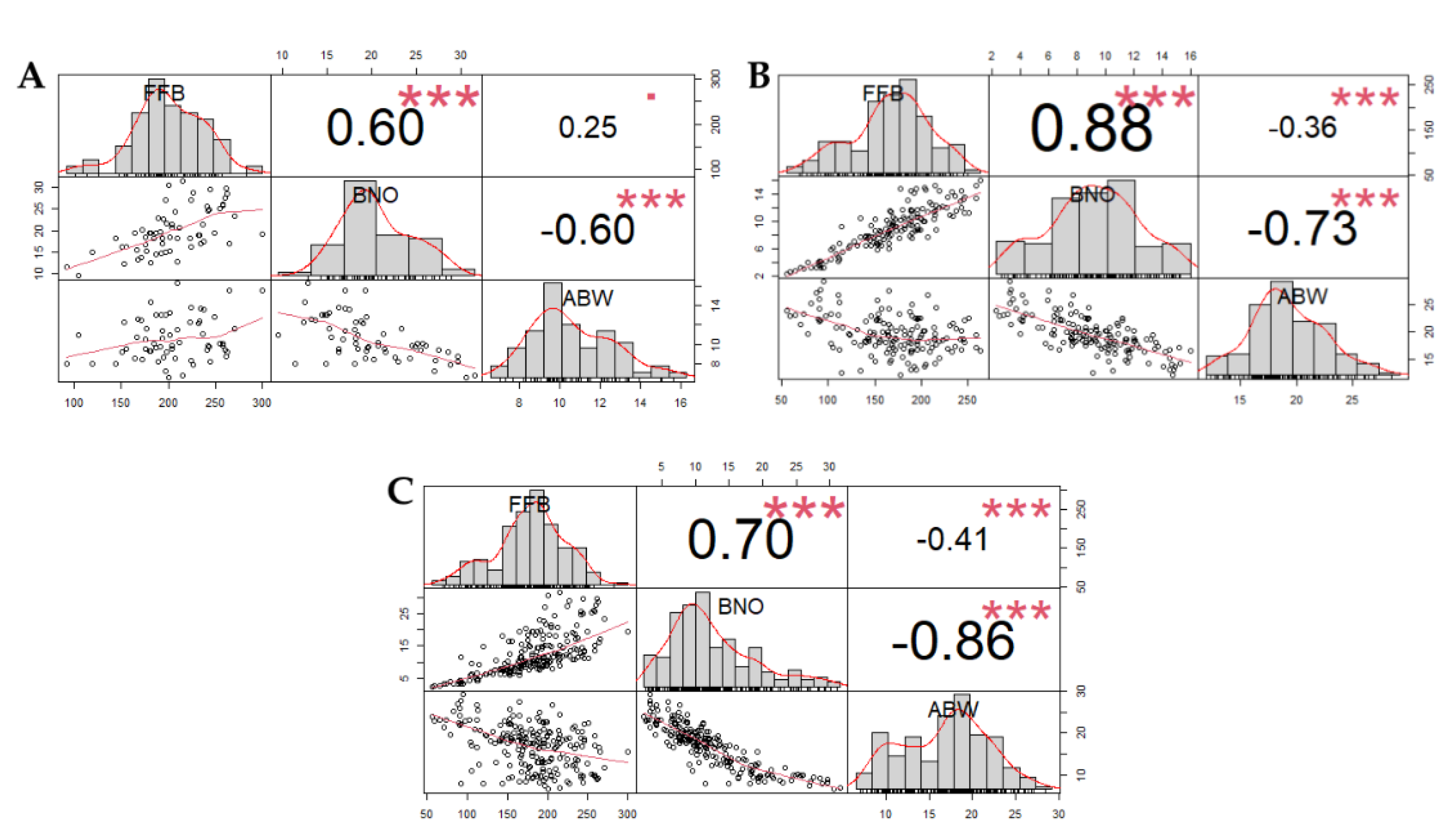
3.1. Yield Component Traits
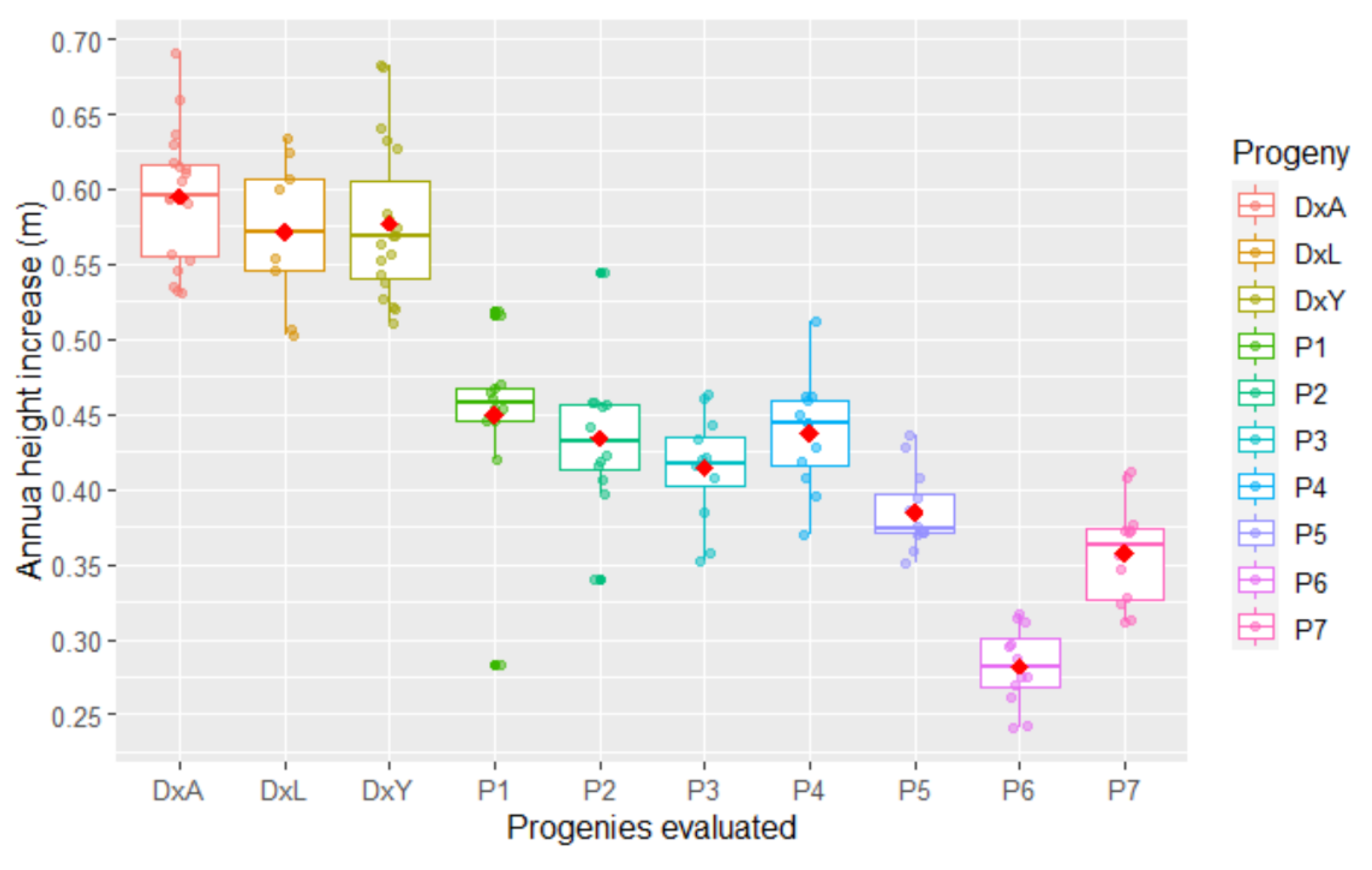
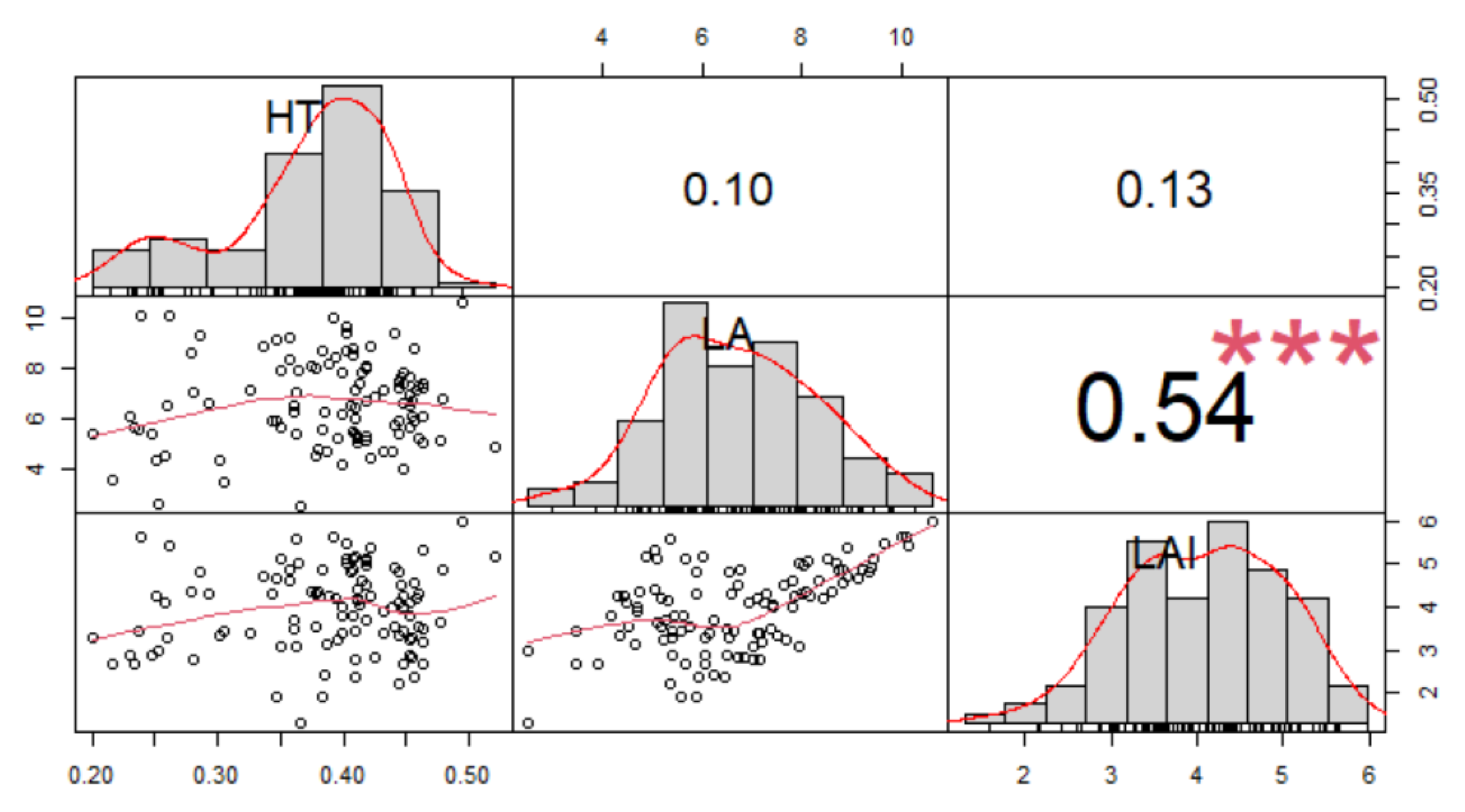
3.2. Vegetative Traits
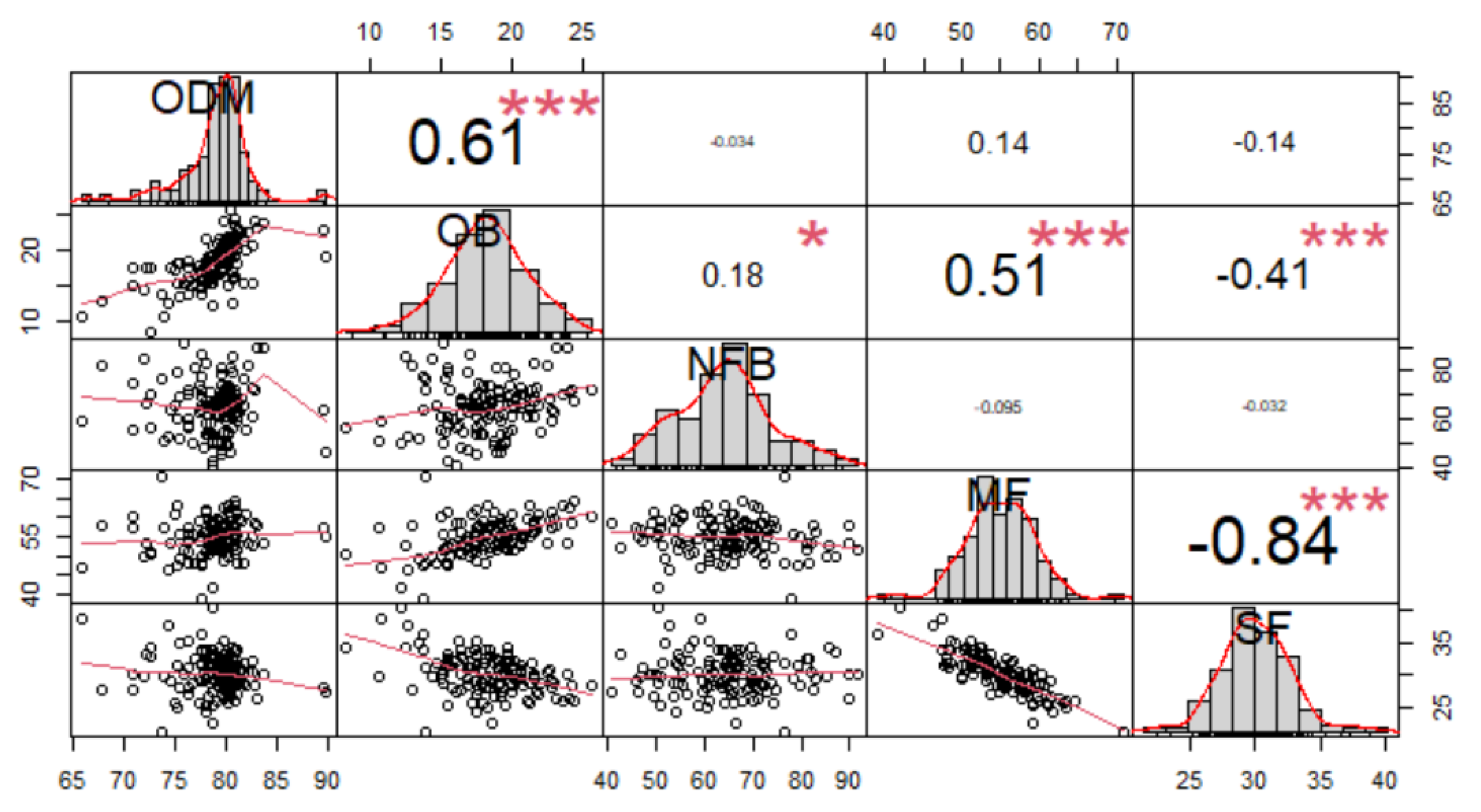
3.3. Bunch Quality Traits
3.4. Heritability and Genetic Parameters
3.5. Analysis of Agronomic Traits
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fedepalma. Anuario estadístico 2020. Principales cifras de la agroindustria de la palma de aceite en Colombia y en el mundo; Fedepalma: Bogotá, Colombia, 2020; p. 238. [Google Scholar]
- Tabi, K.M.A. Effect of dry heat treatment along with some dormancy breaking chemicals on oil palm seed germination. S. Afr. J. Bot. 2017, 112, 489–493. [Google Scholar] [CrossRef]
- Beirnaert, A.D.F.; Vanderweyen, R. Contribution À L’étude Génétique et Biométrique des Variétés D’ Elaeis Guineensis Jacquin; East African Standard: Nairobi, Kenya, 1941; p. 27. [Google Scholar]
- Reyes, P.A.; Ochoa, J.C.; Montoya, C.; Daza, E.; Ayala, I.M.; Romero, H.M. Development and validation of a bi-directional allele-specific PCR tool for differentiation in nurseries of dura, tenera and pisifera oil palms. Agron. Colomb. 2015, 33, 5–10. [Google Scholar] [CrossRef]
- Singh, R.; Low, E.-T.L.; Ooi, L.C.-L.; Ong-Abdullah, M.; Ting, N.-C.; Nagappan, J.; Nookiah, R.; Amiruddin, M.D.; Rosli, R.; Manaf, M.A.A. The oil palm SHELL gene controls oil yield and encodes a homologue of SEEDSTICK. Nature 2013, 500, 340–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, B.K.; Mathur, R.; Kumar, P.N.; Ramajayam, D.; Ravichandran, G.; Venu, M.; Babu, S.S. Development, identification and validation of CAPS marker for SHELL trait which governs dura, pisifera and tenera fruit forms in oil palm (Elaeis guineensis Jacq.). PLoS ONE 2017, 12, e0171933. [Google Scholar] [CrossRef] [Green Version]
- Mondragon-Serna, A.; Baena-Snata, M.A.; González-Díaz, A.; García-Nuñez, J.A.; Ayala-Diaz, I.M.; Romero, H.M. The Oil Palm. In Oil Crops. Growth, Uses, and Toxicity; Perea-Flores, M.J., Camacho-Díaz, B.H., Quintanilla-Carvajal, M.X., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2021; pp. 105–146. [Google Scholar]
- Rajanaidu, N. A Review of Oil Palm Breeding for the Past 50 Years. Rev. Palmas 2016, 37, 190–202. [Google Scholar]
- Noh, A.; Rafii, M.Y.a. Variability and performance evaluation of introgressed Nigerian dura x Deli dura oil palm progenies. Genet. Mol. Res. 2014, 13, 2426–2437. [Google Scholar] [CrossRef]
