
Impacts of Fallow Conditions, Compost and Silicate Fertilizer on Soil Nematode Community in Salt–Affected Paddy Rice Fields in Acid Sulfate and Alluvial Soils in the Mekong Delta, Vietnam

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study location and Experiment Design
2.2. Soil Sampling and Analyses
2.2.1. Soil Sampling
2.2.2. Nematode Processing, Identification and Community Characteristics
2.2.3. Rice Yield and Straw Assessment
2.3. Data Analyses
3. Results
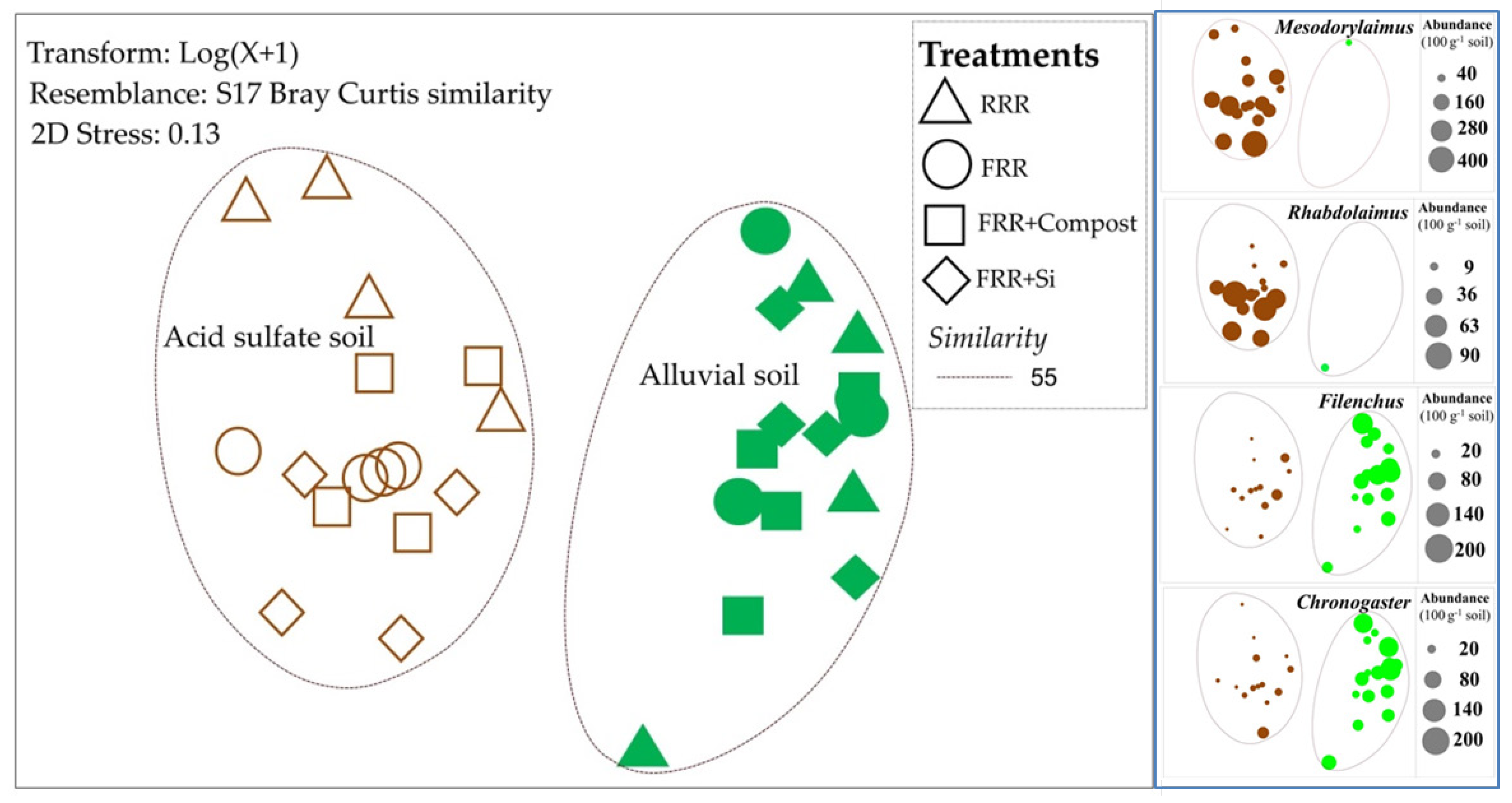
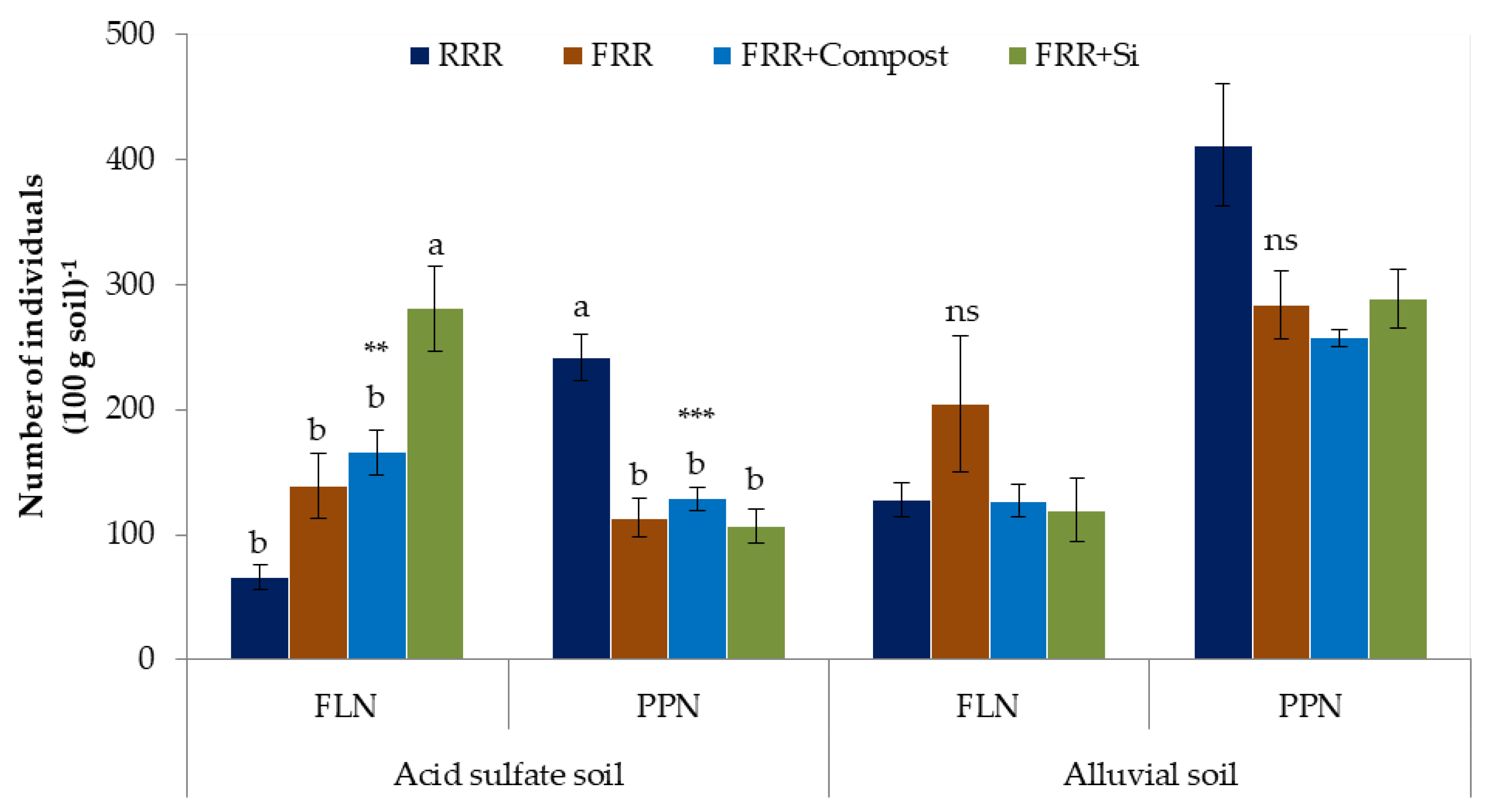
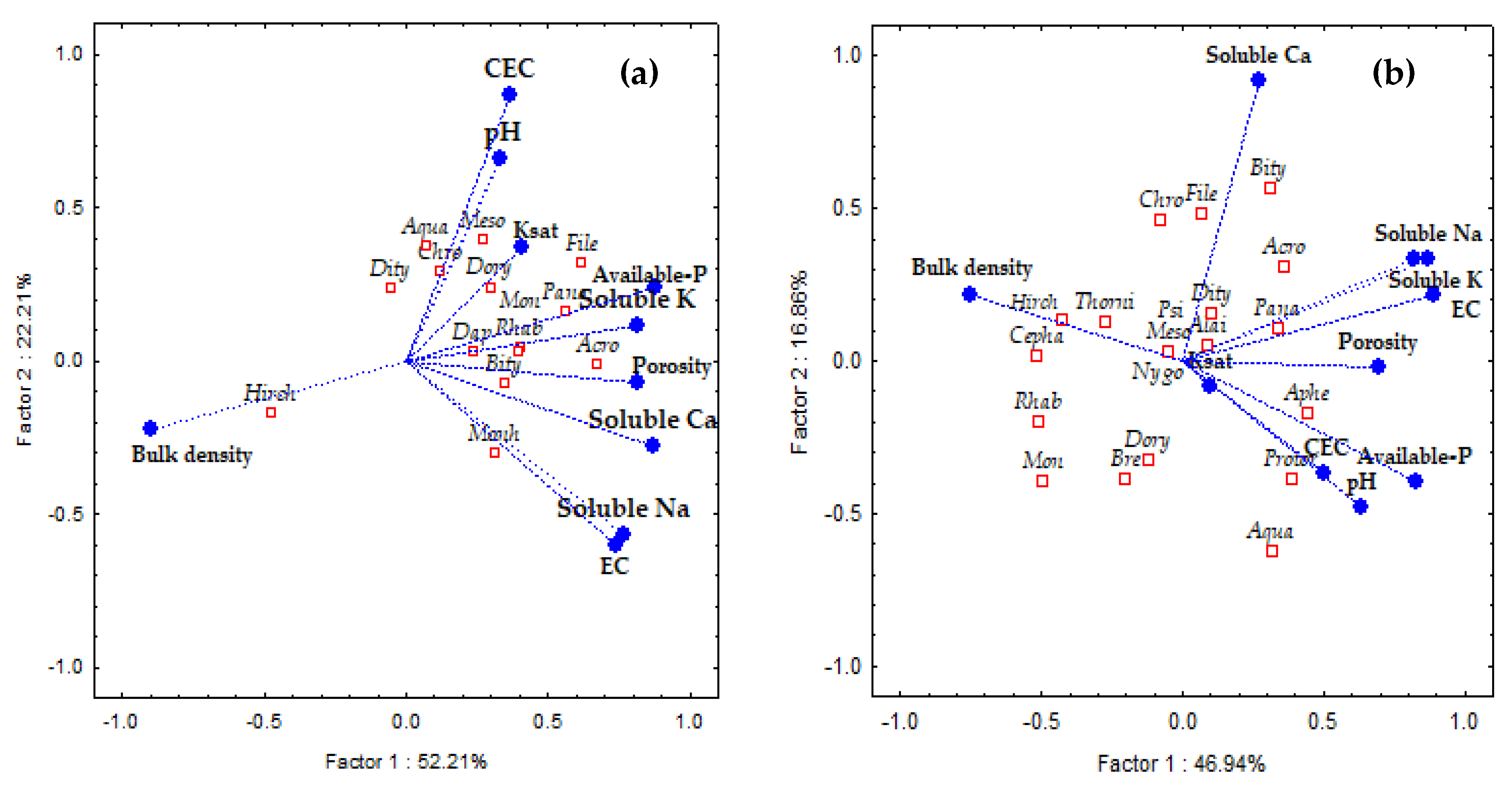
3.1. Impacts of Reduced Rice Cropping, Reduced Rice Cropping Mixed with Compost or Silicate Fertilizer on Nematode Abundance and Composition
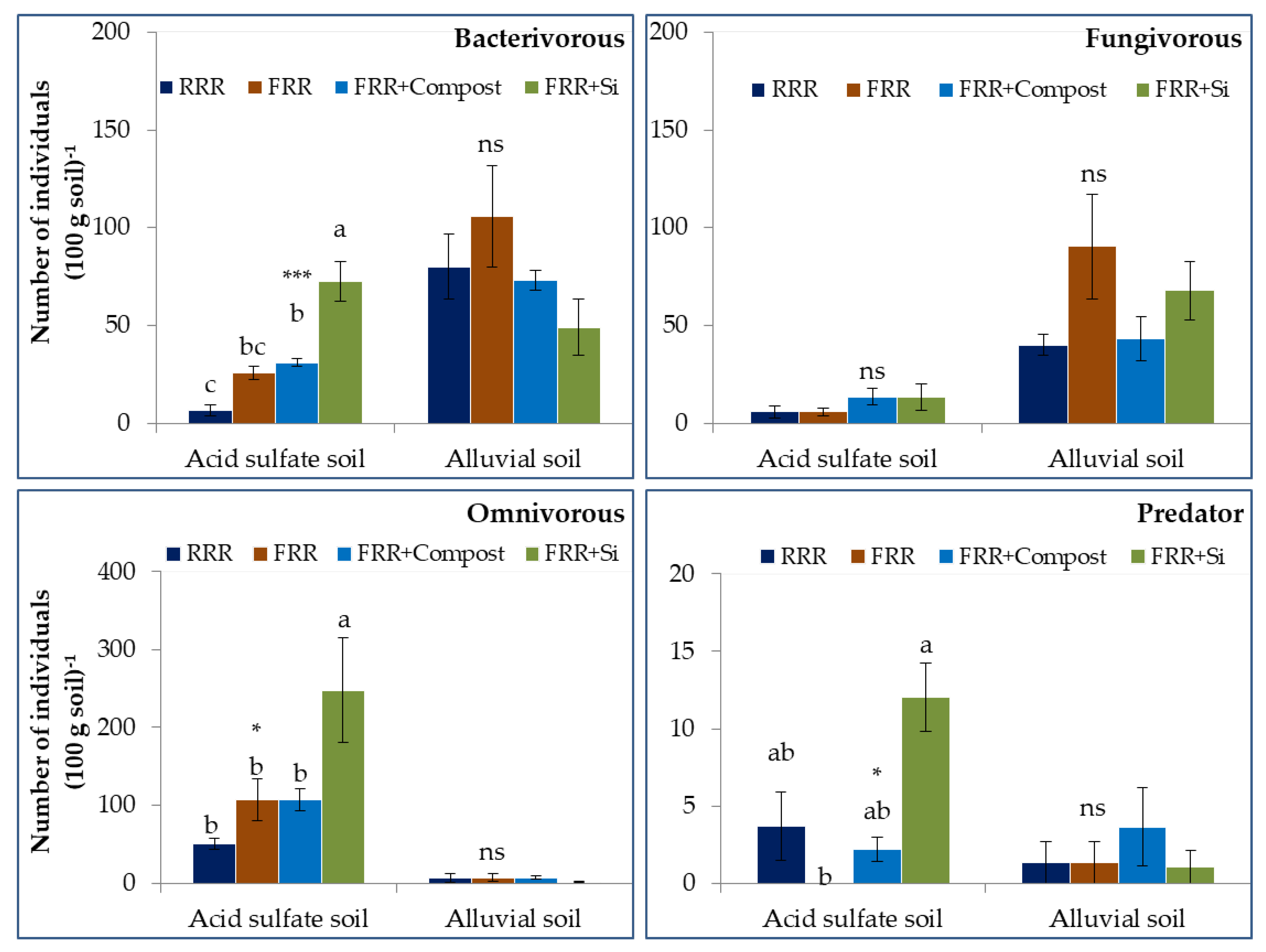
3.2. Impacts of Reduced Rice Cropping, Reduced Rice Cropping Mixed with Compost or Silicate Fertilizer on Trophic Structure of Nematode in Soils
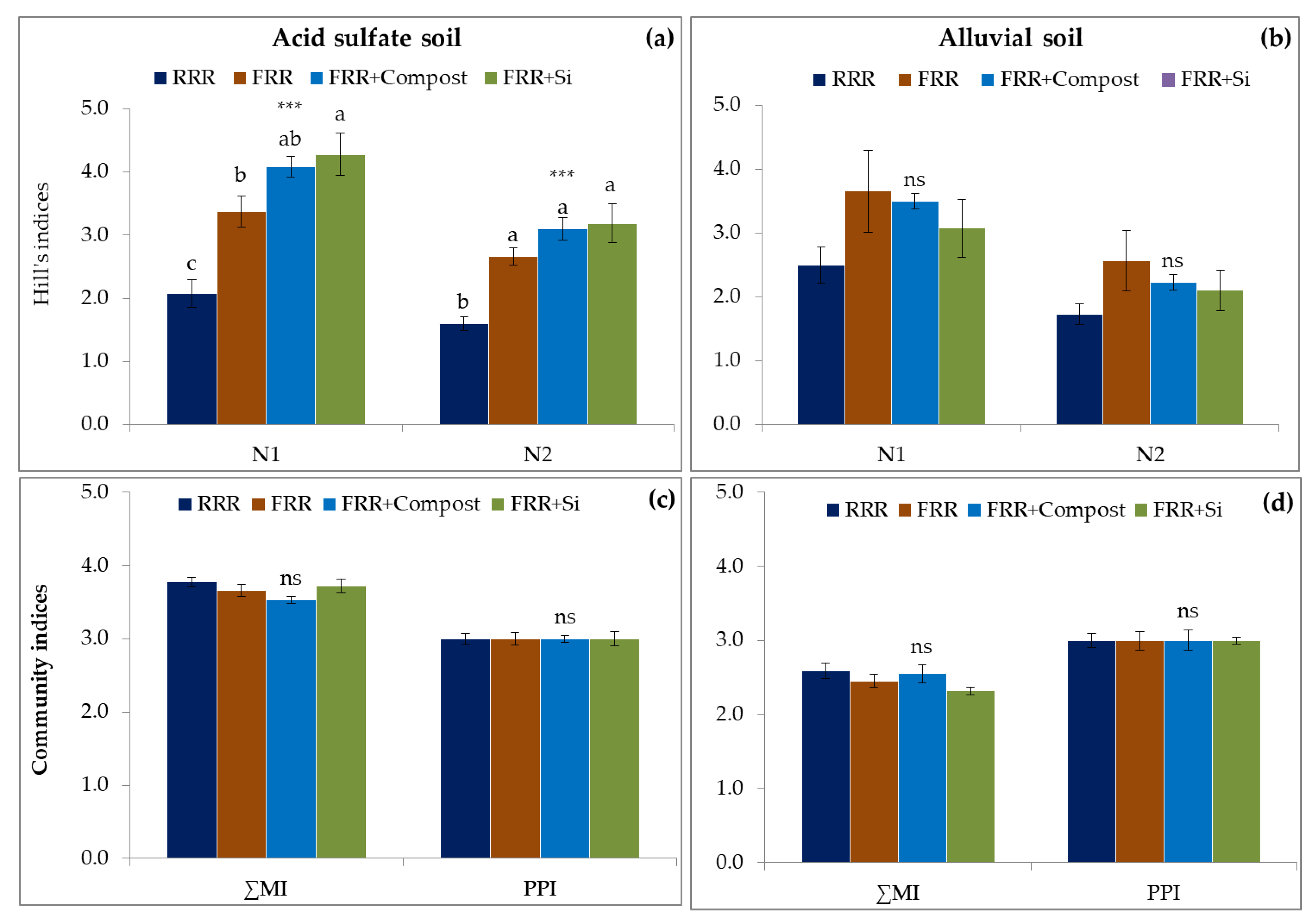
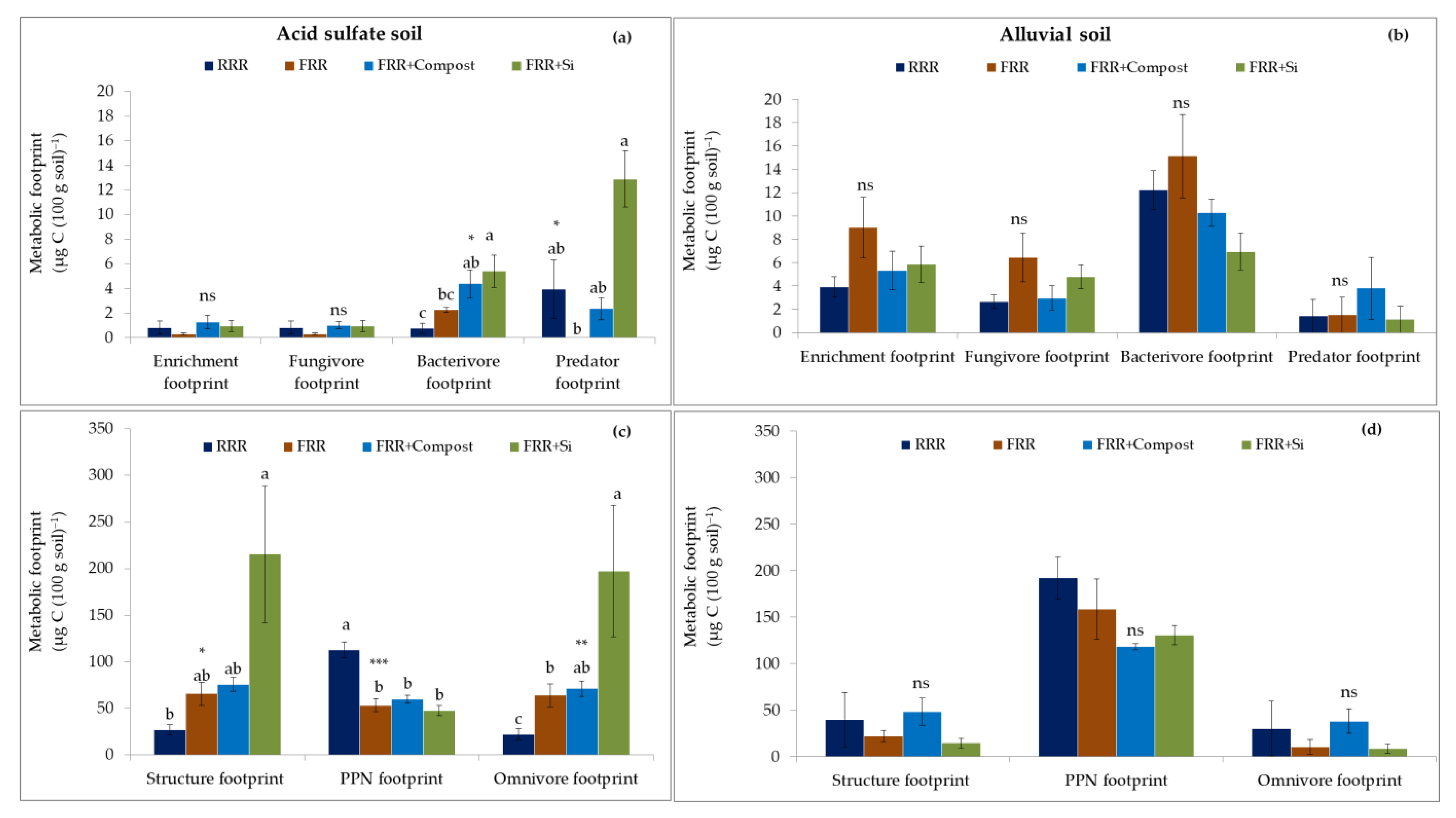
3.3. Impact of Reduced Rice Cropping, Reduced Rice Cropping Mixed with Compost or Silicate Fertilizer on Diversity, Community Indices and Metabolic Footprints of Nematode in Soils
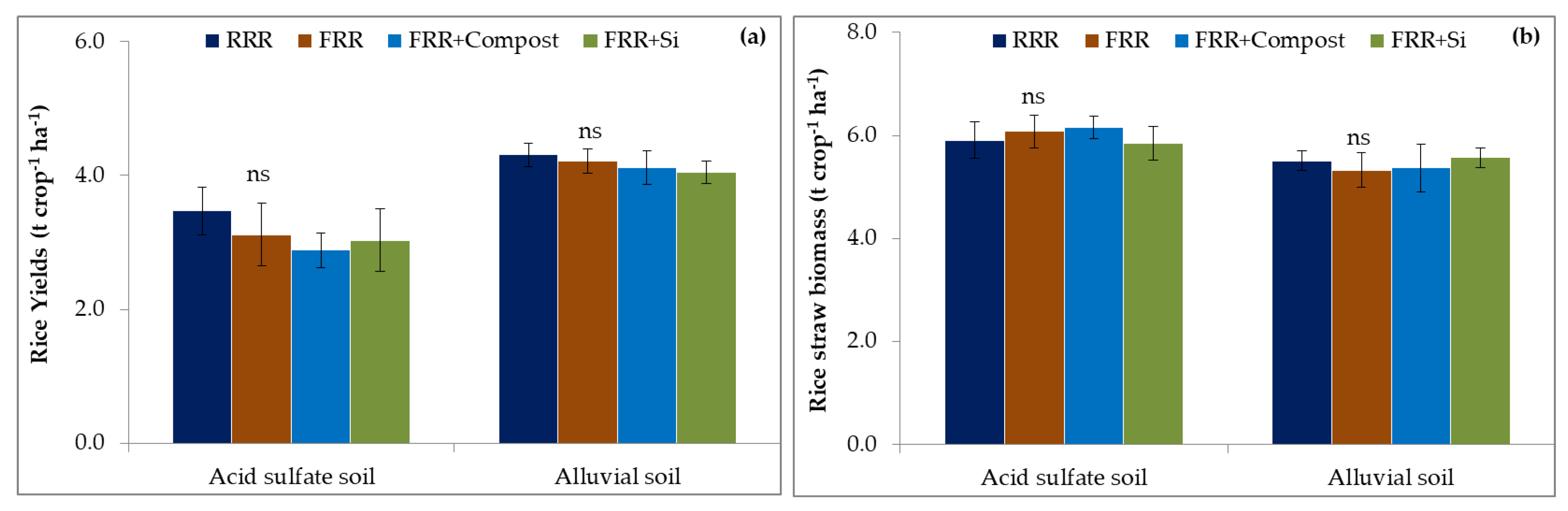
3.4. Impact of Reduced Rice Cropping, Reduced Rice Cropping Mixed with Compost or Silicate Fertilizer on Rice Yield and Straw Biomass and Its Relationship with Abundance of Hirschmanniella in Soils
4. Discussion
4.1. Impacts of Reduced Rice Cropping, Reduced Rice Cropping Mixed with Compost or Silicate Fertilizeron Nematode Abundance and Composition
4.2. Impact of Reduced Rice Cropping Mixed with Compost or Silicate Fertilizer on Trophic Structure of Free-Living Nematode in Soils
4.3. Impact of Reduced Rice Cropping, Reduced Rice Cropping Mixed with Compost or Silicate Fertilizer on Diversity, Community Indices, and Metabolic Footprints of Nematode in Soils
4.4. Effects of Reduced Rice Cropping, Reduced Rice Cropping Mixed with Compost or Silicate Fertilizer on Rice Straw Biomass and Yield
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tran, D.D.; van Halsema, G.; Hellegers, P.; Ludwig, F.; Wyatt, A. Questioning triple rice intensification on the Vietnamese Mekong delta floodplains: An environmental and economic analysis of current land-use trends and alternatives. J. Environ. Manag. 2018, 217, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Stone, E.C.; Hornberger, G.M. Impacts of management alternatives on rice yield and nitrogen losses to the environment: A case study in rural Sri Lanka. Sci. Total Environ. 2016, 542, 271–276. [Google Scholar] [CrossRef]
- Stuart, A.M.; Palenzuela, A.N.; Bernal, C.C.; Ramal, A.F.; Horgan, F.G. Effects of fertiliser applications on survival and recruitment of the apple snail, Pomacea canaliculata (Lamarck). Crop Prot. 2014, 64, 78–87. [Google Scholar] [CrossRef]
- Cassman, K.G.; Harwood, R.R. The nature of agricultural systems-food security and environmental balance. Food Policy 1995, 20, 439–454. [Google Scholar] [CrossRef]
- Nguyen, V.L.; Ta, T.K.O.; Tateishi, M. Late holocene depositional environments and coastal evolution of the Mekong River delta, Southern Vietnam. J. Asian Earth Sci. 2000, 18, 427–439. [Google Scholar] [CrossRef]
