
Influence of Rice Husk Biochar on Soil Nematode Community under Upland and Flooded Conditions: A Microcosm Experiment

 , and
, and 
Abstract
:1. Introduction
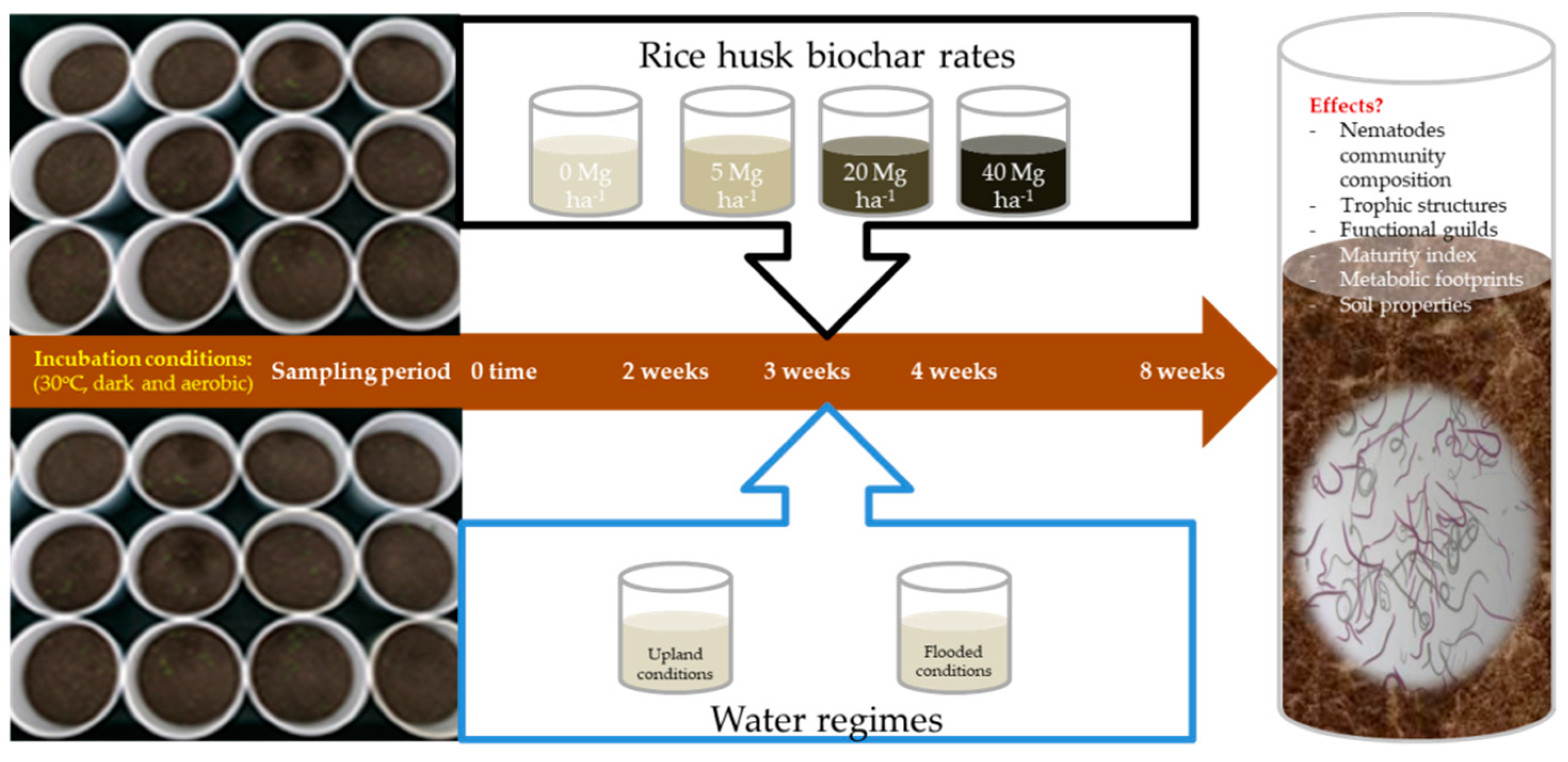
2. Materials and Methods
2.1. Soil and Biochar Preparations
2.2. Biochar-Amended Soil
2.3. Sampling
2.4. Soil Physico-Chemical Properties Analysis
2.5. Microbial Activity Measurement
2.6. Nematode Community Assessment
2.7. Statistics
3. Results
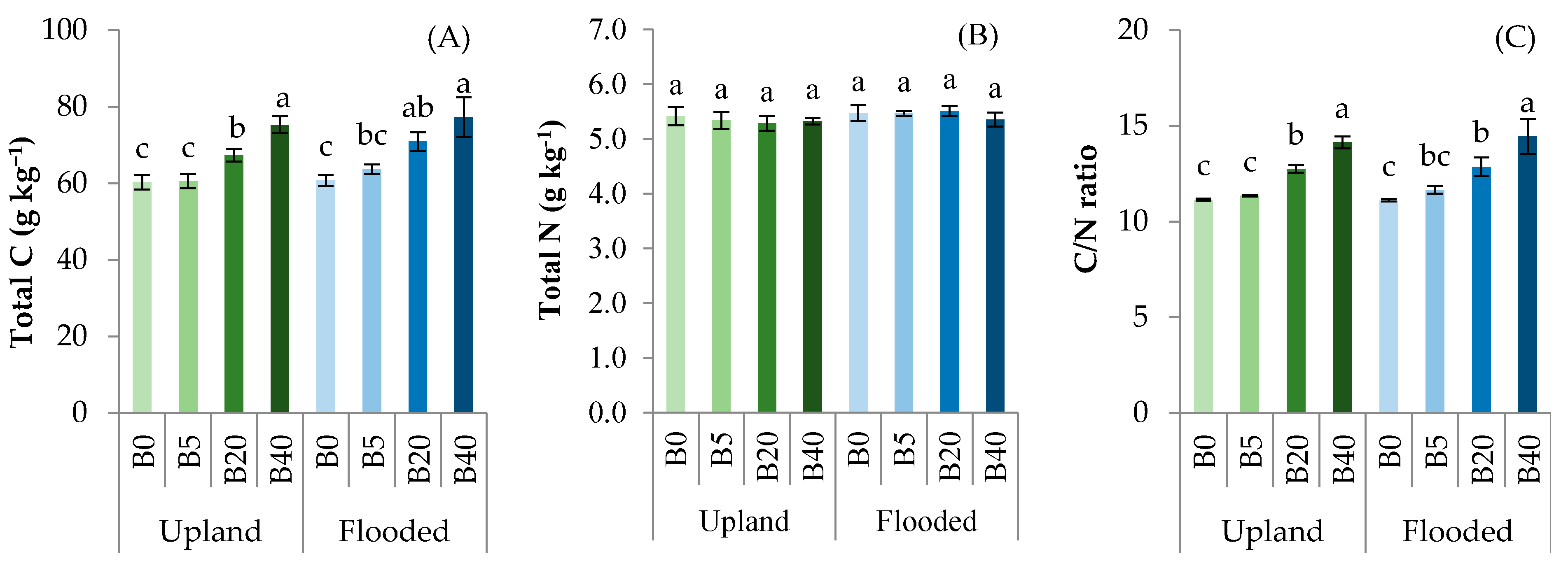
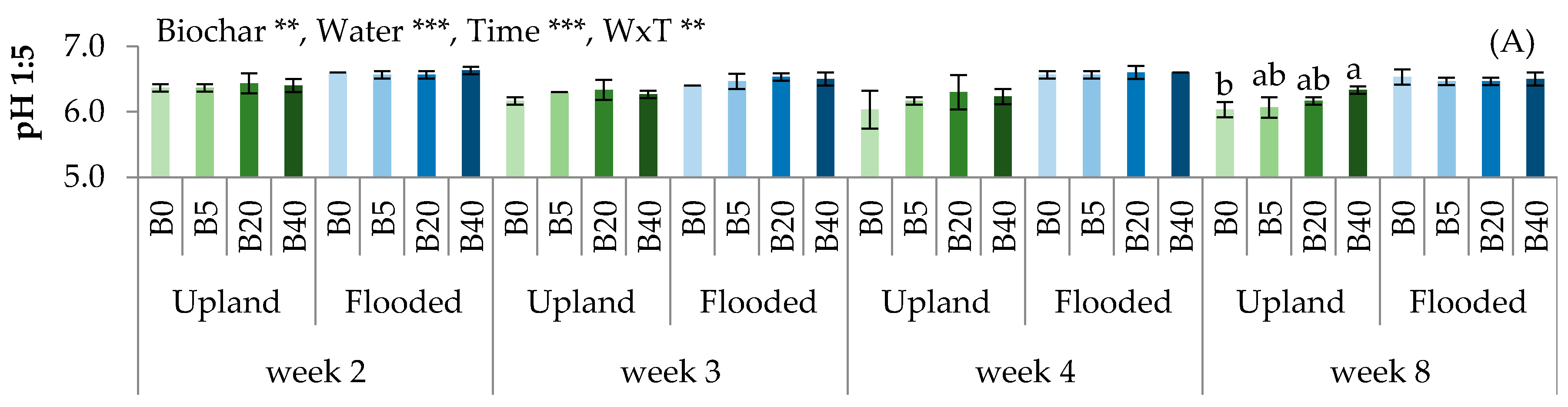
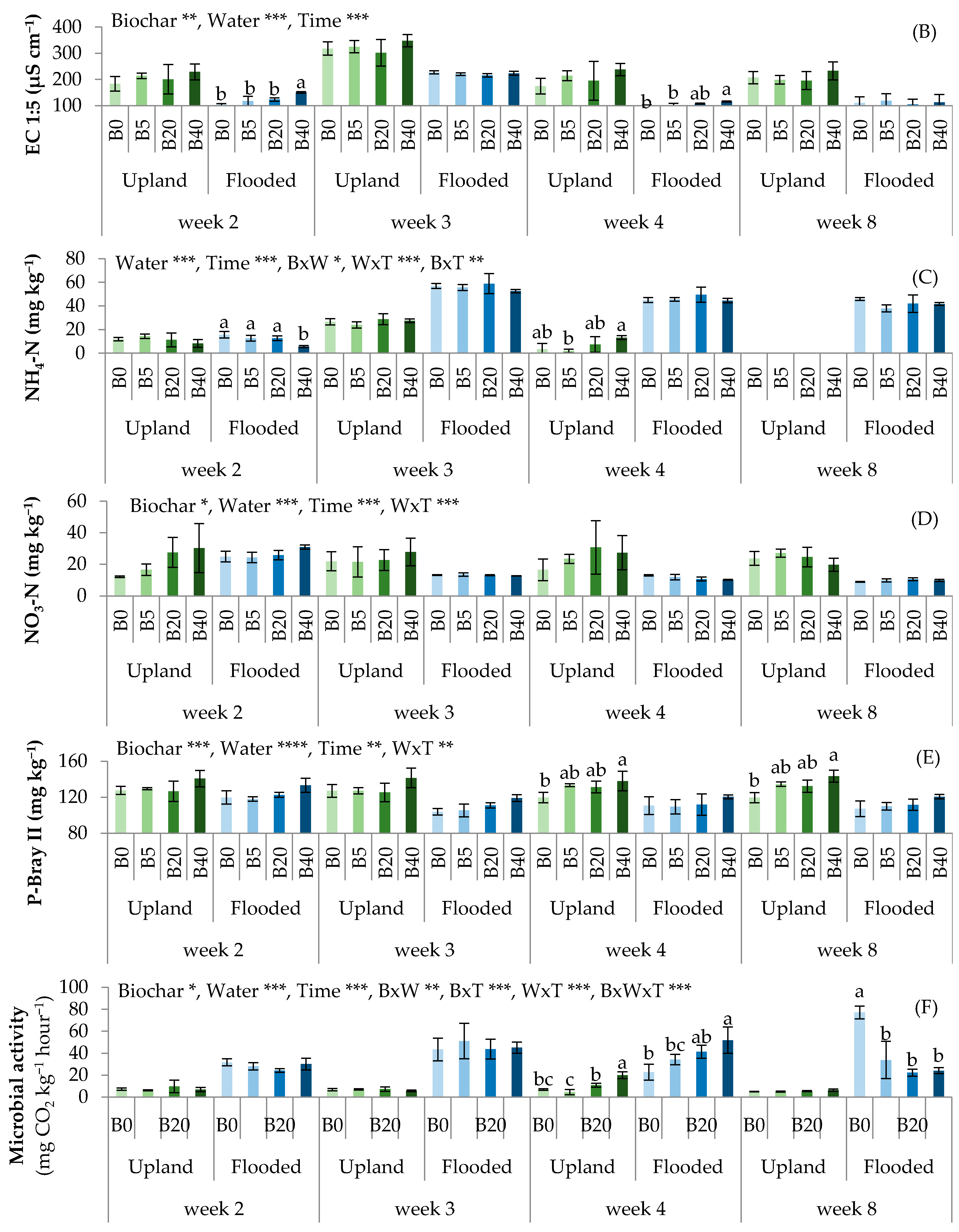
3.1. Biochar Affected Soil Physico-Chemical and Biological Properties
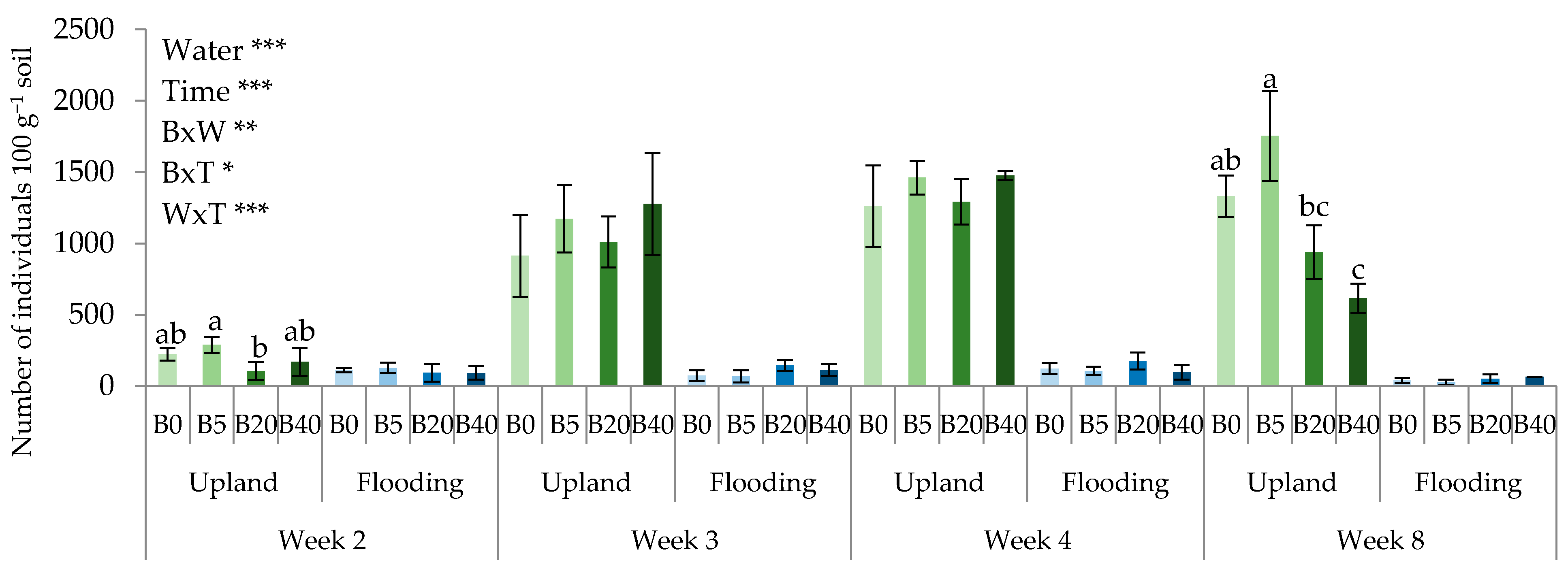
3.2. Biochar Affected Abundance of Nematodes
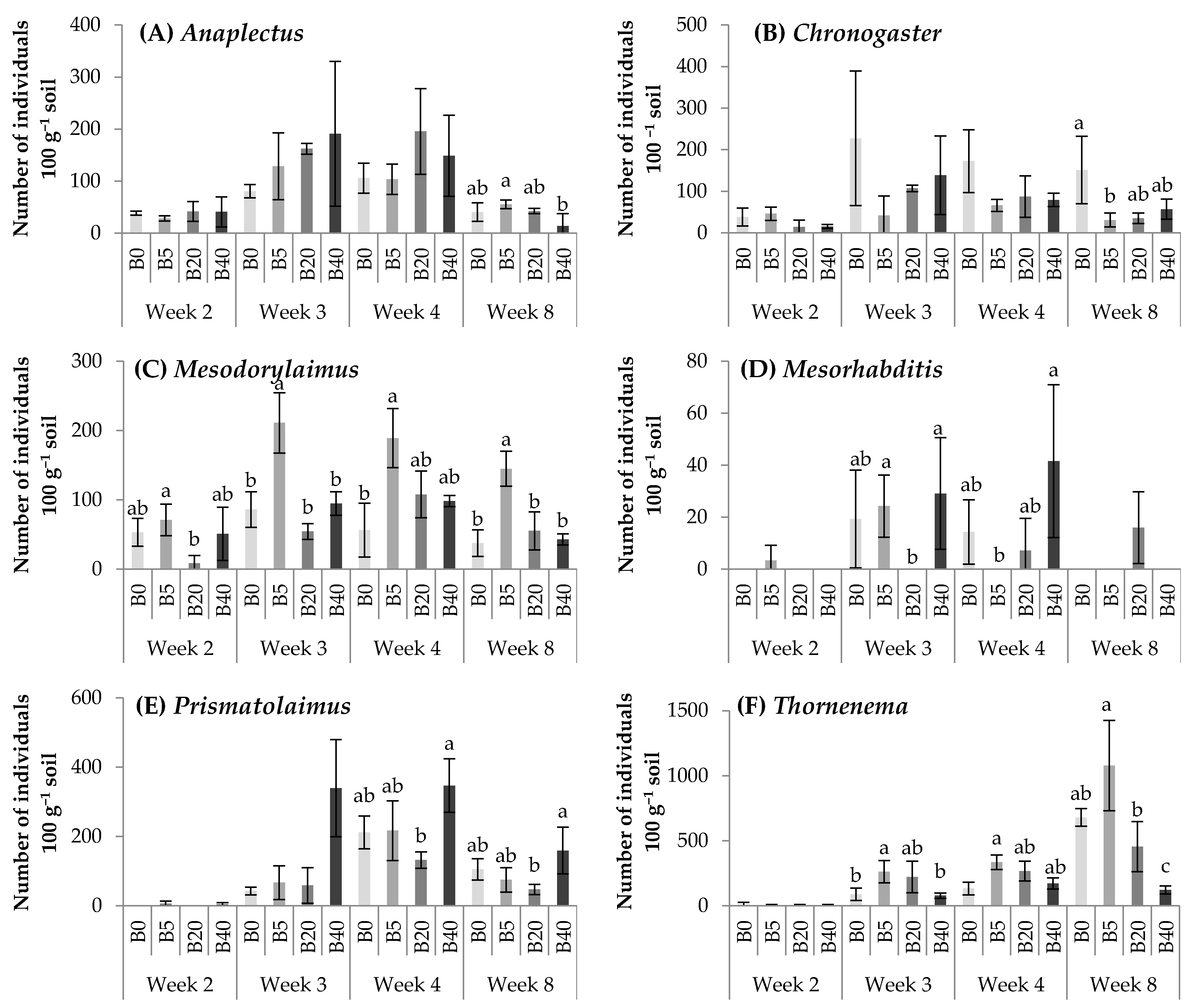
3.3. Biochar Affected Nematodes Community Composition
3.4. Biochar Affected Trophic Structure and Functional Guilds
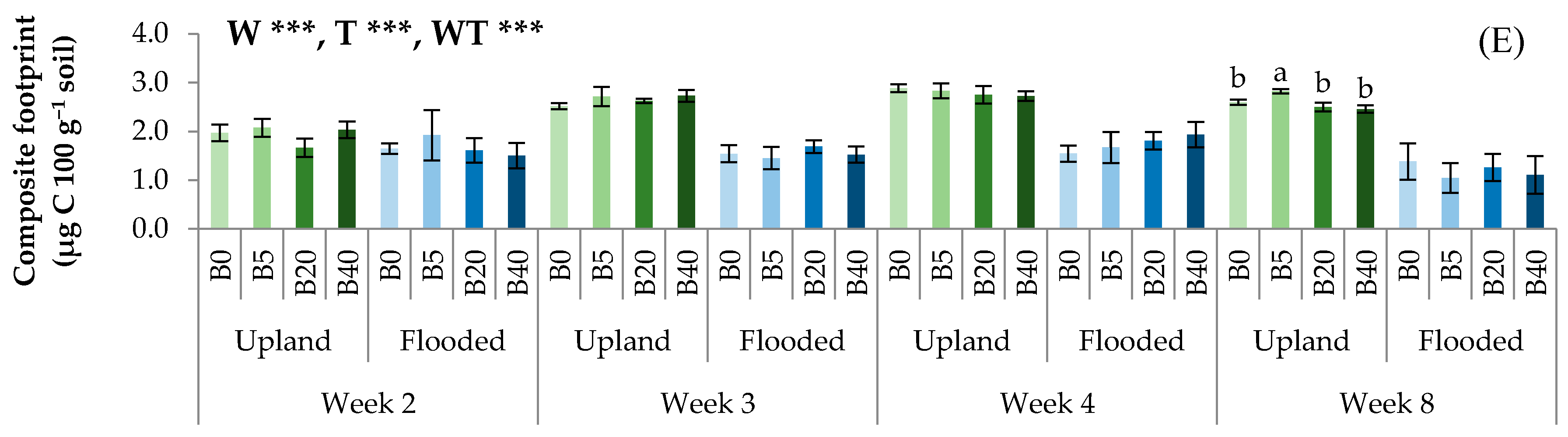
3.5. Biochar Affected Maturity Index, Total Biomass and Metabolic Footprint
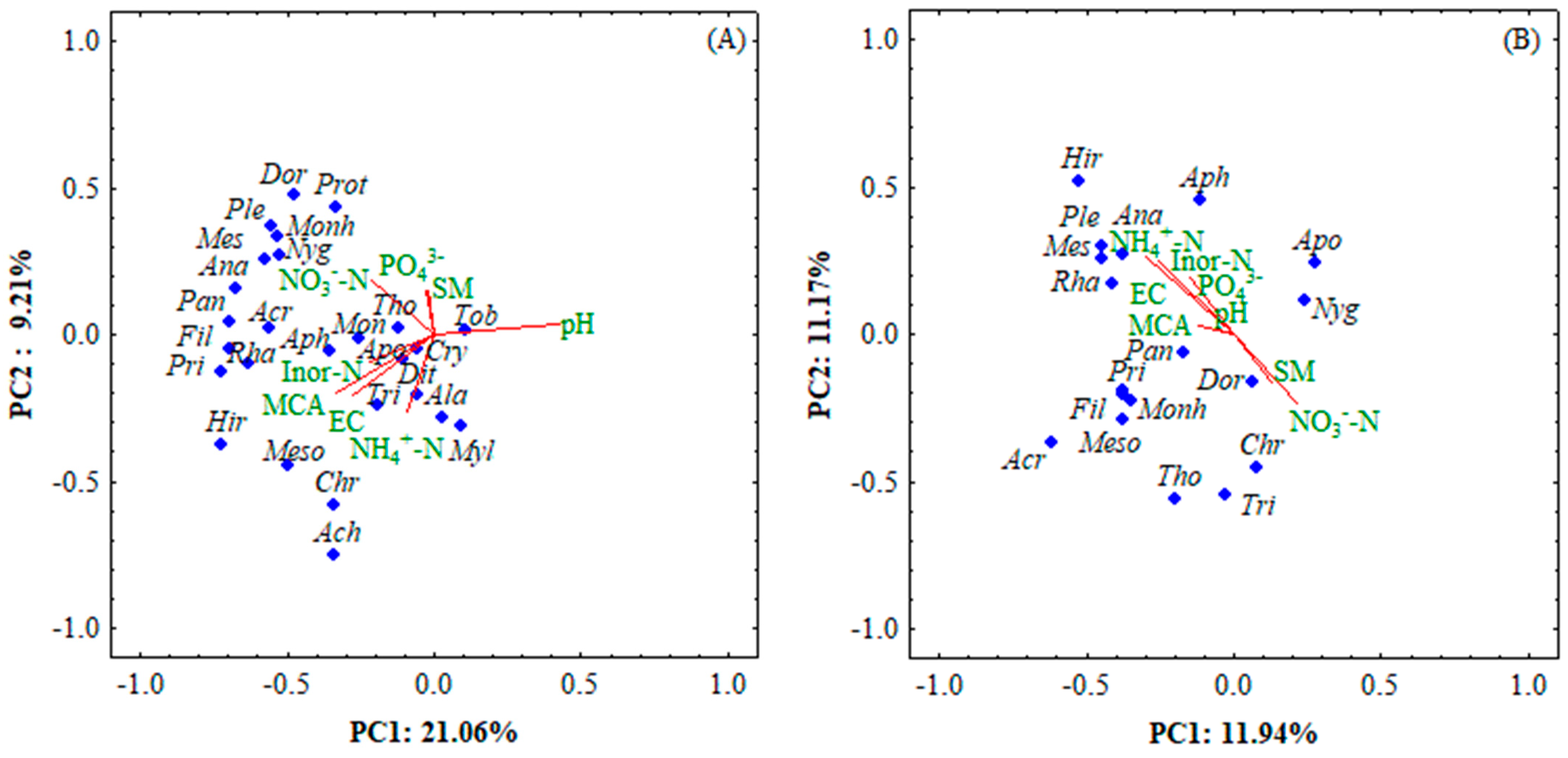
3.6. Relationship between Nematodes Community Composition and Soil Properties
4. Discussion
4.1. Effects of Biochar on Soil Physico-Chemical Properties
4.2. Effects of Biochar on Nematode Community Composition upon Water Regimes
4.3. Effects of Biochar on Nematode Trophic Structure and Functional Guilds
4.4. Effects of Biochar on Maturity Index and Metabolic Footprint of Nematode Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Palansooriya, K.N.; Ok, Y.S.; Awad, Y.M.; Lee, S.S.; Sung, J.K.; Koutsospyros, A.; Moon, D.H. Impacts of biochar application on upland agriculture: A review. J. Environ. Manag. 2019, 234, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, B.J.; Zhang, Y.H.; Lin, Z.B.; Zhu, T.B.; Sun, R.B.; Wang, X.J.; Ma, J.; Bei, Q.C.; Liu, G.; et al. Can biochar alleviate soil compaction stress on wheat growth and mitigate soil N2O emissions? Soil Biol. Biochem. 2017, 104, 8–17. [Google Scholar] [CrossRef]
- Dempster, D.N.; Gleeson, D.B.; Solaiman, Z.M.; Jones, D.L.; Murphy, D.V. Decreased soil microbial biomass and nitrogen mineralisation with Eucalyptus biochar addition to a coarse textured soil. Plant Soil 2012, 354, 311–324. [Google Scholar] [CrossRef]
- Kamau, S.; Karanja, N.K.; Ayuke, F.O.; Lehmann, J. Short-term influence of biochar and fertilizer-biochar blends on soil nutrients, fauna and maize growth. Biol. Fertil. Soils 2019, 55, 661–673. [Google Scholar] [CrossRef]