- Bastidas, S. Genealogíaa del germoplasma de palma de aceite (Elaeis guineensis Jacq.) del proyecto de mejoramiento genético de Corpoica. Rev. Palmas 2003, 24, 21–29. [Google Scholar]
- Ishak, Z.; Hashim, A.T.; Rosli, S.K.; Bakar, D.A.; Ooi, S.E.; Mohd, N.; Ong-Abdullah, M. Oil Palm Tissue Culture: Fast Tracking Elite Commercial Lines. In The Oil Palm Genome; Springer: Berlin/Heidelberg, Germany, 2020; pp. 47–68. [Google Scholar]
- Domiciano, G.P.; Alves, A.A.; Laviola, B.G.; da Conceio, L.D.H.C.S. Parámetros genéticos e diversidade em progenies de macauba com base em características morfológicas e fisiológicas. Cienc. Rural 2015, 45, 1599–1605. [Google Scholar] [CrossRef] [Green Version]
- Hefena, A.; Sultan, M.; Abdel-Moneam, M.; Hammoud, S.; Barutular, C.; El-Sabagh, A. Assessment of Genetic Variability and Correlation Coefficient to Improve Some Agronomic Traits in Rice. J. Exp. Agric. Int. 2016, 14, 1–8. [Google Scholar] [CrossRef]
- Machado, E.L.; Silva, S.A.; Fernandes, L.d.S.; Brasileiro, H.S. Genetic variability and homozygosity in a F4 castor bean population by microsatellite markers. Bragantia 2016, 75, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Shafique, M.S.; Ahsan, M.; Mehmood, Z. Genetic variability and interrelationship of various agronomic traits using correlation and path analysis in chickpea (Cicer arietinum L.). Acad. J. Agric. Res. 2016, 4, 82–85. [Google Scholar] [CrossRef]
- Ayala, I.; Romero, H.M.; Tupaz, A.; Daza, E.; Rincón, A.; Caicedo, A.; Fontanilla, C.; Mosquera, M. Comportamiento Agronómico de Cultivares Comerciales de Palma de Aceite en el Campo Experimental Palmar de la Vizcaína; Cenipalma: Bogotá, Colombia, 2017; p. 138. [Google Scholar]
- Corley, R.H.V.; Breure, C.J. Measurements in Oil Palm Experiments; Internal Report; Unilever Plantations: London, UK, 1981. [Google Scholar]
- Hardon, J.J.; Williams, C.N.; Watson, I. Leaf area and yield in the oil palm in malaya. Exp. Agric. 1969, 5, 25–32. [Google Scholar] [CrossRef]
- Corley, R.H.V.; Tinker, P.B. The Oil Palm, 5th ed.; Wiley Blackwell: Oxford, UK, 2016; p. 650. [Google Scholar]
- Blaak, G.; Sparnaaij, L.; Menedez, T. Breeding and inheritance in the oil palm (Elaeis guineensis Jacq.) II. Methods of bunch quality analysis. J.W. Afr. Inst. Oil Palm Res. 1963, 4, 146–155. [Google Scholar]
- Prada, F.; Romero, H.M. Muestreo y Analisis de Racimos en el Cultivo de la Palma de Aceite. Tecnologías para la Agroindustria de la Palma de Aceite, Guia de Facilitadores; Cenipalma: Bogotá, Colombia, 2012; p. 158. [Google Scholar]