- Xuan, V.T.; Quang, N.K.; Tri, L.Q. Rice Cultivation on Acid Sulfate Soils in the Vietnamese Mekong Delta. In Proceedings of the Bangkok Symposium on Acid Sulfate Soils, Bangkok, Thailand, 18–24 January 1982; Dost, H., Breemen, N.V., Eds.; International Institute for Land Reclamation and Improvement, Wageningen: Bangkok, Thailand, 1984; pp. 251–271. [Google Scholar]
- Vu, D.T.; Yamada, T.; Ishidaira, H. Assessing the impact of sea level rise due to climate change on seawater intrusion in Mekong delta, Vietnam. Water Sci. Technol. 2018, 77, 1632–1639. [Google Scholar] [CrossRef] [Green Version]
- Wassmann, R.; Phong, N.D.; Tho, T.Q.; Hoanh, C.T.; Khoi, N.H.; Hien, N.X.; Vo, T.B.T.; Tuong, T.P. High-resolution mapping of flood and salinity risks for rice production in the Vietnamese Mekong delta. Field Crops Res. 2019, 236, 111–120. [Google Scholar] [CrossRef]
- Lee, S.K.; Dang, T.A. Spatio-temporal variations in meteorology drought over the Mekong river delta of Vietnam in the recent decades. Paddy Water Environ. 2019, 17, 35–44. [Google Scholar] [CrossRef]
- Buchan, D.; Gebremikael, M.T.; Ameloot, N.; Sleutel, S.; De Neve, S. The effect of free-living nematodes on nitrogen mineralisation in undisturbed and disturbed soil cores. Soil Biol. Biochem. 2013, 60, 142–155. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T.; de Goede, R.G.M. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Okada, H.; Niwa, S.; Takemoto, S.; Komatsuzaki, M.; Hiroki, M. How different or similar are nematode communities between a paddy and an upland rice fields across a flooding-drainage cycle? Soil Biol. Biochem. 2011, 43, 2142–2151. [Google Scholar] [CrossRef]
- Korthals, G.W.; Alexiev, A.D.; Lexmond, T.M.; Kammenga, J.E.; Bongers, T. Long-term effects of copper and pH on the nematode community in an agroecosystem. Environ. Toxicol. Chem. 1996, 15, 979–985. [Google Scholar] [CrossRef]
- Nguyen, V.S.; Nguyen, T.K.P.; Araki, M.; Perry, R.N.; Tran, B.L.; Chau, M.K.; Min, Y.Y.; Toyota, K. Seasonal variation and vertical distribution of nematode communities and their relation to the soil chemical property and rice productivity of clay soil under triple rice cultivation in the Mekong delta, Vietnam. Nematology 2020, 23, 33–46. [Google Scholar] [CrossRef]
- Ito, T.; Araki, M.; Higashi, T.; Komatsuzaki, M.; Kaneko, N.; Ohta, H. Responses of soil nematode community structure to soil carbon changes due to different tillage and cover crop management practices over a nine-year period in Kanto, Japan. Appl. Soil Ecol. 2015, 89, 50–58. [Google Scholar] [CrossRef]
- Bongiorno, G.; Bodenhausen, N.; Bunemann, E.K.; Brussaard, L.; Geisen, S.; Mader, P.; Quist, C.W.; Walser, J.C.; de Goede, R.G.M. Reduced tillage, but not organic matter input, increased nematode diversity and food web stability in European long-term field experiments. Mol. Ecol. 2019, 28, 4987–5005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, V.S.; Chau, M.K.; Vo, Q.M.; Le, V.K.; Nguyen, T.K.P.; Araki, M.; Perry, R.N.; Tran, A.D.; Dang, D.M.; Tran, B.L.; et al. Impacts of saltwater intrusion on soil nematodes community in alluvial and acid sulfate soils in paddy rice fields in the Vietnamese Mekong delta. Ecol. Indic. 2021, 122, 107284. [Google Scholar] [CrossRef]
- Khoa, L.V. Physical Fertility of Typical Mekong Delta Soils (Vietnam) and Land Suitability Assessment for Alternative Crops with Rice Cultivation. Ph.D. Thesis, Ghent University, Ghent, Belgium, 2002. [Google Scholar]