- Liu, S.N.; Meng, J.; Jiang, L.L.; Yang, X.; Lan, Y.; Cheng, X.Y.; Chen, W.F. Rice husk biochar impacts soil phosphorous availability, phosphatase activities and bacterial community characteristics in three different soil types. Appl. Soil Ecol. 2017, 116, 12–22. [Google Scholar] [CrossRef]
- Phuong, N.T.K.; Khoi, C.M.; Ritz, K.; Linh, T.B.; Minh, D.D.; Duc, T.A.; Sinh, N.V.; Linh, T.T.; Toyota, K. Influence of rice husk biochar and compost amendments on salt contents and hydraulic properties of soil and rice yield in salt-affected fields. Agronomy 2020, 10, 1101. [Google Scholar] [CrossRef]
- Kumar, S.S.; Kumar, A.; Singh, S.; Malyan, S.K.; Baram, S.; Sharma, J.; Singh, R.; Pugazhendhi, A. Industrial wastes: Fly ash, steel slag and phosphogypsum- potential candidates to mitigate greenhouse gas emissions from paddy fields. Chemosphere 2020, 241, 124824. [Google Scholar] [CrossRef]
- Oni, B.A.; Oziegbe, O.; Olawole, O.O. Significance of biochar application to the environment and economy. Ann. Agric. Sci. 2019, 64, 222–236. [Google Scholar] [CrossRef]
- Gomez, J.D.; Denef, K.; Stewart, C.E.; Zheng, J.; Cotrufo, M.F. Biochar addition rate influences soil microbial abundance and activity in temperate soils. Eur. J. Soil Sci. 2014, 65, 28–39. [Google Scholar] [CrossRef]
- Brtnicky, M.; Datta, R.; Holatko, J.; Bielska, L.; Gusiatin, Z.M.; Kucerik, J.; Hammerschmiedt, T.; Danish, S.; Radziemska, M.; Mravcova, L.; et al. A critical review of the possible adverse effects of biochar in the soil environment. Sci. Total Environ. 2021, 796, 148756. [Google Scholar] [CrossRef]
- Liu, T.; Yang, L.H.; Hu, Z.K.; Xue, J.R.; Lu, Y.Y.; Chen, X.Y.; Griffiths, B.S.; Whalen, J.K.; Liu, M.Q. Biochar exerts negative effects on soil fauna across multiple trophic levels in a cultivated acidic soil. Biol. Fertil. Soils 2020, 56, 597–606. [Google Scholar] [CrossRef]
- Zhang, X.K.; Li, Q.; Liang, W.J.; Zhang, M.; Bao, X.L.; Xie, Z.B. Soil nematode response to biochar addition in a Chinese wheat field. Pedosphere 2013, 23, 98–103. [Google Scholar] [CrossRef]
- Domene, X.; Mattana, S.; Sanchez-Moreno, S. Biochar addition rate determines contrasting shifts in soil nematode trophic groups in outdoor mesocosms: An appraisal of underlying mechanisms. Appl. Soil Ecol. 2021, 158, 103788. [Google Scholar] [CrossRef]
- Marra, R.; Vinale, F.; Cesarano, G.; Lombardi, N.; d’Errico, G.; Crasto, A.; Mazzei, P.; Piccolo, A.; Incerti, G.; Woo, S.L.; et al. Biochars from olive mill waste have contrasting effects on plants, fungi and phytoparasitic nematodes. PLoS ONE 2018, 13, e0198728. [Google Scholar] [CrossRef] [PubMed]
