
Relationship between Delayed Leaf Senescence (Stay-Green) and Agronomic and Physiological Characters in Maize (Zea mays L.)

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
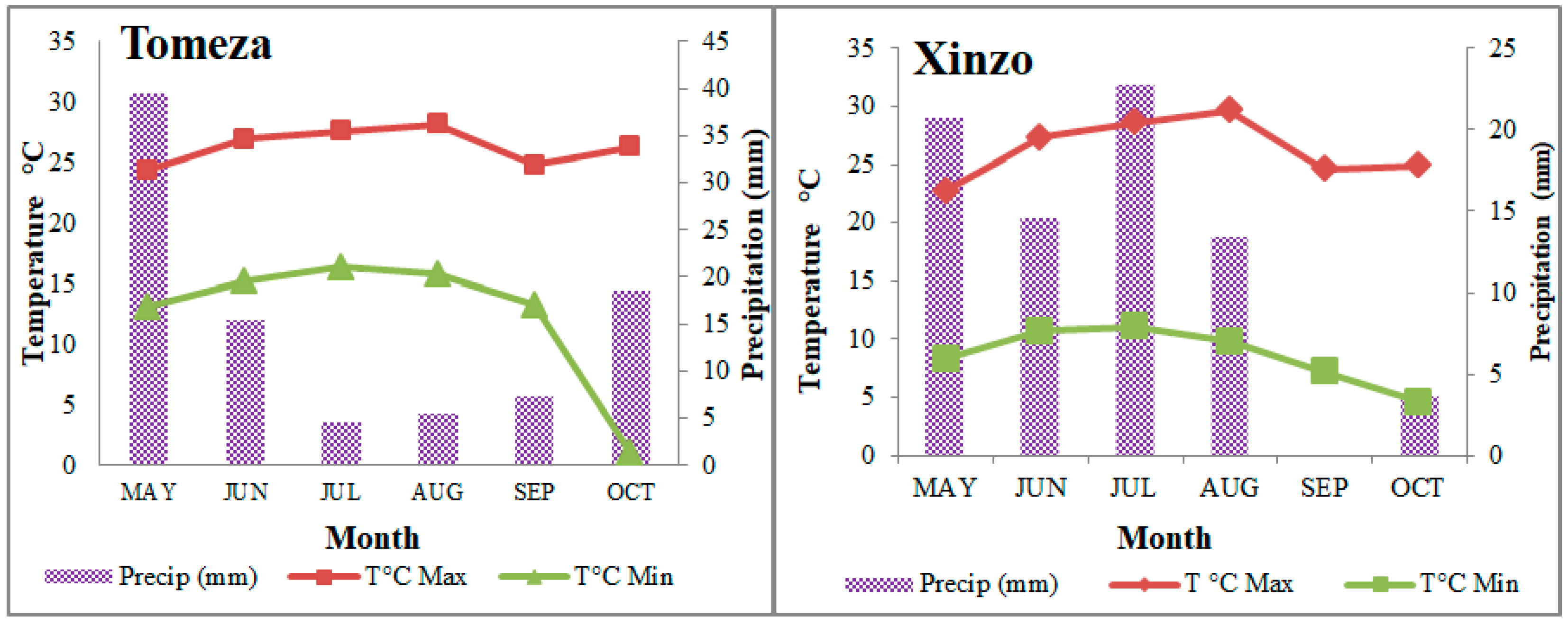
2.1. Locations and Plant Materials
2.2. Experimental Design
2.3. Field Measurements
2.4. Statistical Analyses
3. Results
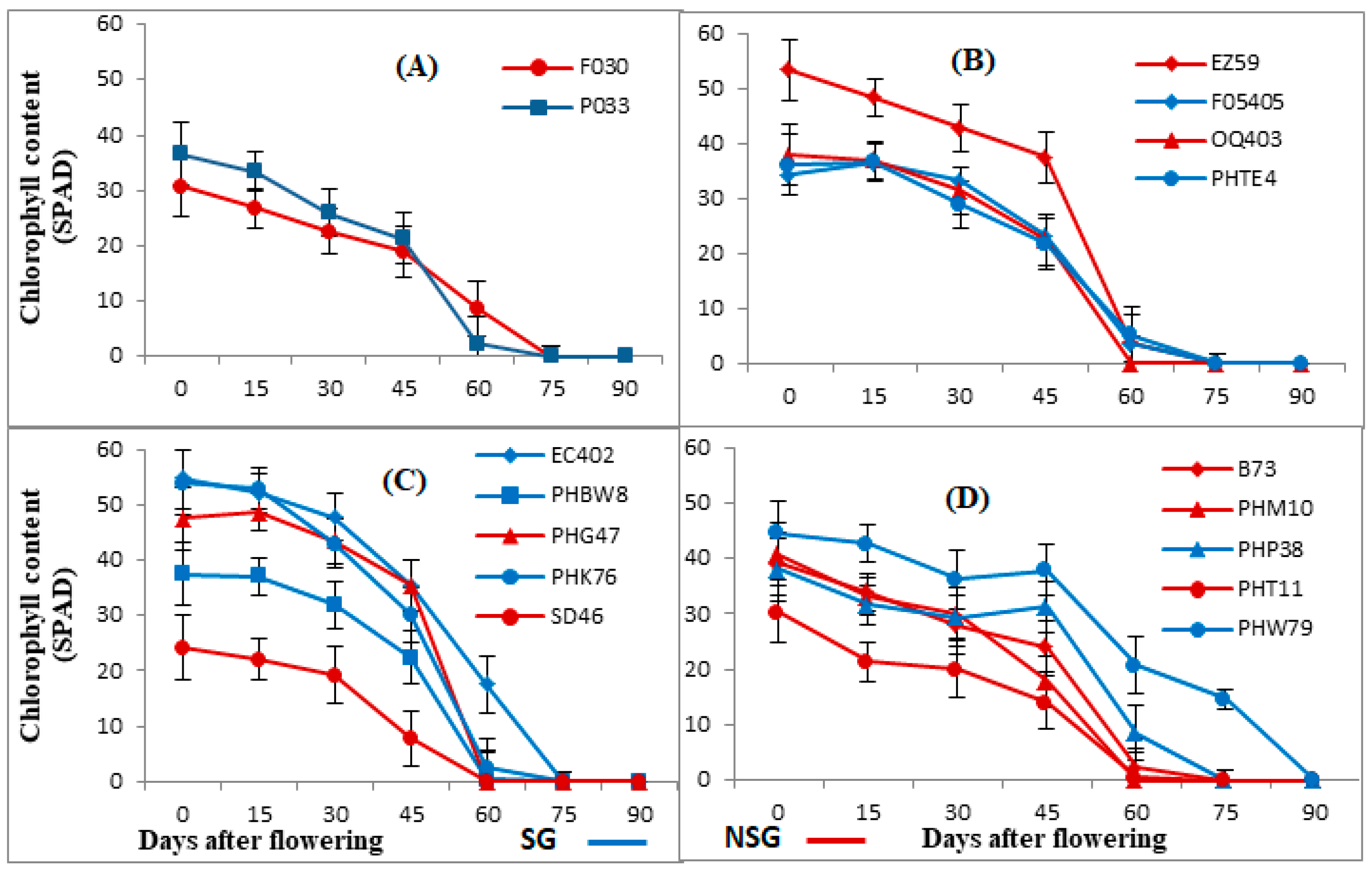
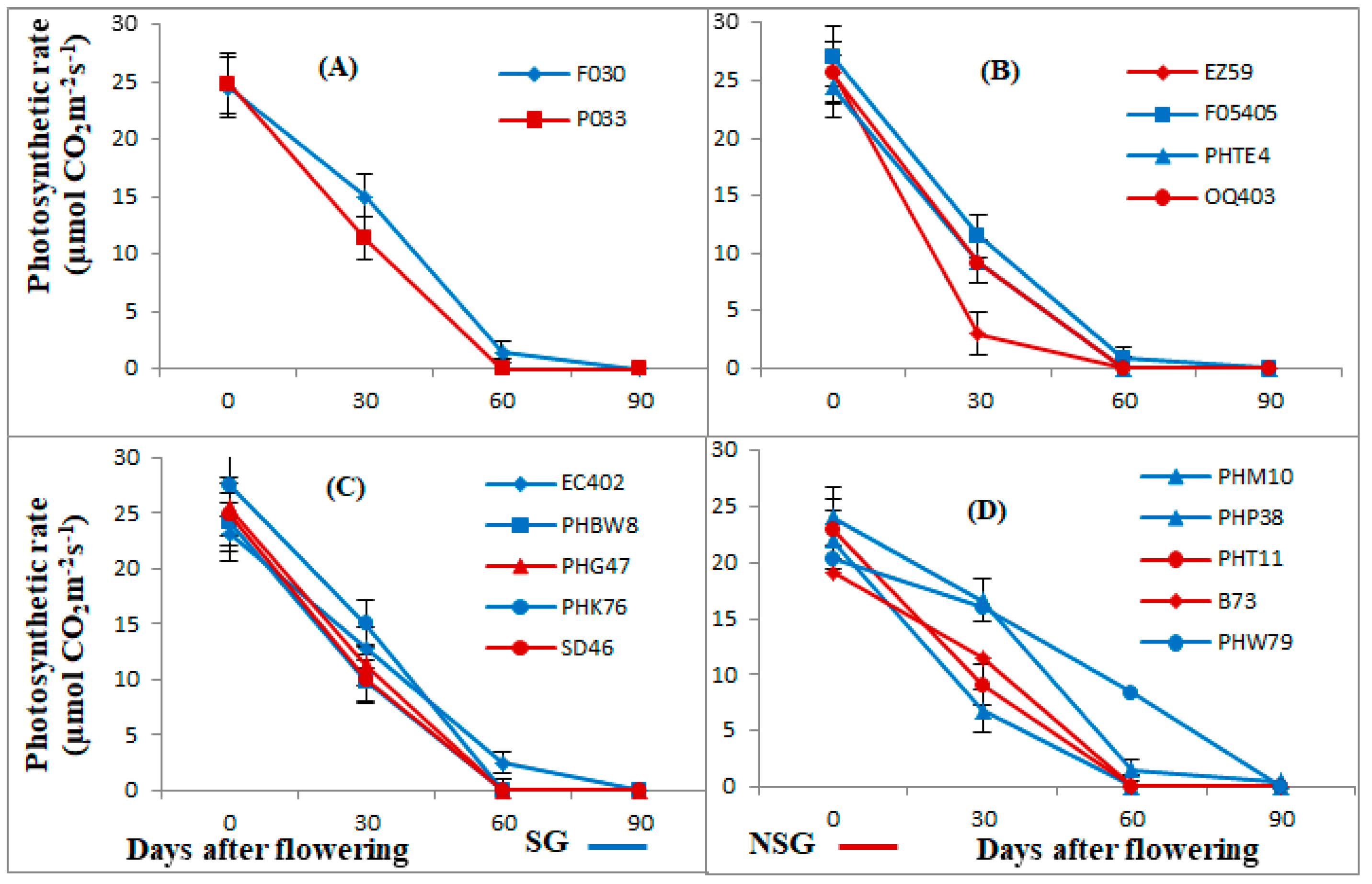
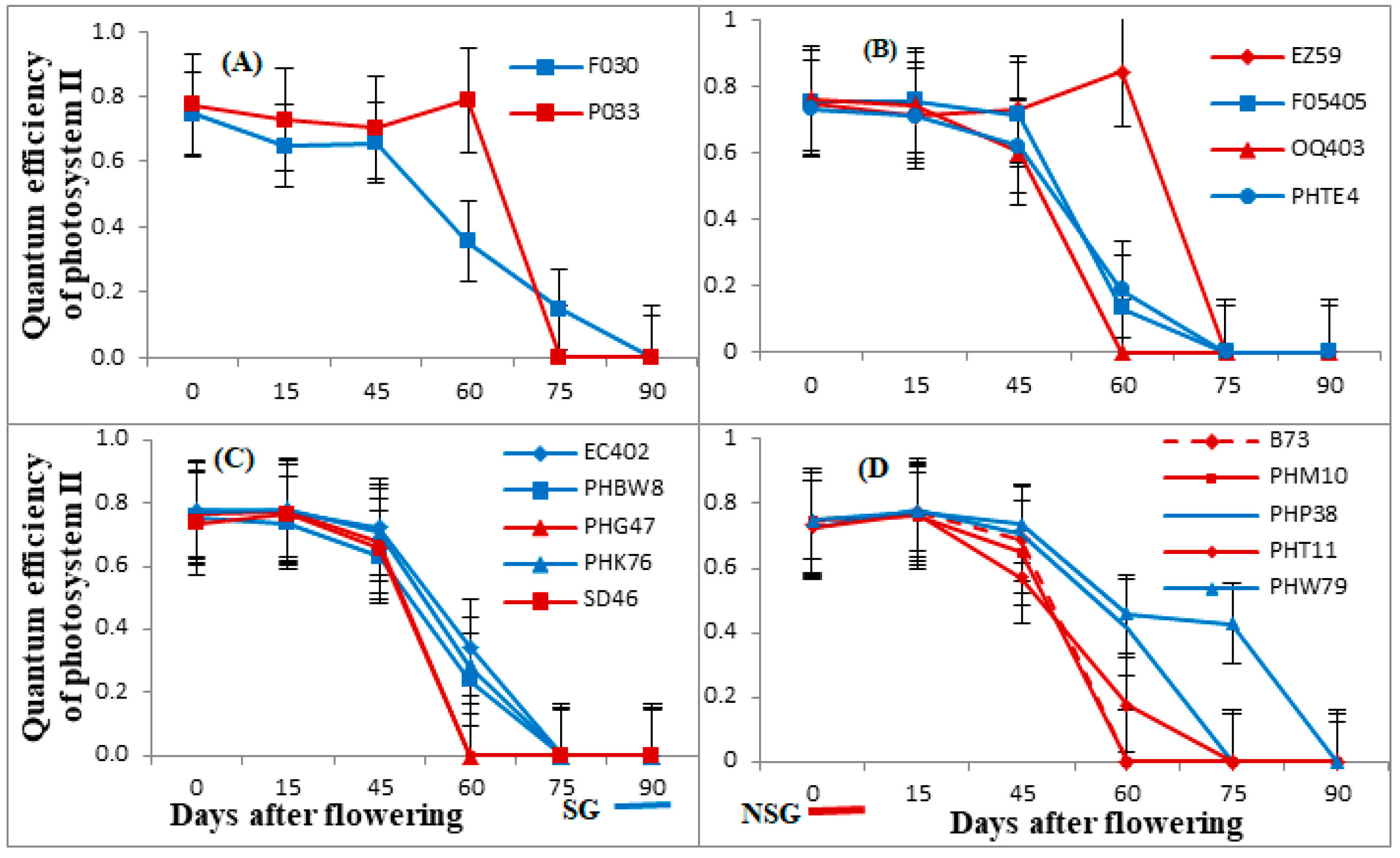
3.1. Photosynthetic Rate, Chlorophyll Content, and Quantum Efficiency of PSII (Fv/Fm)
3.2. Stover and Kernel N Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swanckaert, J.; Pannecoucque, J.; Van Waes, J.; Steppe, K.; Van Labeke, M.C.; Reheul, D. Stay-green characterization in Belgian forage maize. J. Agric. Sci. 2017, 155, 766–776. [Google Scholar] [CrossRef]
- Trachsel, S.; Sun, D.; SanVicente, F.M.; Zheng, H.; Atlin, G.N.; Suarez, E.A.; Babu, R.; Zhang, X. Identification of QTL for early vigor and stay-green conferring tolerance to drought in two connected advanced backcross populations in tropical maize (Zea mays L.). PLoS ONE 2016, 11, 1–22. [Google Scholar]
- Nooden, L.D.; Guiamet, J.J.; Jobn, I. Senescence mechanisms. Physiologia Plantarum 1997, 101, 746–753. [Google Scholar] [CrossRef]
- Lim, P.O.; Kim, H.J.; Nam, H.G. Leaf Senescence. Annu. Rev. Plant Biol. 2007, 58, 115–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.; Wang, X.; Zhang, J.; Liu, P.; Zhao, B.; Li, G.; Dong, S. The role of nitrogen in leaf senescence of summer maize and analysis of underlying mechanisms using comparative proteomics. Plant Science 2015, 233, 72–81. [Google Scholar] [CrossRef]
- Valentinuz, O.; Tollenaar, M. Vertical profile of leaf senescence during the grain-filling period in older and newer maize hybrids. Crop Sci. 2004, 44, 827–834. [Google Scholar]
- Yang, J.; Zhang, J. Grain filling of cereals under soil drying. New Phytol. 2006, 169, 223–236. [Google Scholar] [CrossRef]
- Tollenaar, M.; Ahmadzadeh, A.; Lee, E.A. Physiological basis of heterosis for grain yield in maize. Crop Sci. 2004, 44, 2086–2094. [Google Scholar] [CrossRef]