- Momma, N.; Kobara, Y.; Uematsu, S.; Kita, N.; Shinmura, A. Development of biological soil disinfestations in Japan. Appl. Microbiol. Biotechnol. 2013, 97, 3801–3809. [Google Scholar] [CrossRef]
- Bridge, J.; Plowright, R.A.; Peng, D. Plant Parasitic Nematodes in Subtropical and Tropical Agriculture, 2nd ed.; CAB International Publishing: Wallingford, UK, 2005. [Google Scholar]
- Poussin, J.C.; Neuts, T.; Mateille, T. Interactions between irrigated rice (Oryza sativa) growth, nitrogen amendments and infection by Hirschmanniella oryzae (Nematoda, Tylenchida). Appl. Soil Ecol. 2005, 29, 27–37. [Google Scholar] [CrossRef]
- Nguyen, V.S.; Nguyen, T.K.P.; Araki, M.; Perry, R.N.; Linh, B.T.; Chau, M.K.; Min, Y.Y.; Toyota, K. Effects of cropping systems and soil amendments on nematode community and its relationship with soil physicochemical properties in a paddy rice field in the Vietnamese Mekong delta. Appl. Soil Ecol. 2020, 156, 103683. [Google Scholar] [CrossRef]
- Soriano, I.R.; Reversat, G. Management of Meloidogyne graminicola and yield of upland rice in South-Luzon, Philippines. Nematology 2003, 5, 879–884. [Google Scholar] [CrossRef]
- Hu, C.; Qi, Y.C. Effect of compost and chemical fertilizer on soil nematode community in a Chinese maize field. Eur. J. Soil Biol. 2010, 46, 230–236. [Google Scholar] [CrossRef]
- Huang, W.K.; Ji, H.L.; Gheysen, G.; Debode, J.; Kyndt, T. Biochar-amended potting medium reduces the susceptibility of rice to root-knot nematode infections. BMC Plant Biol. 2015, 15, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.H.; McSorley, R.; Marshall, A.; Gallaher, R.N. Influence of organic Crotalaria juncea hay and ammonium nitrate fertilizers on soil nematode communities. Appl. Soil Ecol. 2006, 31, 186–198. [Google Scholar] [CrossRef]
- Chen, Y.F.; Hu, N.; Zhang, Q.Z.; Lou, Y.L.; Li, Z.F.; Tang, Z.; Kuzyakov, Y.; Wang, Y.D. Impacts of green manure amendment on detritus micro-food web in a double-rice cropping system. Appl. Soil Ecol. 2019, 138, 32–36. [Google Scholar] [CrossRef]
- Bi, L.D.; Zhang, B.; Liu, G.R.; Li, Z.Z.; Liu, Y.R.; Ye, C.; Yu, X.C.; Lai, T.; Zhang, J.G.; Yin, J.M.; et al. Long-term effects of organic amendments on the rice yields for double rice cropping systems in subtropical China. Agric. Ecosyst. Environ. 2009, 129, 534–541. [Google Scholar] [CrossRef]
- Hu, C.; Xia, X.G.; Han, X.M.; Chen, Y.F.; Qiao, Y.; Liu, D.H.; Li, S.L. Soil nematode abundances were increased by an incremental nutrient input in a paddy-upland rotation system. Helminthologia 2018, 55, 322–333. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, M.; Alam, M.M. Utilization of waste materials in nematode control: A review. Bioresour. Technol. 1993, 45, 1–7. [Google Scholar] [CrossRef]
- Atandi, J.G.; Haukeland, S.; Kariuki, G.M.; Coyne, D.L.; Karanja, E.N.; Musyoka, M.W.; Fiaboe, K.K.M.; Bautze, D.; Adamtey, N. Organic farming provides improved management of plant parasitic nematodes in maize and bean cropping systems. Agric. Ecosyst. Environ. 2017, 247, 265–272. [Google Scholar] [CrossRef]
- Khan, Z.; Kim, Y.H. A review on the role of predatory soil nematodes in the biological control of plant parasitic nematodes. Appl. Soil Ecol. 2007, 35, 370–379. [Google Scholar] [CrossRef]