- Briones, M.J.I.; Panzacchi, P.; Davies, C.A.; Ineson, P. Contrasting responses of macro- and meso-fauna to biochar additions in a bioenergy cropping system. Soil Biol. Biochem. 2020, 145, 107803. [Google Scholar] [CrossRef]
- Liu, X.D.; Zhang, D.X.; Li, H.X.; Qi, X.X.; Gao, Y.; Zhang, Y.B.; Han, Y.L.; Jiang, Y.; Li, H. Soil nematode community and crop productivity in response to 5-year biochar and manure addition to yellow cinnamon soil. BMC Ecol. 2020, 20, 39. [Google Scholar] [CrossRef]
- Al-Wabel, M.I.; Hussain, Q.; Usman, A.R.A.; Ahmad, M.; Abduljabbar, A.; Sallam, A.S.; Ok, Y.S. Impact of biochar properties on soil conditions and agricultural sustainability: A review. Land Degrad. Dev. 2018, 29, 2124–2161. [Google Scholar] [CrossRef]
- Majdi, N.; Traunspurger, W.; Fueser, H.; Gansfort, B.; Laffaille, P.; Maire, A. Effects of a broad range of experimental temperatures on the population growth and body-size of five species of free-living nematodes. J. Therm. Biol. 2019, 80, 21–36. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Liu, Y.G.; Liu, S.B.; Li, Z.W.; Tan, X.F.; Huang, X.X.; Zeng, G.M.; Zhou, L.; Zheng, B.H. Biochar to improve soil fertility. A review. Agron. Sustain. Dev. 2016, 36, 36. [Google Scholar] [CrossRef] [Green Version]
- Archidona-Yuste, A.; Wiegand, T.; Castillo, P.; Navas-Cortes, J.A. Spatial structure and soil properties shape local community structure of plant-parasitic nematodes in cultivated olive trees in Southern Spain. Agric. Ecosyst. Environ. 2020, 287, 106688. [Google Scholar] [CrossRef]
- Cardoso, M.S.O.; Pedrosa, E.M.R.; Rolim, M.M.; Oliveira, L.S.C.; Santos, A.N. Relationship between nematode assemblages and physical properties across land use types. Trop. Plant Pathol. 2016, 41, 107–114. [Google Scholar] [CrossRef]
- Kitagami, Y.; Tanikawa, T.; Matsuda, Y. Effects of microhabitats and soil conditions on structuring patterns of nematode communities in Japanese cedar (Cryptomeria japonica) plantation forests under temperate climate conditions. Soil Biol. Biochem. 2020, 151, 108044. [Google Scholar] [CrossRef]