- Falconer, D.; Mackay, T. Introduction to Quantitative Genetics; Longman Group: Essex, UK, 1996. [Google Scholar]
- Arolu, I.W.; Rafii, M.Y.; Marjuni, M.; Hanafi, M.M.; Sulaiman, Z.; Rahim, H.A.; Abidin, M.I.Z.; Amiruddin, M.D.; Din, A.K.; Nookiah, R. Breeding of high yielding and dwarf oil palm planting materials using Deli dura × Nigerian pisifera population. Euphytica 2017, 213, 154. [Google Scholar] [CrossRef]
- Sapey, E.; Peprah, B.B.; Adusei-Fosu, K.; Agyei-Dwarko, D. Genetic Variability of Fresh Fruit Bunch Yield (FFB) Yield in Some Dura × Pisifera Breeding Populations of Oil Palm (Elaeis guineensis Jacq.). Am. -Eurasian J. Agric. Environ. Sci. 2015, 15, 1637–1640. [Google Scholar] [CrossRef]
- Noh, A.; Rafii, M.Y.; Saleh, G.; Kushairi, A.; Latif, M.A. Genetic performance and general combining ability of oil palm Deli dura × AVROS pisifera tested on inland soils. Sci. World J. 2012, 2012, 792601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaray, S.; Amiruddin, M.D.; Rafii, M.Y.; Jamian, S.; Ismail, M.F.; Jalloh, M.; Marjuni, M.; Mohamad, M.M.; Yusuff, O. Influence of parental dura and pisifera genetic origins on oil palm fruit set ratio and yield components in their D × P Progenies. Agronomy 2020, 10, 1793. [Google Scholar] [CrossRef]
- Myint, K.A.; Amiruddin, M.D.; Rafii, M.Y.; Samad, M.Y.A.; Ramlee, S.I.; Yaakub, Z.; Oladosu, Y. Genetic diversity and selection criteria of MPOB-Senegal oil palm (Elaeis guineensis Jacq.) germplasm by quantitative traits. Ind. Crop. Prod. 2019, 139, 111558. [Google Scholar] [CrossRef]
- Alvarado, A.; Henry, J. Evolution blue: A new oil palm variety with reduced growth and high oil content. A.S.D. Oil Palm Pap. 2015, 45, 45. [Google Scholar]
- Saldaña-Villota, T.M.; Cotes-Torres, J.M. Radiation interception and leaf area index from foliage cover in diploid potato. Agron. J. 2020, 112, 2805–2811. [Google Scholar] [CrossRef]
- Breure, C.J. Rate of leaf expansion: A criterion for identifying oil palm (Elaeis guineensis Jacq.) types suitable for planting at high densities. NJAS—Wagening. J. Life Sci. 2010, 57, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Barcelos, E. Oil palm natural diversity and the potential for yield improvement. Front. Plant Sci. 2015, 6, 190. [Google Scholar] [CrossRef]