- Khan, A.; Kamran, M.; Imran, M.; Al-Harrasi, A.; Al-Rawahi, A.; Al-Amri, I.; Lee, I.J.; Khan, A.L. Silicon and salicylic acid confer high-pH stress tolerance in tomato seedlings. Sci. Rep. 2019, 9, 19788. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, N.; Ma, J.F. Spatial distribution and temporal variation of the rice silicon transporter Lsi1. Plant Physiol. 2007, 143, 1306–1313. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.X.; Gong, H.J. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Marafon, A.C.; Endres, L. Silicon: Fertilization and nutrition in higher plants. Rev. Ciênc. Agrár. Amazon. J. Agric. Environ. Sci. 2013, 56, 380–388. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.F.; Xiao, X.M.; Dong, Z.X.; Chen, Y. Silicon effects on antioxidative enzymes and lipid peroxidation in leaves and roots of peanut under aluminum stress. Acta Physiol. Plant. 2014, 36, 3063–3069. [Google Scholar] [CrossRef]
- Van Bockhaven, J.; De Vleesschauwer, D.; Hofte, M. Towards establishing broad-spectrum disease resistance in plants: Silicon leads the way. J. Exp. Bot. 2013, 64, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Sivanesan, I.; Jeong, B.R. Silicon promotes adventitious shoot regeneration and enhances salinity tolerance of Ajuga multiflora Bunge by altering activity of antioxidant enzyme. Sci. World J. 2014, 2014, 521703. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Khan, A.L.; Lee, I.J. Silicon: A duo synergy for regulating crop growth and hormonal signaling under abiotic stress conditions. Crit. Rev. Biotechnol. 2016, 36, 1099–1109. [Google Scholar] [CrossRef]
- Seebold, K.W.; Datnoff, L.E.; Correa-Victoria, F.J.; Kucharek, T.A.; Snyder, G.H. Effect of silicon rate and host resistance on blast, scald, and yield of upland rice. Plant Dis. 2000, 84, 871–876. [Google Scholar] [CrossRef]
- Datnoff, L.E.; Raid, R.N.; Snyder, G.H.; Jones, D.B. Effect of calcium silicate on blast and brown spot intensities and yields of rice. Plant Dis. 1991, 75, 729–732. [Google Scholar] [CrossRef]
- Rodrigues, F.A.; Datnoff, L.E.; Korndorfer, G.H.; Seebold, K.W.; Rush, M.C. Effect of silicon and host resistance on sheath blight development in rice. Plant Dis. 2001, 85, 827–832. [Google Scholar]
- Zhan, L.P.; Peng, D.L.; Wang, X.L.; Kong, L.A.; Peng, H.; Liu, S.M.; Liu, Y.; Huang, W.K. Priming effect of root-applied silicon on the enhancement of induced resistance to the root-knot nematode Meloidogyne graminicola in rice. BMC Plant Biol. 2018, 18. [Google Scholar] [CrossRef] [Green Version]
- Phuong, N.T.K.; Khoi, C.M.; Ritz, K.; Linh, T.B.; Minh, D.D.; Duc, T.A.; Sinh, N.V.; Linh, T.T.; Toyota, K. Influence of rice husk biochar and compost amendments on salt contents and hydraulic properties of soil and rice yield in salt-affected fields. Agronomy 2020, 10, 1101. [Google Scholar] [CrossRef]
- Cobb, N.A. Estimating the Nema Populations of Soil; Bureau of Plant Industrial, US Department of Agricultural: Washington, DC, USA, 1918; Volume 1, p. 48.
- Thistlethwayte, B.; Riedel, R.M. Expressing sucrose concentration in solutions used for extracting nematodes. J. Nematol. 1969, 1, 387–388. [Google Scholar] [PubMed]
- De Grisse, A. Redescription ou modifications de quelques techniques utililisées dans l’etude des nematodes phytoparasitaires. Meded. Rijksfac. Landbouwwet. Gent 1969, 34, 351–369. [Google Scholar]
- Yeates, G.W.; Bongers, T.; Degoede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding-habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Clarke, K.R.; Gorley, R.N. Primer v6: User Manual/Tutorial (Plymouth Routines in Multivariate Ecological Research); PRIMER-E Ltd.: Plymouth, UK, 2006. [Google Scholar]