- Jiang, Y.J.; Zhou, H.; Chen, L.J.; Yuan, Y.; Fang, H.; Luan, L.; Chen, Y.; Wang, X.Y.; Liu, M.Q.; Li, H.X.; et al. Nematodes and microorganisms interactively stimulate soil organic carbon turnover in the macroaggregates. Front. Microbiol. 2018, 9, 2803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haegerbaeumer, A.; Hoss, S.; Heininger, P.; Traunspurger, W. Response of nematode communities to metals and PAHs in freshwater microcosms. Ecotoxicol. Environ. Saf. 2018, 148, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Qin, J.; Shi, D.Z.; Zhang, Y.M.; Ji, W.H. Diversity of soil nematodes in areas polluted with heavy metals and polycyclic aromatic hydrocarbons (PAHs) in Lanzhou, China. Environ. Manag. 2009, 44, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Ruzickova, J.; Koval, S.; Raclavska, H.; Kucbel, M.; Svedova, B.; Raclavsky, K.; Juchelkova, D.; Scala, F. A comprehensive assessment of potential hazard caused by organic compounds in biochar for agricultural use. J. Hazard. Mater. 2021, 403, 123644. [Google Scholar] [CrossRef]
- Wang, C.Y.; Wang, Y.D.; Herath, H. Polycyclic aromatic hydrocarbons (PAHs) in biochar—Their formation, occurrence and analysis: A review. Org. Geochem. 2017, 114, 1–11. [Google Scholar] [CrossRef]
- Wang, J.; Odinga, E.S.; Zhang, W.; Zhou, X.; Yang, B.; Waigi, M.G.; Gao, Y. Polyaromatic hydrocarbons in biochars and human health risks of food crops grown in biochar-amended soils: A synthesis study. Environ. Int. 2019, 130, 104899. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T.; de Goede, R.G.M. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Neher, D.A. Nematode communities in organically and conventionally managed agricultural soils. J. Nematol. 1999, 31, 142–154. [Google Scholar]
- Bongers, T. The maturity index-an ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H.; Venette, R.C.; Scow, K.M. Soil management to enhance bacterivore and fungivore nematode populations and their nitrogen mineralisation function. Appl. Soil Ecol. 2004, 25, 19–35. [Google Scholar] [CrossRef]
- Gebremikael, M.T.; Buchan, D.; De Neve, S. Quantifying the influences of free-living nematodes on soil nitrogen and microbial biomass dynamics in bare and planted microcosms. Soil Biol. Biochem. 2014, 70, 131–141. [Google Scholar] [CrossRef]
- Ritz, K.; Trudgill, D.L. Utility of nematode community analysis as an integrated measure of the functional state of soils: Perspectives and challenges—Discussion paper. Plant Soil 1999, 212, 1–11. [Google Scholar] [CrossRef]
- Anderson, R.V.; Coleman, D.C. Nematode temperature responses-a niche dimension in populations of bacterial-feeding nematodes. J. Nematol. 1982, 14, 69–76. [Google Scholar]
- Hu, C.; Qi, Y.C. Effect of compost and chemical fertilizer on soil nematode community in a Chinese maize field. Eur. J. Soil Biol. 2010, 46, 230–236. [Google Scholar] [CrossRef]