- Lee, E.A.; Tollenaar, M. Physiological basis of successful breeding strategies for maize grain yield. Crop Sci. 2007, 47, 1–14. [Google Scholar] [CrossRef]
- Ciampitti, I.; Vyn, T. A comprehensive study of plant density consequences on nitrogen uptake dynamics of maize plants from vegetative to reproductive stages. Field Crop. Res. 2011, 121, 2–18. [Google Scholar] [CrossRef]
- Gallais, A.; Coque, M.; Le Gouis, J.; Prioul, J.L.; Hirel, B.; Quilléré, I. Estimating the proportion of nitrogen remobilization and of postsilking nitrogen uptake allocated to maize kernels by nitrogen-15 labeling. Crop Sci. 2007, 47, 685–693. [Google Scholar]
- Ning, P.; Li, S.; Yu, P.; Zhang, Y.; Li, C. Post-silking accumulation and partitioning of dry matter, nitrogen, phosphorus and potassium in maize varieties differing in leaf longevity. Field Crop. Res. 2013, 144, 19–27. [Google Scholar] [CrossRef]
- Thomas, H.; Howarth, C.J. Five ways to stay green. J. Exp. Bot. 2000, 51, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kante, M.; Revilla, P.; De La Fuente, M.; Caicedo, M.; Ordás, B. Stay-green QTLs in temperate elite maize. Euphytica 2016, 207, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Thomas, H.; Ougham, H. The stay-green trait. J. Exp. Bot. 2014, 65, 3889–3900. [Google Scholar]
- Mueller, S.M.; Vyn, T.J. Maize plant resilience to N stress and post-silking N capacity changes over time: A review. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bender, R.R.; Haegele, J.W.; Ruffo, M.L.; Below, F.E. Nutrient uptake, partitioning, and remobilization in modern, transgenic insect-protected maize hybrids. Agron. J. 2013, 105, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Pommel, B.; Gallais, A.; Coque, M.; Quillere, I.; Hirel, B.; Prioul, J.L.; Andrieu, B.; Floriot, M. Carbon and nitrogen allocation and grain filling in three maize hybrids differing in leaf senescence. Eur. J. Agron. 2006, 24, 203–211. [Google Scholar] [CrossRef]
- Luche, H.D.S.; da Silva, J.A.G.; Nornberg, R.; Zimmer, C.M.; Arenhardt, E.G.; da Rosa Caetano, V.; da Maia, L.C.; de Oliveira, A.C. Stay-green effects on adaptability and stability in wheat. Afr. J. Agric. Res. 2015, 10, 1142–1149. [Google Scholar]