- Sieriebriennikov, B.; Ferris, H.; de Goede, R.G.M. Ninja: An automated calculation system for nematode-based biological monitoring. Eur. J. Soil Biol. 2014, 61, 90–93. [Google Scholar] [CrossRef]
- Ferris, H. Form and function: Metabolic footprints of nematodes in the soil food web. Eur. J. Soil Biol. 2010, 46, 97–104. [Google Scholar] [CrossRef]
- Sanchez-Moreno, S.; Nicola, N.L.; Ferris, H.; Zalom, F.G. Effects of agricultural management on nematode-mite assemblages: Soil food web indices as predictors of mite community composition. Appl. Soil Ecol. 2009, 41, 107–117. [Google Scholar] [CrossRef]
- Ciobanu, M.; Popovici, I.; Zhao, J.; Stoica, I.A. Patterns of relative magnitudes of soil energy channels and their relationships with environmental factors in different ecosystems in Romania. Sci. Rep. 2015, 5, 17606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legendre, L.; Legendre, P. Numerical ecology. J. Environ. Manag. 1987, 24, 202–203. [Google Scholar]
- Martinez, J.G.; Torres, M.A.; dos Santos, G.; Moens, T. Influence of heavy metals on nematode community structure in deteriorated soil by gold mining activities in Sibutad, Southern Philippines. Ecol. Indic. 2018, 91, 712–721. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. Permanova+ for Primer: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Bert, W.; Borgonie, G.; Traunspurger, W.; Andrássy, I. Order Rhabditida: Sub Order Tylenchina, Freshwater Nematodes: Ecology and Taxonomy; CAB International Publishing: London, UK, 2006; pp. 648–695. [Google Scholar]
- Liu, T.; Whalen, J.K.; Shen, Q.R.; Li, H.X. Increase in soil nematode abundance due to fertilization was consistent across moisture regimes in a paddy rice-upland wheat system. Eur. J. Soil Biol. 2016, 72, 21–26. [Google Scholar] [CrossRef]
- Linh, T.B.; Van, K.L.; Van Elsacker, S.; Cornelis, W.M. Effect of cropping system on physical properties of clay soil under intensive rice cultivation. Land Degrad. Dev. 2016, 27, 973–982. [Google Scholar] [CrossRef]
- Pan, F.J.; Han, X.Z.; Li, N.; Yan, J.; Xu, Y.L. Effect of organic amendment amount on soil nematode community structure and metabolic footprints in soybean phase of a soybean-maize rotation on Mollisols. Pedosphere 2020, 30, 544–554. [Google Scholar] [CrossRef]
- Herren, G.L.; Habraken, J.; Waeyenberge, L.; Haegeman, A.; Viaene, N.; Cougnon, M.; Reheul, D.; Steel, H.; Bert, W. Effects of synthetic fertilizer and farm compost on soil nematode community in long-term crop rotation plots: A morphological and metabarcoding approach. PLoS ONE 2020, 15, e0230153. [Google Scholar] [CrossRef]
- Renco, M.; Sasanelli, N.; D’Addabbo, T.; Papajova, I. Soil nematode community changes associated with compost amendments. Nematology 2010, 12, 681–692. [Google Scholar] [CrossRef]
- Yang, L.; Han, Y.Q.; Li, P.; Li, F.; Ali, S.; Hou, M.L. Silicon amendment is involved in the induction of plant defense responses to a phloem feeder. Sci. Rep. 2017, 7, 4232. [Google Scholar] [CrossRef] [PubMed]
- Frew, A.; Powell, J.R.; Hiltpold, I.; Allsopp, P.G.; Sallam, N.; Johnson, S.N. Host plant colonisation by arbuscular mycorrhizal fungi stimulates immune function whereas high root silicon concentrations diminish growth in a soil-dwelling herbivore. Soil Biol. Biochem. 2017, 112, 117–126. [Google Scholar] [CrossRef]