- Alvarado, A.; Chinchilla, C.; Rodríguez, J. Desempeño de dos variedades de palma aceitera (Deli × AVROS y Deli × Ghana) plantadas a diferentes densidades en dos sitios en Costa Rica. ASD Oil Palm Pap. 2007, 8, 35–41. [Google Scholar]
- Sunilkumar, K.; Mathur, R.K.; Sparjanbabu, D.S.; Pillai, R.S.N. Evaluation of interspecific oil palm hybrids for dwarfness. J. Plant. Crop. 2015, 43, 29–34. [Google Scholar]
- Donough, C.R. Breeding Oil Palms for High Oil Yield in I.O.I. Group: 1. First Cycle Development of OPGL-Derived Materials; MPOB National Seminar: Kuala Lumpur, Malaysia, 2005; p. 125. [Google Scholar]
- Marhalil, M.; Zulkifli, Y.; Kushairi, A.; Adura, Z.S.; Nurniwalis, A.; Zubaidah, R. Potential oil palm genetic materials derived from introgression of germplasm (MPOB-Nigeria, MPOB-Zaire and MPOB-Cameroon accessions) to advanced (AVROS) breeding populations. J. Oil Palm Res. 2020, 32, 569–581. [Google Scholar]
- Murugesan, P.; Rani, K.L.M.; Ramajayam, D.; Kumar, K.S.; Mathur, R.K.; Ravichandran, G.; Kumar, P.N.; Arunachalam, V. Genetic diversity of vegetative and bunch traits of African oil palm (Elaeis guineensis) germplasm in India. Indian J. Agric. Sci. 2015, 85, 892–895. [Google Scholar]
- Ortega Cedillo, D.; Barrera, C.F.; Ortega Cedillo, J.; Orellana Carrera, J.; Vilela de Resende, M.D.; Damião Cruz, C. Estimates of parameters, prediction and selection of an oil palm population in Ecuador. Rev. Fac. Nac. Agron. Medellín 2018, 71, 8477–8487. [Google Scholar] [CrossRef] [Green Version]
- De Almeida Rios, S.; da Cunha, R.N.V.; Lopes, R.; Barcelos, E.; da Rocha, R.N.C.; de Lima, W.A.A. Correlation and Path analysis for yield components in dura oil palm germplasm. Ind. Crop. Prod. 2018, 112, 724–733. [Google Scholar] [CrossRef]
- Cedillo, D.S.O.; Barros, W.S.; Ferreira, F.M.; Dias, L.A.D.S.; Rocha, R.B.; Cruz, C.D. Correlation and repeatability in progenies of African oil palm. Acta Sci. Agron. 2008, 30, 197–201. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scheme | df | MS | EMS |
|---|---|---|---|
| Replicates (R) | (r−1) | MS1 | σ2 e + σ2 ga + σ2 r |
| Progenies (G) | (g−1) | MS2 | σ2 e + σ2 ga + σ2 g |
| Year (E) | (a−1) | MS3 | σ2 e + σ2 ga + σ2 a |
| G × E | (g−1) (a−1) | MS4 | σ2 e + σ2 ga |
| Error | σ2 e |
| Source of Variation | df | FFB | BNO | ABW |
|---|---|---|---|---|
| Replicates (R) | 2 | 5329.40 | 15.29 | 4.72 |
| Progenies (G) | 6 | 7739.85 ** | 47.99 ** | 50.45 ** |
| Year (E) | 9 | 29,154.64 ** | 782.39 ** | 533.39 ** |
| G × E | 54 | 808.50 ns | 10.54 ** | 3.00 ** |