- Reguera, M.; Peleg, Z.; Abdel Tawab, Y.M.; Tumimbang, E.B.; Delatorre, C.A.; Blumwald, E. Stress-induced cytokinin synthesis increases drought tolerance through the coordinated regulation of carbon and nitrogen assimilation in rice. Am. Soc. Plant Biol. 2013, 163, 1609–1622. [Google Scholar] [CrossRef] [Green Version]
- Munaiz, E.D.; Martínez, S.; Kumar, A.; Caicedo, M.; Ordás, B. The senescence (stay-green)—An important trait to exploit crop residuals for bioenergy. Energies 2020, 13, 790. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Fengler, K.A.; Van Hemert, J.L.; Gupta, R.; Mongar, N.; Sun, J.; Allen, W.B.; Wang, Y.; Weers, B.; Mo, H.; et al. Identification and characterization of a novel stay-green QTL that increases yield in maize. Plant Biotechnol. J. 2019, 17, 2272–2285. [Google Scholar] [CrossRef]
- Borrell, A.; Hammer, G.; Van Oosterom, E. Stay-green: A consequence of the balance between supply and demand for nitrogen during grain filling? Ann. Appl. Biol. 2001, 138, 91–95. [Google Scholar] [CrossRef]
- Kosgey, J.R.; Moot, D.J.; Fletcher, A.L.; McKenzie, B.A. Dry matter accumulation and post-silking N economy of ‘stay-green’ maize (Zea mays L.) hybrids. Eur. J. Agron. 2013, 51, 43–52. [Google Scholar] [CrossRef]
- Gregersen, P.L.; Holm, P.B.; Krupinska, K. Leaf senescence and nutrient remobilisation in barley and wheat. Plant Biol. 2008, 10, 37–49. [Google Scholar] [CrossRef]
- Chen, K.; Kumudini, S.V.; Tollenaar, M.; Vyn, T.J. Plant biomass and nitrogen partitioning changes between silking and maturity in newer versus older maize hybrids. Field Crops Res. 2015, 183, 315–328. [Google Scholar] [CrossRef]
- White, M.R.; Mikel, M.A.; de Leon, N.; Kaeppler, S.M. Diversity and heterotic patterns in North American proprietary dent maize germplasm. Crop Sci. 2020, 60, 100–114. [Google Scholar] [CrossRef]
- Mikel, M.A.; Dudley, J.W. Evolution of North American dent corn from public to proprietary germplasm. Crop Sci. 2006, 46, 1193–1205. [Google Scholar] [CrossRef]
