
Effect of Soil-Applied L-tryptophan on the Amount of Biomass and Nitrogen and Sulfur Utilization by Maize



Abstract
:1. Introduction
2. Materials and Methods
2.1. Growing Experiment
2.2. Analysis of Plant Material
2.3. Calculations and Statistical Analysis
2.4. Statistics
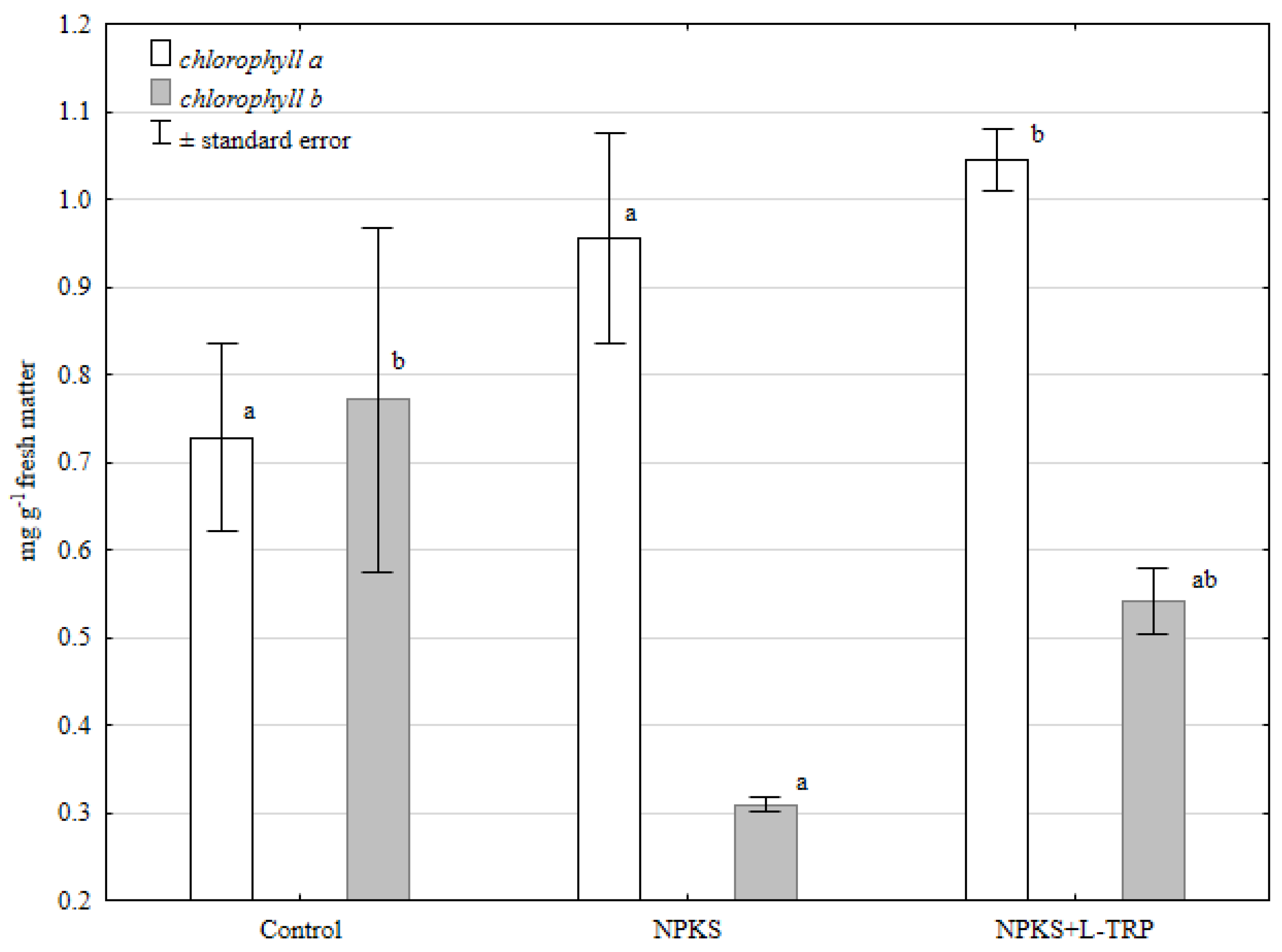
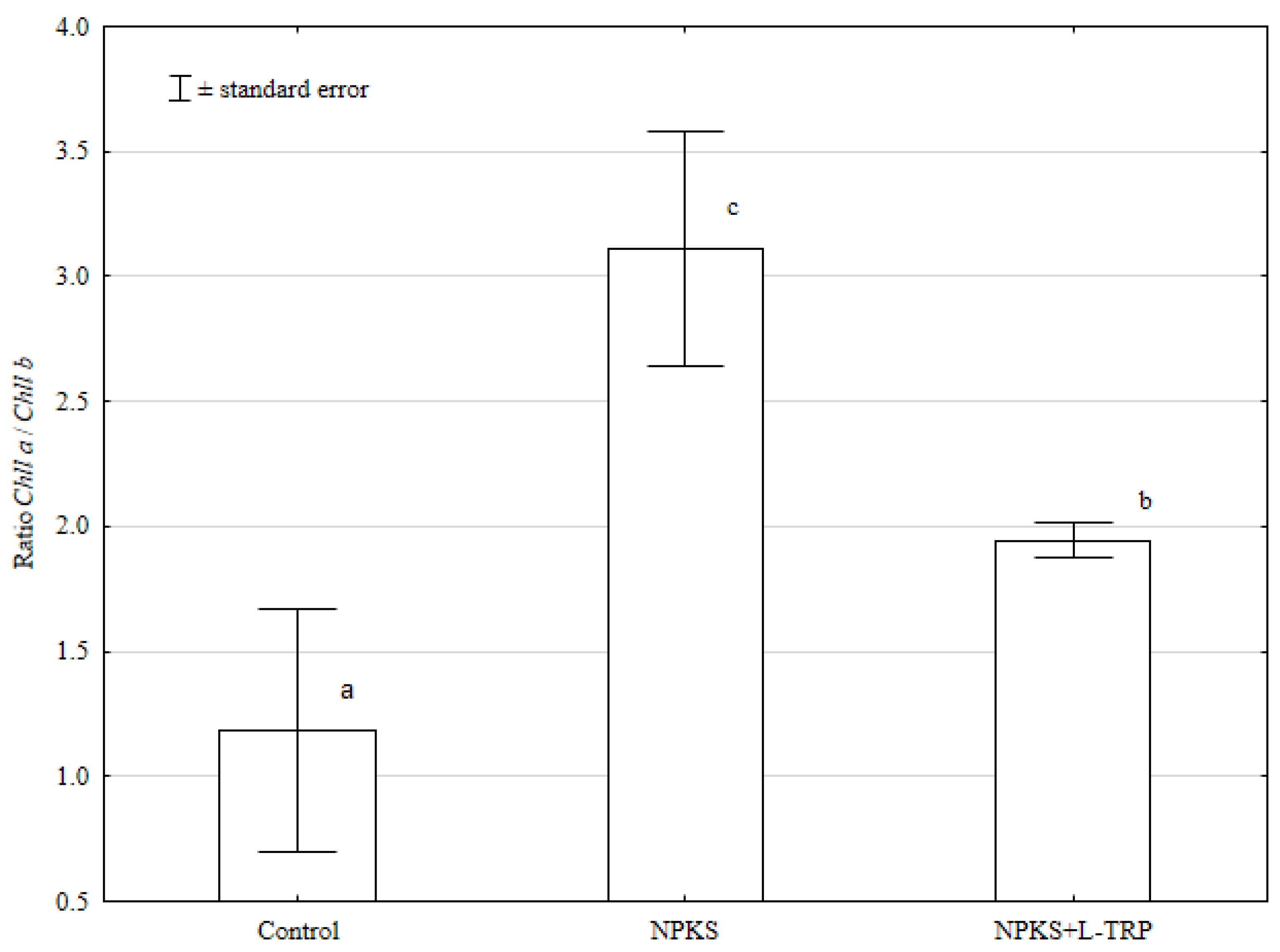
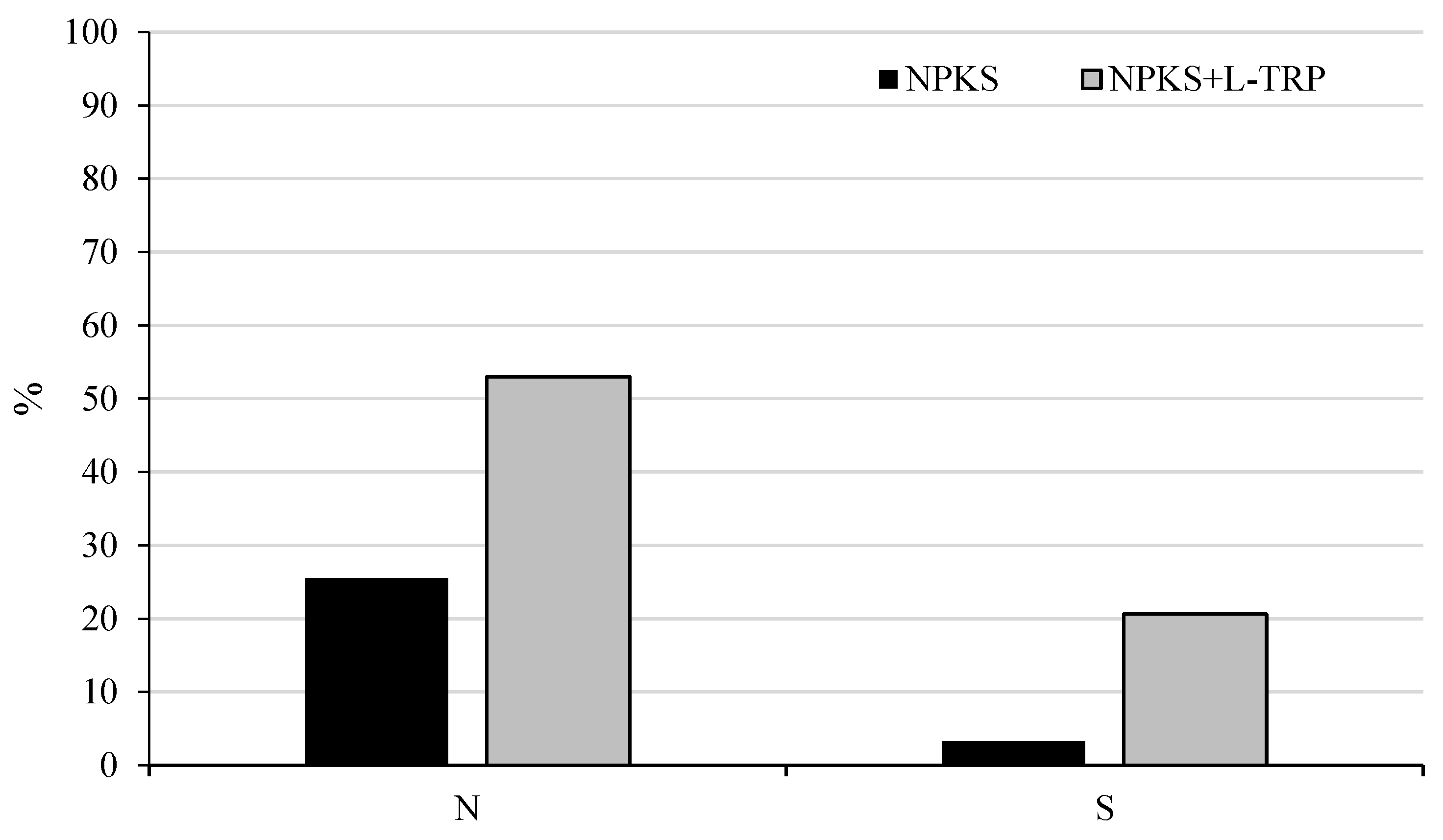
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Regulation of the Minister of Agriculture and Rural Development of 9 March 2015, Item 344, on the Issue of Norms in the Scope of Good Agricultural Culture in Accordance with Environmental Protection. Available online: http://isap.sejm.gov.pl/isap.nsf/download.xsp/WDU20150000344/O/D20150344.pdf (accessed on 16 December 2021). (In Polish)
- Pecio, A. Innovative products used in plant production. Stud. Rep. IUNG 2020, 63, 163–178. (In Polish) [Google Scholar]
- Regulation (EU) 2019/1009 of the European Parliament and of the Council of 5 June 2019 Laying down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repaling Regulation (EC) No 2003/2003. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32019R1009 (accessed on 16 December 2021).