- Renco, M.; Sasanelli, N.; D’Addabbo, T.; Papajova, I. Soil nematode community changes associated with compost amendments. Nematology 2010, 12, 681–692. [Google Scholar] [CrossRef]
- Briar, S.S.; Grewal, P.S.; Somasekhar, N.; Stinner, D.; Miller, S.A. Soil nematode community, organic matter, microbial biomass and nitrogen dynamics in field plots transitioning from conventional to organic management. Appl. Soil Ecol. 2007, 37, 256–266. [Google Scholar] [CrossRef]
- Neher, D.A.; Nishanthan, T.; Grabau, Z.J.; Chen, S.Y. Crop rotation and tillage affect nematode communities more than biocides in monoculture soybean. Appl. Soil Ecol. 2019, 140, 89–97. [Google Scholar] [CrossRef]
- Zhang, X.K.; Li, Q.; Zhu, A.N.; Liang, W.J.; Zhang, J.B.; Steinberger, Y. Effects of tillage and residue management on soil nematode communities in North China. Ecol. Indic. 2012, 13, 75–81. [Google Scholar] [CrossRef]
- Bongiorno, G.; Bodenhausen, N.; Bunemann, E.K.; Brussaard, L.; Geisen, S.; Mader, P.; Quist, C.W.; Walser, J.C.; de Goede, R.G.M. Reduced tillage, but not organic matter input, increased nematode diversity and food web stability in european long-term field experiments. Mol. Ecol. 2019, 28, 4987–5005. [Google Scholar] [CrossRef] [Green Version]
- Phuong, N.T.K.; Khoi, C.M.; Ritz, K.; Sinh, N.V.; Tarao, M.; Toyota, K. Potential use of rice husk biochar and compost to improve P availability and reduce GHG emissions in acid sulfate soil. Agronomy 2020, 10, 685. [Google Scholar] [CrossRef]
- Mianowski, A.; Owczarek, M.; Marecka, A. Surface area of activated carbon determined by the iodine adsorption number. Energy Sources A Recovery Util. Environ. Eff. 2007, 29, 839–850. [Google Scholar] [CrossRef]
- Houba, V.J.G.; Vanderlee, J.J.; Novozamsky, I. Soil and plant analysis: A series of syllabi. In Part 5b Soil Analysis Procedures Other Procedures, 6th ed.; Department of Soil Science and Plant Nutrition, Wageningen Agricultural University: Wageningen, The Netherlands, 1995. [Google Scholar]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T.; Degoede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding-habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Sieriebriennikov, B.; Ferris, H.; de Goede, R.G.M. Ninja: An automated calculation system for nematode-based biological monitoring. Eur. J. Soil Biol. 2014, 61, 90–93. [Google Scholar] [CrossRef]