- Lin, M.; Pang, C.; Fan, S.; Song, M.; Wei, H.; Yu, S. Global analysis of the Gossypium hirsutum L. Transcriptome during leaf senescence by RNA-Seq. BMC Plant Biol. 2015, 15, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.-Y.; Hu, W.-J.; Luo, H.; Xia, Y.; Zhao, Y.; Wang, L.-D.; Zhang, L.-M.; Luo, J.-C.; Jing, H.-C. Transcriptome profiling of developmental leaf senescence in sorghum (Sorghum bicolor). Plant Mol. Biol. 2016, 92, 555–580. [Google Scholar] [CrossRef]
- Yang, Z.; Li, X.; Zhang, N.; Wang, X.; Zhang, Y.; Ding, Y.; Kuai, B.; Huang, X. Mapping and validation of the quantitative trait loci for leaf stay-green-associated parameters in maize. Plant Breed. 2017, 136, 188–196. [Google Scholar] [CrossRef]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Dai, J.; Gao, H.; Dai, Y.; Zou, Q. Changes in activity of energy dissipating mechanisms in wheat flag leaves during senescence. Plant Biol. 2004, 6, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, G.; Gao, H.; Zhang, L.; Yang, C.; Liu, P.; Meng, Q. Characterization of photosynthetic performance during senescence in stay-green and quick-leaf-senescence Zea mays L. inbred lines. PLoS ONE 2012, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Luo, P.; Ren, Z.; Wu, X.; Zhang, H.; Zhang, H.; Feng, J. Structural and biochemical mechanism responsible for the stay-green phenotype in common wheat. Chin. Sci. Bull. 2006, 51, 2595–2603. [Google Scholar] [CrossRef]
- He, P.; Zhou, W.; Jin, J. Effect of nitrogen application on redistribution and transformation of photosynthesized14C during grain formation in two maize cultivars with different senescence appearance. J. Plant Nutr. 2002, 25, 2443–2456. [Google Scholar] [CrossRef]
- Ding, L.; Wang, K.J.; Jiang, G.M.; Biswas, D.K.; Xu, H.; Li, L.F.; Li, Y.H. Effects of nitrogen deficiency on photosynthetic traits of maize hybrids released in different years. Ann. Bot. 2005, 96, 925–930. [Google Scholar] [CrossRef] [Green Version]
- Caicedo, B.M. Mejora Getética de Maiz Para Senescencia Retrasada ‘Stay Green’. Ph.D. Thesis, University Santiago de Compostela, Santiago de Compostela/Lugo, Spain, 2018. [Google Scholar]