- Wu, G. Amino acids: Metabolizm, functions, and nutrition. Amino Acids. 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Häusler, R.E.; Ludewig, F.; Krueger, S. Amino acids—A life between metabolism and signaling. Plant Sci. 2014, 229, 225–237. [Google Scholar] [CrossRef]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, T.M.; Nesi, A.N.; Araujo, W.L.; Braun, H.-P. Amono acid catabolism in plants. Molecul. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Hendawy, S.F.; El-Din, A.A.E. Growth and yield of Foeniculum vulgare var. Azoricum as influenced by some vitamins and amino AIDS. Ozean. J. Appl. Sci. 2010, 3, 113–123. [Google Scholar]
- Mustafa, A.; Imran, M.; Ashraf, M.; Mahmood, K. Perspectives of using L-tryptophan for improving productivity of agricultural crops: A review. Pedosphere 2018, 28, 16–34. [Google Scholar] [CrossRef]
- Hassan, T.U.; Bano, A. The stimulatory effects of L-tryptophan and plant growth promoting rhizobacteria (PGPR) on soil health and physiology of wheat. J. Soil Sci. Plant Nutr. 2015, 15, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, A.; Hussain, A.; Naveed, M.; Ditta, A.; Nazli, Z.E.H.; Sattar, A. Response of okra (Abelmoschus esculentus L.) to soil and foliar applied L-tryptophan. Soil Environ. 2016, 35, 76–84. [Google Scholar]
- Khan, S.; Yu, H.; Li, Q.; Gao, Y.; Sallam, B.N.; Wang, H.; Liu, P.; Jiang, W. Exogenous application of amino acids improves the growth and yield of lettuce by enhancing photosynthetic Assimilation and nutrient availability. Agronomy 2019, 9, 266. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.L.; Madrid, R.; Gimeno, V.; Rodriguez-Ortega, W.M.; Nicolas, N.; Garcia-Sanchez, F. The effects of amino acids fertilization incorporated to the nutrient solution on mineral composition and growth in tomato seedlings. Span. J. Agricult. Res. 2011, 9, 852–861. [Google Scholar] [CrossRef] [Green Version]
- Popko, M.; Michalak, I.; Wilk, R.; Gramza, M.; Chojnacka, K.; Górecki, H. Effect of the new growth biostimulants based on amino acids on yield and grain quality of winter wheat. Molecules 2018, 23, 470. [Google Scholar] [CrossRef] [Green Version]
- Mohga, K.; Hosni, A.M.; Ibrahim, A.K.; Hewidy, M. Effect of amino Acids application on the growth and quality of octopus tree (Schefflera actinophylla L.). J. Biol. Chem. Environ. Sci. 2016, 11, 283–293. [Google Scholar]
- Sarwar, M.; Frankenberger, W.T., Jr. Influence of L-tryptophan and auxins applied to the rhizosphere on the vegetative growth of Zea mays L. Plant Soil 1994, 160, 97–104. [Google Scholar] [CrossRef]
- Kravchenko, L.V.; Azarova, T.S.; Makarova, N.M.; Tikhonovich, I.A. The effect of tryptophan of plant root metabolites on the phytostimulating activity of rhizobacteria. Mikrobiology 2004, 73, 195–198. [Google Scholar] [CrossRef]
- Oleszczuk, N.; Castro, J.T.; da Silva, M.M.; Korn, M.; Welz, B.; Vale, M.G. Method development for the determination of manganese, cobalt and copper in green coffee comparing direct solid sampling electrothermal atomic absorption spectrometry and inductively coupled plasma optical emission spectrometry. Talanta 2007, 73, 862–869. [Google Scholar] [CrossRef] [PubMed]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of diffrent resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Ostrowska, A.; Gawliński, A.; Szczubiałka, Z. Methods of Analysis and Evaluation of Soils and Plants; Institute of Environmental Protection: Warszawa, Poland, 1991. (In Polish) [Google Scholar]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Szparaga, A.; Kocira, S.; Kocira, A.; Czerwinska, E.; Swieca, M.; Lorencowicz, E.; Kornas, R.; Koszely, M.; Oniszczuk, T. Modification of growth, yield, and the nutraceutical and antioxidative potential of soybean through the use of synthetic biostimulants. Front. Plant Sci. 2018, 9, 1401. [Google Scholar] [CrossRef]