| Error | 138 | 854.14 | 3.66 | 2.02 |
| Variance components | ||||
| σ2 G | 231.05 (21.5) § | 1.25 (18.2) | 1.58 (40.7) | |
| σ2 G.E. | −13.04 (−1.2) | 1.96 (28.6) | 0.28 (7.2) | |
| σ2 e | 854.14 (79.7) | 3.66 (53.3) | 2.02 (52.0) |
| Progenies | FFB | BN | ABW |
|---|---|---|---|
| P1 | 192.62 b | 11.52 bc | 18.75 a |
| P2 | 173.07 cd | 11.75 bc | 16.71 b |
| P3 | 208.44 a | 13.99 a | 17.40 b |
| P4 | 165.02 d | 12.17 b | 15.30 c |
| P5 | 182.50 bc | 14.25 a | 15.86 c |
| P6 | 169.34 cd | 11.04 c | 18.15 a |
| P7 | 166.27 cd | 11.64 bc | 15.81 c |
| Trial mean | 179.61 | 12.34 | 16.85 |
| CV (%) | 8.94 | 10.25 | 7.69 |
| Years after Planting (Y.A.P.) | |||
| Y.A.P. 3 | 185.07 c | 20.80 b | 9.15 g |
| Y.A.P. 4 | 221.70 a | 23.28 a | 9.55 g |
| Y.A.P. 5 | 206.65 ab | 15.87 c | 13.02 f |
| Y.A.P. 6 | 193.49 bc | 13.13 d | 14.72 e |
| Y.A.P. 7 | 175.87 c | 9.05 f | 19.51 c |
| Y.A.P. 10 | 207.77 ab | 11.76 e | 17.71 d |
| Y.A.P. 11 | 176.01 c | 8.95 f | 19.76 c |
| Y.A.P. 12 | 186.44 c | 9.67 f | 19.37 c |
| Y.A.P. 13 | 154.39 d | 7.07 g | 21.96 b |
| Y.A.P. 14 | 88.69 e | 3.78 h | 23.78 a |
| Young stage mean (A.Y.P. 3–5) | 204.47 | 19.98 | 10.57 |
| CV (%) | 9.00 | 18.88 | 20.13 |
| Adult stage mean (Y.A.P. 6–14) | 168.95 | 9.05 | 19.54 |
| CV (%) | 23.13 | 33.68 | 14.84 |
| Trial mean | 179.61 | 12.34 | 16.85 |
| CV (%) | 20.75 | 49.48 | 29.90 |
| Source of Variation | df | FP | PCS | RL | LL | LW | LN | HT | LA | LAI | DI |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Replicates (R) | 2 | 50.280 | 0.100 | 0.020 | 119.870 | 0.410 | 35.200 | 0.019 | 0.656 | 0.495 | 0.000 |
| Progenie (G) | 6 | 91.480 ** | 0.200 * | 0.640 ** | 520.360 ** | 3.080 * | 410.760 ** | 0.061 ** | 2.200 ** | 1.213 ** | 0.005 ns |
| Year (E) | 4 | 3269.880 ** | 102.300 ** | 7.610 ** | 27,410.430 ** | 154.850 ** | 3276.130 ** | 0.003 ns | 58.512 ** | 13.218 ** | 0.347 ** |
| G × E | 24 | 22.720 ns | 0.060 ns | 0.060 ns | 75.260 * | 0.840 ** | 29.610 ns | 0.002 ns | 0.454 ns | 0.300 ns | 0.003 ** |
| Error | 68 | 23.880 | 0.070 | 0.050 | 44.380 | 0.180 | 28.860 | 0.002 | 0.284 | 0.252 | 0.000 |
| Variance components | |||||||||||
| σ2 G | 4.584 (16.3) § | 0.009 (12.2) | 0.039 (42.2) | 29.673 (35.8) | 0.149 (28.8) | 25.410 (46.6) | 0.004 (71.6) | 0.116 (25.9) | 0.061 (18.7) | 0.000 (12.6) | |
| σ2 G.E. | −0.331 (−1.2) | −0.003 (−3.7) | 0.003 (3.1) | 8.823 (10.6) | 0.189 (36.4) | 0.214 (0.4) | 0.000 (0.7) | 0.049 (10.8) | 0.014 (4.3) | 0.001 (55.0) | |