- Quist, C.W.; Gort, G.; Mooijman, P.; Brus, D.J.; van den Elsen, S.; Kostenko, O.; Vervoort, M.; Bakker, J.; van der Putten, W.H.; Helder, J. Spatial distribution of soil nematodes relates to soil organic matter and life strategy. Soil Biol. Biochem. 2019, 136, 107542. [Google Scholar] [CrossRef]
- Epstein, E. Silicon. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Fang, Z.P.; Liang, Y.Q.; Huang, X.H.; Yang, X.; Chen, S.S.; Xie, X.M.; Xu, C.X.; Guo, J.W. Effects of supplying silicon nutrient on utilization rate of nitrogen and phosphorus nutrients by rice and its soil ecological mechanism in a hybrid rice double-cropping system. J. Zhejiang Univ. Sci. B 2020, 21, 474–484. [Google Scholar] [CrossRef]
- Yool, A.; Tyrrell, T. Role of diatoms in regulating the ocean’s silicon cycle. Global Biogeochem. Cycles 2003, 17. [Google Scholar] [CrossRef]
- Martin-Jezequel, V.; Hildebrand, M.; Brzezinski, M.A. Silicon metabolism in diatoms: Implications for growth. J. Phycol. 2000, 36, 821–840. [Google Scholar] [CrossRef]
- Whitton, B.A.; Aziz, A.; Kawecka, B.; Rother, J.A. Ecology of deepwater rice-fields in Bangladesh 3. Associated algae and macrophytes. Hydrobiologia 1988, 169, 31–42. [Google Scholar] [CrossRef]
- Fujita, Y.; Nakahara, H. Variations in the microalgal structure in paddy soil in Osaka, Japan: Comparison between surface and subsurface soils. Limnology 2006, 7, 83–91. [Google Scholar] [CrossRef]
- Nche-Fambo, F.A.; Scharler, U.M.; Tirok, K. Resilience of estuarine phytoplankton and their temporal variability along salinity gradients during drought and hypersalinity. Estuar. Coast. Shelf Sci. 2015, 158, 40–52. [Google Scholar] [CrossRef]
- Sun, J.H.; Li, W.W.; Wu, N.H.; Lai, Z.N.; Du, W.L.; Jia, H.J.; Ge, D.Y.; Wang, C. Spatio-temporal patterns and predictions of size-fractionated chlorophyll a in a large subtropical river, China. J. Freshwater Ecol. 2020, 35, 1–12. [Google Scholar] [CrossRef]
- Yen, N.T.M.; Vanreusel, A.; Lins, L.; Thai, T.T.; Bezerra, T.N.; Quang, N.X. The effect of a dam construction on subtidal nematode communities in the Ba Lai estuary, Vietnam. Diversity 2020, 12, 137. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.A. Historic drought and salinity intrusion in the Mekong delta in 2016: Lessons learned and response solutions. Vietnam J. Sci. Technol. Eng. 2017, 59, 93–96. [Google Scholar] [CrossRef]
- Sohlenius, B. Influence of climatic conditions on nematode coexistence-a laboratory experiment with a coniferous forest soil. Oikos 1985, 44, 430–438. [Google Scholar] [CrossRef]
- Majdi, N.; Traunspurger, W. Free-living nematodes in the freshwater food web: A review. J. Nematol. 2015, 47, 28–44. [Google Scholar]
- Lazarova, S.; Coyne, D.; Rodríguez, M.G.; Peteira, B.; Ciancio, A. Functional diversity of soil nematodes in relation to the impact of agriculture—A review. Diversity 2021, 13, 64. [Google Scholar] [CrossRef]
- Das, S.; Gwon, H.S.; Khan, M.I.; Van Nostrand, J.D.; Alam, M.A.; Kim, P.J. Taxonomic and functional responses of soil microbial communities to slag-based fertilizer amendment in rice cropping systems. Environ. Int. 2019, 127, 531–539. [Google Scholar] [CrossRef]
- Das, S.; Lee, J.G.; Cho, S.R.; Song, H.J.; Kim, P.J. Silicate fertilizer amendment alters fungal communities and accelerates soil organic matter decomposition. Front. Microbiol. 2019, 10, 2950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.F.; Fu, H.L.; Traore, S. Biodiversity conservation in rice paddies in China: Toward ecological sustainability. Sustainability 2014, 6, 6107–6124. [Google Scholar] [CrossRef] [Green Version]