- Ahmad, R.; Khalid, A.; Arshad, M.; Zahir, Z.A.; Mahmood, T. Effect of compost enriched with N and L-tryptophan on soil and maize. Agron. Sustain. Dev. 2008, 28, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Souri, M.K.; Yaghoubi, F.; Moghadamyar, M. Growth and quality of cucumber, tomato, and green bean plants under foliar and soil applications of an aminochelate fertilizer. Horticult. Environ. Biotechnol. 2017, 58, 530–536. [Google Scholar] [CrossRef]
- Mohammandipour, N.; Souri, M.K. Effects of different levels of glycine in the nutrient solution on the growth, nutrirnt composition, and antioxidant activity of coriander (Coriandrum sativum L.). Acta Agrobot. 2019, 72, 1759. [Google Scholar] [CrossRef]
- Hammad, S.A.R.; Ali, O.A.M. Physiological and biochemical studies on drought tolerance of wheat plants by application of amino acids and yeast extract. Ann. Agricult. Sci. 2014, 59, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Filipek-Mazur, B.; Gorczyca, O.; Tabak, M. Effect of mineral containing sulfur on the content of nitrogen, sulfur and sulfur amono acids in spring rape seeds and winter wheat grains. Fragm. Agron. 2017, 34, 33–43. [Google Scholar]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxic. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Morkunas, I.; Woźniak, A.; Van Chung, M.; Rucińska-Sobkowiak, R.; Jeandet, P. The role of heavy metals in plant response to biotic stress. Molecules 2018, 23, 2320. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Determination | Unit | Value Determination (Mean ± S.D. 1) |
|---|---|---|
| pH H2O | - | 4.87 ± 0.04 |
| pH KCl | - | 3.79 ± 0.06 |
| Electrical conductivity (EC) | µS cm−1 | 68.7 ± 9.8 |
| Total carbon | g kg−1 DM 2 | 18.1 ± 2.0 |
| Total nitrogen | g kg−1 DM | 2.08 ± 0.07 |
| Total sulfur | g kg−1 DM | 0.15 ± 0.01 |
| Total magnesium | g kg−1 DM | 3.13 ± 0.02 |
| Total potassium | g kg−1 DM | 3.08 ± 0.25 |
| Total phosphorus | g kg−1 DM | 1.01 ± 0.01 |
| Total calcium | g kg−1 DM | 1.70 ± 0.01 |
| Total sodium | g kg−1 DM | 0.74 ± 0.04 |
| Sand | g kg−1 DM | 230 |
| Silt | g kg−1 DM | 630 |
| Clay | g kg−1 DM | 140 |
| Treatments | Aboveground Parts | Roots | ||||
|---|---|---|---|---|---|---|
| Ash | N | S | Ash | N | S | |
| g kg−1 DM | g kg−1 DM | |||||
| Control | 201.6 b | 49.3 a | 9.28 b | 51.1 a | 46.4 b | 11.54 b |
| NPKS | 163.9 a | 47.3 a | 6.11 b | 74.6 b | 34.3 a | 9.26 a |
| NPKS + L-TRP | 137.5 a | 46.7 a | 6.03 b | 73.5 b | 25.6 a | 13.35 b |
| Treatments | Nitrogen | Sulfur | ||||
|---|---|---|---|---|---|---|
| Aboveground Parts | Roots | Σ | Aboveground Parts | Roots | Σ | |
| g pot−1 | ||||||
| Control | 0.038 a | 0.011 b | 0.049 a | 0.007 a | 0.003 a | 0.010 a |
| NPKS | 0.077 b | 0.007 a | 0.084 b | 0.010 b | 0.002 a | 0.012 a |
| NPKS + L-TRP | 0.111 c | 0.011 b | 0.122 c | 0.014 c | 0.006 b | 0.020 b |
| Treatments | N:S | N:P | K:Ca | K:Mg | K:(Ca + Mg) | (K + Na): (Ca + Mg) |
|---|---|---|---|---|---|---|
| Mass Rations | Ionic Rations | |||||
| Control | 5.3 a | 3.8 a | 14.6 b | 34.4 b | 10.3 b | 10.7 b |
| NPKS | 7.8 b | 5.5 ab | 14.3 b | 34.7 b | 10.1 b | 10.0 b |
| NPKS + L-TRP | 7.8 b | 7.7 c | 11.2 a | 27.2 a | 7.9 a | 7.7 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gondek, K.; Mierzwa-Hersztek, M. Effect of Soil-Applied L-tryptophan on the Amount of Biomass and Nitrogen and Sulfur Utilization by Maize. Agronomy 2021, 11, 2582. https://doi.org/10.3390/agronomy11122582
Gondek K, Mierzwa-Hersztek M. Effect of Soil-Applied L-tryptophan on the Amount of Biomass and Nitrogen and Sulfur Utilization by Maize. Agronomy. 2021; 11(12):2582. https://doi.org/10.3390/agronomy11122582
Chicago/Turabian StyleGondek, Krzysztof, and Monika Mierzwa-Hersztek. 2021. "Effect of Soil-Applied L-tryptophan on the Amount of Biomass and Nitrogen and Sulfur Utilization by Maize" Agronomy 11, no. 12: 2582. https://doi.org/10.3390/agronomy11122582
APA StyleGondek, K., & Mierzwa-Hersztek, M. (2021). Effect of Soil-Applied L-tryptophan on the Amount of Biomass and Nitrogen and Sulfur Utilization by Maize. Agronomy, 11(12), 2582. https://doi.org/10.3390/agronomy11122582