- Ferris, H. Form and function: Metabolic footprints of nematodes in the soil food web. Eur. J. Soil Biol. 2010, 46, 97–104. [Google Scholar] [CrossRef]
- Nguyen, B.T.; Lehmann, J.; Kinyangi, J.; Smernik, R.; Riha, S.J.; Engelhard, M.H. Long-term black carbon dynamics in cultivated soil. Biogeochemistry 2009, 92, 163–176. [Google Scholar] [CrossRef]
- Oladele, S.O. Changes in physicochemical properties and quality index of an alfisol after three years of rice husk biochar amendment in rainfed rice—Maize cropping sequence. Geoderma 2019, 353, 359–371. [Google Scholar] [CrossRef]
- Nielsen, U.N.; Ayres, E.; Wall, D.H.; Li, G.; Bardgett, R.D.; Wu, T.H.; Garey, J.R. Global-scale patterns of assemblage structure of soil nematodes in relation to climate and ecosystem properties. Glob. Ecol. Biogeogr. 2014, 23, 968–978. [Google Scholar] [CrossRef]
- Uhlirova, E.; Elhottova, D.; Triska, J.; Santruckova, H. Physiology and microbial community structure in soil at extreme water content. Folia Microbiol. 2005, 50, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Gebremikael, M.T.; De Waele, J.; Buchan, D.; Soboksa, G.E.; De Neve, S. The effect of varying gamma irradiation doses and soil moisture content on nematodes, the microbial communities and mineral nitrogen. Appl. Soil Ecol. 2015, 92, 1–13. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Dicke, C.; Kalderis, D.; Kern, J. Rice husks and their hydrochars cause unexpected stress response in the nematode Caenorhabditis elegans: Reduced transcription of stress-related genes. Environ. Sci. Pollut. Res. 2015, 22, 12092–12103. [Google Scholar] [CrossRef] [PubMed]
- Tudorancea, C.; Zullini, A. Associations and distribution of benthic nematodes in the Ethiopian rift-valley lakes. Hydrobiologia 1989, 179, 81–96. [Google Scholar] [CrossRef]
- Nguyen, V.S.; Chau, M.K.; Vo, Q.M.; Le, V.K.; Nguyen, T.K.P.; Araki, M.; Perry, R.N.; Tran, A.D.; Dang, D.M.; Tran, B.L.; et al. Impacts of saltwater intrusion on soil nematodes community in alluvial and acid sulfate soils in paddy rice fields in the Vietnamese Mekong Delta. Ecol. Indic. 2021, 122, 107284. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, F.M.; Li, J.; Zou, B.; Wang, X.L.; Li, Z.A.; Fu, S.L. Effects of experimental nitrogen and/or phosphorus additions on soil nematode communities in a secondary tropical forest. Soil Biol. Biochem. 2014, 75, 1–10. [Google Scholar] [CrossRef]
- McSorley, R. Ecology of the dorylaimid omnivore genera Aporcelaimellus, Eudorylaimus and Mesodorylaimus. Nematology 2012, 14, 645–663. [Google Scholar] [CrossRef]