- Antonietta, M.; Fanello, D.D.; Acciaresi, H.A.; Guiamet, J.J. Senescence and yield responses to plant density in stay green and earlier-senescing maize hybrids from Argentina. Field Crops Res. 2014, 155, 111–119. [Google Scholar] [CrossRef]
- Kumari, M.; Singh, V.P.; Tripathi, R.; Joshi, A.K. Variation for staygreen trait and its association with canopy temperature depression and yield traits under terminal heat stress in wheat. In Wheat Production in Stressed Environments; Springer: Dordrecht, The Netherlands, 2007; pp. 357–363. [Google Scholar]
- Gous, P.W.; Hickey, L.; Christopher, J.T.; Franckowiak, J.; Fox, G.P. Discovery of QTL for stay-green and heat-stress in barley (Hordeum vulgare) grown under simulated abiotic stress conditions. Euphytica 2016, 207, 305–317. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, L.; Hou, Y.; Wang, P.; Yang, H.; Wei, C.-L. Differential transcriptome analysis of leaves of tea plant (Camellia sinensis) provides comprehensive insights into the defense responses to Ectropis oblique attack using RNA-seq. Funct. Integr. Genom. 2016, 16, 383–398. [Google Scholar] [CrossRef]
- Silva, A.S.; de Carvalho, F.I.F.; Nedel, J.L.; Cruz, P.J.; Peske, S.T.; Simioni, D.; Cargnin, A. Enchimento de sementes em linhas quase-isogênicas de trigo com presença e ausência do caráter ‘stay-green’. Pesquisa Agropecuaria Brasileira 2003, 38, 613–618. [Google Scholar] [CrossRef]
- Uhart, S.A.; Andrade, F.H. Nitrogen and carbon accumulation and remobilization during grain filling in maize under different source/sink ratios. Crop Sci. 1995, 35, 183–190. [Google Scholar] [CrossRef]
- Van Oosterom, E.J.; Chapman, S.C.; Borrell, A.K.; Broad, I.J.; Hammer, G.L. Functional dynamics of the nitrogen balance of sorghum. II. grain filling period. Field Crops Res. 2010, 115, 29–38. [Google Scholar] [CrossRef]
- Rajcan, I.; Tollenaar, M. Source: Sink ratio and leaf senescence in maize: II. Nitrogen metabolism during grain filling. Field Crops Res. 1999, 60, 255–265. [Google Scholar] [CrossRef]
- Subedi, K.D.; Ma, B.L. Nitrogen uptake and partitioning in stay-green and leafy maize hybrids. Crop Sci. 2005, 45, 740–747. [Google Scholar] [CrossRef]