| σ2 e | 23.880 (84.9) | 0.070 (91.5) | 0.050 (54.6) | 44.380 (53.5) | 0.180 (34.8) | 28.860 (53.0) | 0.002 (27.7) | 0.284 (63.3) | 0.252 (77.1) | 0.000 (32.4) |
| Progenies | FP | PCS | RL | LL | LW | LN | HT | LA | LAI | DI |
|---|---|---|---|---|---|---|---|---|---|---|
| P1 | 46.67 a | 24.16 ab | 4.68 a | 93.78 a | 8.71 a | 157.71 a | 0.47 a | 7.35 a | 5.49 a | 0.33 a |
| P2 | 45.90 ab | 23.87 ab | 4.34 b | 75.09 c | 7.62 c | 150.49 b | 0.43 ab | 6.59 cd | 5.09 ab | 0.33 a |
| P3 | 41.33 ab | 26.32 a | 4.76 a | 85.46 b | 8.57 a | 156.98 a | 0.42 b | 7.19 ab | 5.17 ab | 0.30 bc |
| P4 | 46.21 a | 22.68 b | 4.28 b | 78.68 bc | 7.56 c | 144.08 c | 0.43 ab | 6.40 d | 5.12 b | 0.32 ab |
| P5 | 43.06 ab | 24.01 ab | 4.68 a | 82.54 b | 7.86 bc | 157.38 a | 0.39 b | 6.54 d | 4.72 b | 0.29 c |
| P6 | 42.33 ab | 24.81 ab | 4.69 a | 84.57 b | 8.28 ab | 158.42 a | 0.29 d | 7.17 abc | 5.43 ab | 0.29 c |
| P7 | 40.74 b | 23.50 ab | 4.34 b | 81.55 bc | 7.92 bc | 154.01 ab | 0.33 c | 6.63 bcd | 5.09 b | 0.30 c |
| Trial mean | 43.75 | 24.19 | 4.54 | 83.09 | 8.07 | 154.15 | 0.39 | 6.84 | 5.16 | 0.31 |
| CV (%) | 5.64 | 4.72 | 4.56 | 7.09 | 5.61 | 3.39 | 16.14 | 5.60 | 4.93 | 5.65 |
| Years after planting (Y.A.P.) | ||||||||||
| YAP 4 | 63.05 a | 9.96 d | 4.28 c | 103.60 b | 5.62 d | 135.00 d | 0.38 a | 4.76 e | 4.30 b | 0.25 a |
| YAP 6 | 48.04 b | 11.84 c | 4.77 b | 114.74 a | 6.12 c | 150.28 c | 0.40 a | 5.91 d | 4.02 b | 0.26 a |
| YAP 8 | 30.43 d | 40.54 b | 3.60 d | 47.65 c | 11.10 a | 157.23 b | 0.41 a | 6.76 c | 2.94 c | 0.25 a |
| YAP 10 | 37.68 c | 12.39 c | 4.96 ab | 109.41 a | 6.58 b | 160.03 b | 0.40 a | 7.61 b | 4.06 b | 0.25 a |
| Y.A.P. 14 | 39.53 c | 65.66 a | 5.06 a | 40.07 d | 10.96 a | 168.23 a | 0.39 a | 9.16 a | 5.16 a | N.A. |
| Trial mean | 43.75 | 28.08 | 4.54 | 83.09 | 8.08 | 154.15 | 0.40 | 6.84 | 4.10 | 0.25 |
| CV (%) | 28.53 | 87.34 | 13.27 | 43.48 | 33.66 | 8.10 | 2.88 | 24.43 | 19.37 | 1.98 |
| Source of Variation | df | MFW | SB | ODM | OFM | OB | NFB | FS | MF | K/F | S/F | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Replicates (R) | 2 | 5.51 | 3.6 | 15.52 | 60.32 | 18.69 | 88.1 | 45.63 | 18.56 | 2.53 | 9.05 | |
| Progenies (G) | 6 | 11.62 ** | 21.21 ** | 5.86 ns | 68.96 ** | 20.14 * | 158.12 ns | 83.51 ns | 114.52 ** | 47.48 ** | 32.74 ** | |
| Year (E) | 5 | 42.24 ** | 9.00 ** | 76.46 ** | 94.4 ** | 61.26 ** | 1037.93 ** | 1782.49 ** | 98.99 ** | 51.7 ** | 50.32 ** | |