- Quilliam, R.S.; Glanville, H.C.; Wade, S.C.; Jones, D.L. Life in the ‘charosphere’—Does biochar in agricultural soil provide a significant habitat for microorganisms? Soil Biol. Biochem. 2013, 65, 287–293. [Google Scholar] [CrossRef]
- Pietikäinen, J.; Kiikkilä, O.; Fritze, H. Charcoal as a habitat for microbes and its effect on the microbial community of the underlying humus. Oikos 2000, 89, 231–242. [Google Scholar] [CrossRef]
- Okada, H.; Niwa, S.; Takemoto, S.; Komatsuzaki, M.; Hiroki, M. How different or similar are nematode communities between a paddy and an upland rice fields across a flooding-drainage cycle? Soil Biol. Biochem. 2011, 43, 2142–2151. [Google Scholar] [CrossRef]
- Liu, T.; Whalen, J.K.; Shen, Q.; Li, H. Increase in soil nematode abundance due to fertilization was consistent across moisture regimes in a paddy rice–upland wheat system. Eur. J. Soil Biol. 2016, 72, 21–26. [Google Scholar] [CrossRef]
- Randle-Boggis, R.J.; Ashton, P.D.; Helgason, T. Increasing flooding frequency alters soil microbial communities and functions under laboratory conditions. MicrobiologyOpen 2018, 7, e00548. [Google Scholar] [CrossRef]
- Redman, F.H. Effect of Submergence on Several Biological and Chemical Soil Properties; Agricultural Experiment Station, Louisiana State University and Agricultural and Mechanical College: Baton Rouge, LA, USA, 1965. [Google Scholar]
- Ponting, J.; Kelly, T.J.; Verhoef, A.; Watts, M.J.; Sizmur, T. The impact of increased flooding occurrence on the mobility of potentially toxic elements in floodplain soil—A review. Sci. Total Environ. 2021, 754, 142040. [Google Scholar] [CrossRef]
- Unger, I.M.; Kennedy, A.C.; Muzika, R.M. Flooding effects on soil microbial communities. Appl. Soil Ecol. 2009, 42, 1–8. [Google Scholar] [CrossRef]
- Trap, J.; Bonkowski, M.; Plassard, C.; Villenave, C.; Blanchart, E. Ecological importance of soil bacterivores for ecosystem functions. Plant Soil 2016, 398, 1–24. [Google Scholar] [CrossRef]
- Shaw, E.A.; Boot, C.M.; Moore, J.C.; Wall, D.H.; Barone, J.S. Long-term nitrogen addition shifts the soil nematode community to bacterivore-dominated and reduces its ecological maturity in a subalpine forest. Soil Biol. Biochem. 2019, 130, 177–184. [Google Scholar] [CrossRef]
- Wu, J.T.; Zhou, Q.Q.; Huang, R.; Wu, K.J.; Li, Z.A. Contrasting impacts of mobilisation and immobilisation amendments on soil health and heavy metal transfer to food chain. Ecotoxicol. Environ. Saf. 2021, 209, 111836. [Google Scholar] [CrossRef]