- Fu, W.; Wang, Y.; Ye, Y.; Zhen, S.; Zhou, B.; Wang, Y.; Hu, Y.; Zhao, Y.; Huang, Y. Grain yields and nitrogen use efficiencies in different types of stay-green maize in response to nitrogen fertilizer. Plants 2020, 9, 474. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Reisdorf-Cren, M.; Orsel, M. Leaf nitrogen remobilisation for plant development and grain filling. Plant Biol. 2008, 10, 23–36. [Google Scholar] [CrossRef]
- Thomas, H.; Smart, C.M. Crops that stay green1. Ann. Appl. Biol. 1993, 123, 193–219. [Google Scholar] [CrossRef]
- Bekavac, G.; Purar, B.; Stojakovic, M.; Jockovic, D.J.; Ivanovic, M.; Nastasic, A. Genetic analysis of stay-green trait in broad-based maize populations. Cereal Res. Commun. 2007, 35, 31–41. [Google Scholar] [CrossRef]
- Melchinger, A.E.; Dhillon, B.S.; Grieder, C.; Orsini, E.; Mittweg, G.; Montes, J.M. Determination of methane fermentation yield and its kinetics by near infrared spectroscopy and chemical composition in maize. J. Near Infrared Spectrosc. 2011, 19, 463–477. [Google Scholar]
- Singh, V.; Haken, A.E.; Paulsen, M.R.; Eckhoff, S.R. Starch yield sensitivity of maize hybrids to drying temperature and harvest moisture content. Starch/Staerke 1998, 50, 181–183. [Google Scholar] [CrossRef]
- Maiorano, A.; Fanchini, D.; Donatelli, M. MIMYCS. Moisture, a process-based model of moisture content in developing maize kernels. Eur. J. Agron. 2014, 59, 86–95. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
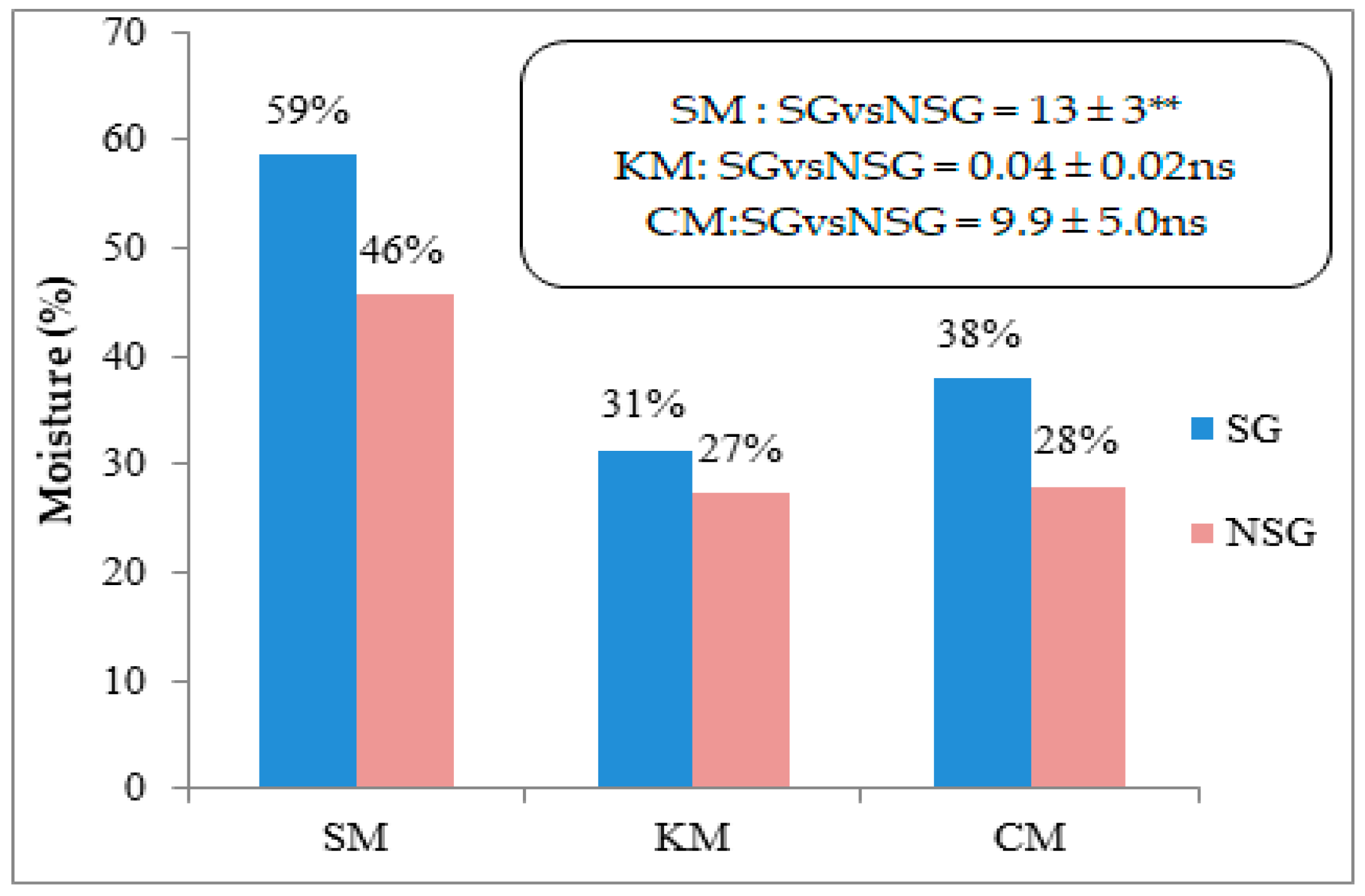{kind=link}
{kind=link}
| Inbred Lines | GS | FG | Heterotic Group | Origen |
|---|---|---|---|---|
| F030 | SG | 1 | European Flint | University of Hohenheim |
| F05405 | SG | 2 | Unknown | INRA |
| PHTE4 | SG | 2 | PHJ40/Other SS * | exPVP PI 9400094 |
| EC402 | SG | 3 | Stiff Stalk Synthetic | Centro de Investigaciones Agrarias de Mabegondo (Xunta de Galicia) |
| PHK76 | SG | 3 | B37/Lancaster/Iodent * | exPVP PI 601496 |
| PHBW8 | SG | 3 | PHJ40/Other SS * | exPVP PI 559936 |
| PHW79 | SG | 4 | Oh7/Midland ** | exPVP PI 601576 |
| PHP38 | SG | 1 | B37/PHG39 * | exPVP PI 543845 |