- Siebert, J.; Ciobanu, M.; Schadler, M.; Eisenhauer, N. Climate change and land use induce functional shifts in soil nematode communities. Oecologia 2020, 192, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Kardol, P.; Thakur, M.P.; Gruss, I.; Wu, G.L.; Eisenhauer, N.; Schadler, M. Soil functional biodiversity and biological quality under threat: Intensive land use outweighs climate change. Soil Biol. Biochem. 2020, 147, 107847. [Google Scholar] [CrossRef]
- Vazquez, C.; de Goede, R.G.M.; Korthals, G.W.; Rutgers, M.; Schouten, A.J.; Creamer, R. The effects of increasing land use intensity on soil nematodes: A turn towards specialism. Funct. Ecol. 2019, 33, 2003–2016. [Google Scholar] [CrossRef] [Green Version]
- Asiedu, O.; Kwoseh, C.K.; Melakeberhan, H.; Adjei-Gyapong, T. Nematode distribution in cultivated and undisturbed soils of Guinea Savannah and Semi-deciduous Forest zones of Ghana. Geosci. Front. 2019, 10, 381–387. [Google Scholar] [CrossRef]
- Gregory, A.S.; Ritz, K.; McGrath, S.P.; Quinton, J.N.; Goulding, K.W.T.; Jones, R.J.A.; Harris, J.A.; Bol, R.; Wallace, P.; Pilgrim, E.S.; et al. A review of the impacts of degradation threats on soil properties in the UK. Soil Use Manag. 2015, 31, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Thoden, T.C.; Korthals, G.W.; Termorshuizen, A.Z. Organic amendments and their influences on plant-parasiticand free-living nematodes: A promising method fornematode management? Nematology 2011, 13, 133–153. [Google Scholar] [CrossRef]
- Zhong, S.; Zeng, H.C. Effect of peanut (Arachis hypogaea L.)/cowpea (Vigna unquiculata L.) intercropping combined with organic mature application on soil microfauna. Geoderma 2019, 354, 113863. [Google Scholar] [CrossRef]
- Sys, C.; Van Ranst, E.; Debaveye, J.; Beernaert, F. Land Evaluation Part III, Crop Requirements; Greenville Area Development Corporation (GADC): Brussels, Belgium, 1993. [Google Scholar]
- Watanabe, T.; Luu, H.M.; Inubushi, K. Effects of the continuous application of rice straw compost and chemical fertilizer on soil carbon and available silicon under a double rice cropping system in the Mekong delta, Vietnam. Jpn. Agric. Res. Q. 2017, 51, 233–239. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil | SM | pH1:2.5 | EC1:2.5 | Total C | Total N | C/N | Na+ | K+ | Ca2+ | ESP |
|---|---|---|---|---|---|---|---|---|---|---|
| Acid sulfate soil | 29 ± 1.5 b | 5.0 ± 0.1 | 1.18 ± 0.04 a | 18.3 ± 0.5 b | 1.7 ± 0.02 b | 10.6 ± 0.2 | 681 ± 26 a | 38.3 ± 1.4 a | 172 ± 0.4 a | 19.2 |
| Alluvial soil | 36 ± 0.6 a | 4.9 ± 0.1 | 0.56 ± 0.04 b | 22.8 ± 1.1 a | 2.0 ± 0.04 a | 11.4 ± 0.3 | 294 ± 25 b | 31.2 ± 1.5 b | 10.9 ± 2.3 b | 8.8 |
| p-value | ** | ns | *** | ** | ** | ns | *** | * | *** |
| Treatments | Cropping Sequence | Amendment (Amount Crop−1) | Cultivation Seasons | ||||
|---|---|---|---|---|---|---|---|
| 2018 | 2019 | ||||||
| SS | SA | WS | SS | SA | |||
| (Feb–May) | (Jun–Sep) | (Oct–Jan) | (Feb–May) | (Jun–Sep) | |||
| RRR | R − R − R − R − R − R | No | R− | R− | R− | R− | R− |
| FRR | F − R − R − F − R − R | No | F | R− | R− | F | R− |
| FRR + Compost | F − R − R − F − R − R | Compost (3 Mg ha−1) | F | R+ | R+ | F | R+ |
| FRR + Si | F − R − R − F − R − R | Silicate fertilizer (100 kg ha−1) | F | R+ | R+ | F | R+ |