| G × E | 30 | 1.47 ns | 1.90 * | 7.54 ns | 11.78 ns | 6.01 ns | 85.18 ns | 44.93 ns | 11.83 ns | 3.1 ns | 5.67 ns | |
| Error | 82 | 1.99 | 1.04 | 4.84 | 11.44 | 5.79 | 58.35 | 52.44 | 11.88 | 3.07 | 5.22 | |
| σ2 G | 0.56 (23.44) § | 1.07 (45.49) | 0.09 (−1.69) | 3.18 (21.59) | 0.79 (11.83) | 4.05 (5.78) | 2.14 (4.09) | 5.71 (32.47) | 2.47 (44.47) | 1.50 (21.95) | ||
| σ2 GE | −0.15 (−6.18) | 0.25 (10.42) | 0.77 (13.98) | 0.10 (0.66) | 0.06 (0.95) | 7.67 (10.94) | −2.15 (−4.09) | −0.01 (−0.08) | 0.01 (0.15) | 0.13 (1.88) | ||
| σ2 e | 1.99 (82.73) | 1.04 (44.10) | 4.84 (87.71) | 11.44 (77.75) | 5.79 (87.23) | 58.35 (83.28) | 52.44 (100) | 11.88 (67.61) | 3.07 (55.37) | 5.22 (76.18) |
| Progenies | MFW | SB | ODM | OFM | OB | NFB | FS | MF | KF | SF |
|---|---|---|---|---|---|---|---|---|---|---|
| P1 | 10.42 ab | 90.54 bc | 78.77 a | 51.19 b | 18.51 a | 67.46 a | 74.09 a | 57.30 ab | 13.81 bc | 28.89 b |
| P2 | 10.94 a | 91.81 a | 78.42 a | 53.32 ab | 18.96 a | 65.86 a | 76.35 a | 56.04 abc | 14.82 b | 29.13 b |
| P3 | 10.08 ab | 89.78 cd | 78.38 a | 52.03 ab | 16.84 a | 61.03 a | 69.98 a | 52.89 cd | 14.68 b | 32.42 a |
| P4 | 9.67 abc | 91.49 ab | 78.00 a | 55.07 a | 19.02 a | 66.39 a | 74.71 a | 53.97 bcd | 15.27 b | 30.75 ab |
| P5 | 8.53 c | 88.97 d | 79.62 a | 55.12 a | 18.98 a | 68.67 a | 72.44 a | 54.55 bcd | 15.18 b | 30.27 ab |
| P6 | 10.29 ab | 89.50 d | 78.29 a | 50.68 b | 16.61 a | 65.34 a | 71.92 a | 51.87 d | 17.82 a | 30.31 ab |
| P7 | 9.25 bc | 91.23 ab | 79.21 a | 55.21 a | 18.84 a | 61.17 a | 71.60 a | 59.04 a | 12.48 c | 28.48 b |
| Trial mean | 9.88 | 90.48 | 78.67 | 53.23 | 18.25 | 65.13 | 73.01 | 55.10 | 14.87 | 30.04 |
| CV (%) | 8.13 | 1.20 | 0.73 | 3.68 | 5.80 | 4.55 | 2.95 | 4.58 | 10.92 | 4.49 |
| Years after planting (Y.A.P.) | ||||||||||
| YAP 3 | 8.70 d | 91.49 a | 79.55 a | 51.34 c | 17.93 b | 74.88 a | 87.13 a | 53.28 c | 16.6 a | 30.12 a |
| YAP 5 | 9.18 cd | 90.85 ab | 74.94 b | 51.47 bc | 15.61 c | 68.51 ab | 79.72 b | 52.95 c | 15.54 ab | 31.51 a |
| YAP 7 | 11.85 a | 89.95 bc | 79.48 a | 54.48 ab | 17.82 b | 54.93 d | 62.77 e | 54.10 c | 14.93 b | 30.98 a |
| YAP 9 | 11.12 ab | 89.80 c | 80.25 a | 56.88 a | 20.86 a | 63.62 bc | 70.38 cd | 58.16 a | 11.96 c | 29.87 a |
| Y.A.P. 11 | 10.21 bc | 90.71 abc | 79.23 a | 52.89 bc | 18.96 ab | 68.51 ab | 73.32 bc | 54.71 bc | 14.66 b | 30.63 a |
| YAP 14 | 8.25 d | 90.05 bc | 78.57 a | 52.32 bc | 18.31 b | 60.33 cd | 64.75 de | 57.37 ab | 15.52 ab | 27.11 b |