| P033 | NSG | 2 | Iodent | University of Hohenheim |
| EZ59 | NSG | 2 | European Fllint | Estación Experimental de Aula Dei (CSIC) |
| OQ403 | NSG | 3 | Pioneer 3901 hybrid X K81-336 | exPVP PI 559920 |
| PHG47 | NSG | 3 | Lancaster * | exPVP PI 601318 |
| SD46 | NSG | 4 | Pioneer hybrid 3709 | South Dakota University |
| B73 | NSG | 4 | ||
| PHT11 | NSG | 4 | B37/PHG39 * | exPVP PI 548807 |
| PHM10 | NSG | 4 | B37/PHG39 * | exPVP PI 601778 |
| Total N in Plant | N in Kernels | N in Stover | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Genotypes | SG Trait | TN (gplant−1) | TN_UF (gplant−1) | TN_AF (gplant−1) | KN (gplant−1) | KN_R (%) | KN_NR (%) | SN_R (gplant−1) | SN_NR (gplant−1) |
| F030 | SG | 3.08 ± 0.36 | 2.37(77%) ± 0.37 | 0.70(23%) ± 0.40 | 2.01 ± 0.54 | 64 ± 16 | 36 ± 16 | 1.33 (52%) ± 0.47 | 1.04(48%) ± 0.23 |
| P033 | NSG | 2.90 ± 0.37 | 2.17(76%) ± 0.37 | 0.71(24%) ± 0.40 | 2.12 ± 0.54 | 65 ± 16 | 35 ± 16 | 1.42(60%) ± 0.47 | 0.76(39%) ± 0.23 |
| EZ59 | NSG | 2.25 ± 0.36 | 2.66(100%) ± 0.37 | −0.41 (0%) ± 0.40 | 1.24 ± 0.54 | 100 ± 16 | 0 | 1.65(56%) ± 0.47 | 1.00(44%) ± 0.23 |
| F05405 | SG | 3.63 ± 0.36 | 2.27(63%) ± 0.37 | 1.36(37%) ± 0.40 | 2.11 ± 0.54 | 35 ± 16 | 65 ± 16 | 0.74(30%) ± 0.47 | 1.52(70%) ± 0.23 |
| OQ403 | NSG | 2.35 ± 0.36 | 1.78(79%) ± 0.37 | 0.56(21%) ± 0.40 | 1.40 ± 0.54 | 68 ± 16 | 32 ± 16 | 0.84(47%) ± 0.47 | 0.94(53%) ± 0.23 |
| PHTE4 | SG | 2.76 ± 0.36 | 1.54(57%) ± 0.37 | 1.21(43%) ± 0.40 | 1.78 ± 0.54 | 33 ± 16 | 67 ± 16 | 0.57(37%) ± 0.47 | 0.97(63%) ± 0.23 |
| EC402 | SG | 4.55 ± 0.36 | 3.13(68%) ± 0.37 | 1.40(32%) ± 0.40 | 2.86 ± 0.54 | 48 ± 16 | 52 ± 16 | 1.47(41%) ± 0.47 | 1.67(59%) ± 0.23 |
| PHBW8 | SG | 3.59 ± 0.36 | 2.67(77%) ± 0.37 | 0.90(23%) ± 0.40 | 2.49 ± 0.54 | 62 ± 16 | 38 ± 16 | 1.60(55%) ± 0.47 | 1.07(45%) ± 0.23 |
| PHG47 | NSG | 2.67 ± 0.36 | 2.03(77%) ± 0.37 | 0.62(23%) ± 0.40 | 1.74 ± 0.54 | 63 ± 16 | 37 ± 16 | 1.13(54%) ± 0.47 | 0.90(46%) ± 0.23 |
| PHK76 | SG | 3.50 ± 0.38 | 2.39(72%) ± 0.40 | 1.11(28%) ± 0.46 | 1.80 ± 0.56 | 45 ± 19 | 55 ± 18 | 0.72(24%) ± 0.50 | 1.68(76%) ± 0.24 |