| RRR | FRR | FRR + Compost | FRR + Si | |
|---|---|---|---|---|
| Acid sulfate soil | ||||
| RRR | Hirschmanniella (82.9%) | |||
| Mesodorylaimus (15.7%) | ||||
| FRR | Hirschmanniella (58.8%) | |||
| Mesodorylaimus(22.7%) | Hirschmanniella (51.2%) | |||
| Rhabdolaimus (6.4%) | Mesodorylaimus (36.2%) | |||
| FRR + Compost | Hirschmanniella (50.7%) | Mesodorylaimus(36.6%) | ||
| Mesodorylaimus(21.5%) | Hirschmanniella (23.2%) | Hirschmanniella (51.5%) | ||
| Rhabdolaimus (10.6%) | Rhabdolaimus (16.3%) | Mesodorylaimus (36.5%) | ||
| Filenchus (5.7%) | ||||
| FRR + Si | Mesodorylaimus (38.0%) | Mesodorylaimus (47.8%) | Mesodorylaimus (45.5%) | Hirschmanniella (48.2%) |
| Hirschmanniella(33.8%) | Rhabdolaimus (15.7%) | Rhabdolaimus (14.9%) | Mesodorylaimus (28.2%) | |
| Rhabdolaimus (12.6%) | Hirschmanniella(11.2%) | Hirschmanniella(12.1%) | Rhabdolaimus (14.2%) | |
| Dorylaims (7.0%) | Dorylaims (7.2%) | |||
| Alluvial soil | ||||
| RRR | Hirschmanniella (82.7%) | |||
| Chronogaster (8.0%) | ||||
| FRR | Hirschmanniella (44.2%) | Hirschmanniella (67.7%) | ||
| Filenchus (16.1%) | Filenchus (12.7%) | |||
| Chronogaster (12.8%) | Chronogaster (10.3%) | |||
| Acrobeloides (5.7%) | ||||
| FRR + Compost | Hirschmanniella (58.6%) | Hirschmanniella (33.4%) | Hirschmanniella (73.1%) | |
| Chronogaster (10.2%) | Filenchus (21.2%) | Chronogaster (11.4%) | ||
| Filenchus (5.4%) | Chronogaster (16.2%) | Filenchus (7.6%) | ||
| Acrobeloides (6.6%) | ||||
| FRR + Si | Hirschmanniella (53.4%) | Hirschmanniella (34.8%) | Hirschmanniella (29.0%) | |
| Chronogaster (12.9%) | Filenchus (18.9%) | Filenchus (17.5%) | Hirschmanniella (77.2%) | |
| Filenchus (8.4%) | Chronogaster (18.3%) | Chronogaster (12.7%) | Filenchus (12.0%) | |
| Acrobeloides (6.6%) | Bitylenchus (7.9%) | Chronogaster (5.9%) | ||
| Bitylenchus (5.8%) | Acrobeloides (5.6%) | |||
| Ditylenchus (5.3%) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinh, N.V.; Khoi, C.M.; Phuong, N.T.K.; Linh, T.B.; Minh, D.D.; Perry, R.N.; Toyota, K. Impacts of Fallow Conditions, Compost and Silicate Fertilizer on Soil Nematode Community in Salt–Affected Paddy Rice Fields in Acid Sulfate and Alluvial Soils in the Mekong Delta, Vietnam. Agronomy 2021, 11, 425. https://doi.org/10.3390/agronomy11030425
Sinh NV, Khoi CM, Phuong NTK, Linh TB, Minh DD, Perry RN, Toyota K. Impacts of Fallow Conditions, Compost and Silicate Fertilizer on Soil Nematode Community in Salt–Affected Paddy Rice Fields in Acid Sulfate and Alluvial Soils in the Mekong Delta, Vietnam. Agronomy. 2021; 11(3):425. https://doi.org/10.3390/agronomy11030425
Chicago/Turabian StyleSinh, Nguyen Van, Chau Minh Khoi, Nguyen Thi Kim Phuong, Tran Ba Linh, Dang Duy Minh, Roland N. Perry, and Koki Toyota. 2021. "Impacts of Fallow Conditions, Compost and Silicate Fertilizer on Soil Nematode Community in Salt–Affected Paddy Rice Fields in Acid Sulfate and Alluvial Soils in the Mekong Delta, Vietnam" Agronomy 11, no. 3: 425. https://doi.org/10.3390/agronomy11030425
APA StyleSinh, N. V., Khoi, C. M., Phuong, N. T. K., Linh, T. B., Minh, D. D., Perry, R. N., & Toyota, K. (2021). Impacts of Fallow Conditions, Compost and Silicate Fertilizer on Soil Nematode Community in Salt–Affected Paddy Rice Fields in Acid Sulfate and Alluvial Soils in the Mekong Delta, Vietnam. Agronomy, 11(3), 425. https://doi.org/10.3390/agronomy11030425