| Trial mean | 9.89 | 90.48 | 78.67 | 53.23 | 18.25 | 65.13 | 73.01 | 55.10 | 14.87 | 30.04 |
| CV (%) | 14.35 | 0.72 | 2.43 | 3.98 | 9.36 | 10.79 | 12.62 | 3.94 | 10.55 | 5.15 |
| Trait | σ2g | σ2ge | σ2e | σ2p | H2B (%) |
|---|---|---|---|---|---|
| FFB | 231.0452 | −13.0404 | 854.1390 | 1072.14 | 21.55 |
| BNO | 1.2485 | 1.9637 | 3.6625 | 6.87 | 18.16 |
| ABW | 1.5816 | 0.2805 | 2.0212 | 3.88 | 40.73 |
| FP | 4.5840 | -0.3314 | 23.8800 | 28.13 | 16.29 |
| PCS | 0.0093 | −0.0029 | 0.0700 | 0.08 | 12.20 |
| RL | 0.0387 | 0.0029 | 0.0500 | 0.09 | 42.25 |
| LL | 29.6733 | 8.8229 | 44.3800 | 82.88 | 35.80 |
| LW | 0.1493 | 0.1886 | 0.1800 | 0.52 | 28.83 |
| LN | 25.4100 | 0.2143 | 28.8600 | 54.48 | 46.64 |
| HT | 0.0039 | 0.0000 | 0.0015 | 0.01 | 71.62 |
| LA | 0.1164 | 0.0486 | 0.2840 | 0.45 | 25.93 |
| LAI | 0.0609 | 0.0139 | 0.2516 | 0.33 | 18.65 |
| DI | 0.0001 | 0.0006 | 0.0004 | 0.00 | 12.60 |
| MFW | 0.5639 | −0.1486 | 1.9900 | 2.41 | 23.44 |
| SB | 1.0728 | 0.2457 | 1.0400 | 2.36 | 45.49 |
| ODM | 0.0933 | 0.7714 | 4.8400 | 5.52 | 1.69 |
| OFM | 3.1767 | 0.0971 | 11.4400 | 14.71 | 21.59 |
| OB | 0.7850 | 0.0629 | 5.7900 | 6.64 | 11.83 |
| NFB | 4.0522 | 7.6657 | 58.3500 | 70.07 | 5.78 |
| FS | 2.1433 | −2.1457 | 52.4400 | 52.44 | 4.09 |
| MF | 5.7050 | −0.0143 | 11.8800 | 17.57 | 32.47 |
| KF | 2.4656 | 0.0086 | 3.0700 | 5.54 | 44.47 |
| SF | 1.5039 | 0.1286 | 5.2200 | 6.85 | 21.95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tupaz-Vera, A.; Ayala-Diaz, I.; Barrera, C.F.; Romero, H.M. Selection of Elite dura-Type Parents to Produce Dwarf Progenies of Elaeis guineensis Using Genetic Parameters. Agronomy 2021, 11, 2581. https://doi.org/10.3390/agronomy11122581
Tupaz-Vera A, Ayala-Diaz I, Barrera CF, Romero HM. Selection of Elite dura-Type Parents to Produce Dwarf Progenies of Elaeis guineensis Using Genetic Parameters. Agronomy. 2021; 11(12):2581. https://doi.org/10.3390/agronomy11122581
Chicago/Turabian StyleTupaz-Vera, Andrés, Iván Ayala-Diaz, Carlos Felipe Barrera, and Hernán Mauricio Romero. 2021. "Selection of Elite dura-Type Parents to Produce Dwarf Progenies of Elaeis guineensis Using Genetic Parameters" Agronomy 11, no. 12: 2581. https://doi.org/10.3390/agronomy11122581
APA StyleTupaz-Vera, A., Ayala-Diaz, I., Barrera, C. F., & Romero, H. M. (2021). Selection of Elite dura-Type Parents to Produce Dwarf Progenies of Elaeis guineensis Using Genetic Parameters. Agronomy, 11(12), 2581. https://doi.org/10.3390/agronomy11122581