| SD46 | NSG | 3.18 ± 0.38 | 1.97(72%) ± 0.40 | 1.21(28%) ± 0.46 | 2.07 ± 0.56 | 57 ± 19 | 44 ± 18 | 0.90(48%) ± 0.50 | 1.08(52%) ± 0.24 |
| B73 | NSG | 3.73 ± 0.36 | 2.85(78%) ± 0.37 | 0.83(22%) ± 0.46 | 1.86 ± 0.54 | 62 ± 16 | 38 ± 16 | 1.01(36%) ± 0.47 | 1.82(64%) ± 0.23 |
| PHM10 | NSG | 3.02 ± 0.36 | 2.36(79%) ± 0.37 | 0.62(21%) ± 0.40 | 1.75 ± 0.54 | 60 ± 16 | 40 ± 16 | 1.11(45%) ± 0.47 | 1.23(55%) ± 0.23 |
| PHP38 | SG | 4.15 ± 0.36 | 1.87(53%) ± 0.37 | 2.23(47%) ± 0.40 | 2.48 ± 0.54 | 30 ± 16 | 70 ± 16 | 0.24(26%) ± 0.47 | 1.62(74%) ± 0.23 |
| PHT11 | NSG | 2.99 ± 0.36 | 2.32(80%) ± 0.37 | 0.63(20%) ± 0.40 | 1.43 ± 0.54 | 60 ± 16 | 40 ± 16 | 0.79(34%) ± 0.47 | 1.51(66%) ± 0.23 |
| PHW79 | SG | 4.18 ± 0.36 | 2.29(58%) ± 0.37 | 1.84(42%) ± 0.40 | 2.38 ± 0.54 | 25 ± 16 | 75 ± 16 | 0.53(22%) ± 0.47 | 1.75(78%) ± 0.23 |
| SG vs. NSG | 0.79 ± 0.1 ** | 0.05(−16%) ± 0.1 ns | 0.75(17%) ± 0.14 ** | 0.54 ± 0.09 ** | −27 ± 6 ** | 27 ± 6 ** | −0.21(−12%) ± 0.10 * | 0.26(12%) ± 0.04 ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chibane, N.; Caicedo, M.; Martinez, S.; Marcet, P.; Revilla, P.; Ordás, B. Relationship between Delayed Leaf Senescence (Stay-Green) and Agronomic and Physiological Characters in Maize (Zea mays L.). Agronomy 2021, 11, 276. https://doi.org/10.3390/agronomy11020276
Chibane N, Caicedo M, Martinez S, Marcet P, Revilla P, Ordás B. Relationship between Delayed Leaf Senescence (Stay-Green) and Agronomic and Physiological Characters in Maize (Zea mays L.). Agronomy. 2021; 11(2):276. https://doi.org/10.3390/agronomy11020276
Chicago/Turabian StyleChibane, Nadia, Marlon Caicedo, Susana Martinez, Purificación Marcet, Pedro Revilla, and Bernardo Ordás. 2021. "Relationship between Delayed Leaf Senescence (Stay-Green) and Agronomic and Physiological Characters in Maize (Zea mays L.)" Agronomy 11, no. 2: 276. https://doi.org/10.3390/agronomy11020276
APA StyleChibane, N., Caicedo, M., Martinez, S., Marcet, P., Revilla, P., & Ordás, B. (2021). Relationship between Delayed Leaf Senescence (Stay-Green) and Agronomic and Physiological Characters in Maize (Zea mays L.). Agronomy, 11(2), 276. https://doi.org/10.3390/agronomy11020276