- Liu, T.; Chen, X.Y.; Hu, F.; Ran, W.; Shen, Q.R.; Li, H.X.; Whalen, J.K. Carbon-rich organic fertilizers to increase soil biodiversity: Evidence from a meta-analysis of nematode communities. Agric. Ecosyst. Environ. 2016, 232, 199–207. [Google Scholar] [CrossRef]
- Giesselmann, U.C.; Borchard, N.; Traunspurger, W.; Witte, K. Long-term effects of charcoal on nematodes and other soil meso- and microfaunal groups at historical kiln-sites—A pilot study. Eur. J. Soil Biol. 2019, 93, 103095. [Google Scholar] [CrossRef]
- Oladele, S.O.; Adeyemo, A.J.; Awodun, M.A. Influence of rice husk biochar and inorganic fertilizer on soil nutrients availability and rain-fed rice yield in two contrasting soils. Geoderma 2019, 336, 1–11. [Google Scholar] [CrossRef]
- Win, K.T.; Okazaki, K.; Ohkama-Ohtsu, N.; Yokoyama, T.; Ohwaki, Y. Short-term effects of biochar and Bacillus pumilus Tuat-1 on the growth of forage rice and its associated soil microbial community and soil properties. Biol. Fertil. Soils 2020, 56, 481–497. [Google Scholar] [CrossRef]
- Gul, S.; Whalen, J.K.; Thomas, B.W.; Sachdeva, V.; Deng, H.Y. Physico-chemical properties and microbial responses in biochar-amended soils: Mechanisms and future directions. Agric. Ecosyst. Environ. 2015, 206, 46–59. [Google Scholar] [CrossRef]
- Lopes, É.M.G.; Reis, M.M.; Frazão, L.A.; da Mata Terra, L.E.; Lopes, E.F.; dos Santos, M.M.; Fernandes, L.A. Biochar increases enzyme activity and total microbial quality of soil grown with sugarcane. Environ. Technol. Innov. 2021, 21, 101270. [Google Scholar] [CrossRef]
- Wang, J.L.; Wang, S.Z. Preparation, modification and environmental application of biochar: A review. J. Clean. Prod. 2019, 227, 1002–1022. [Google Scholar] [CrossRef]
- Masiello, C.A.; Chen, Y.; Gao, X.D.; Liu, S.; Cheng, H.Y.; Bennett, M.R.; Rudgers, J.A.; Wagner, D.S.; Zygourakis, K.; Silberg, J.J. Biochar and microbial signaling: Production conditions determine effects on microbial communication. Environ. Sci. Technol. 2013, 47, 11496–11503. [Google Scholar] [CrossRef] [Green Version]
- Rafique, M.; Ortas, I.; Ahmed, I.A.M.; Rizwan, M.; Afridi, M.S.; Sultan, T.; Chaudhary, H.J. Potential impact of biochar types and microbial inoculants on growth of onion plant in differently textured and phosphorus limited soils. J. Environ. Manag. 2019, 247, 672–680. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (T) | Water (W) | Biochar (B) | Ba | Fu | Om | Pre | Her |
|---|---|---|---|---|---|---|---|
| Week 2 | Upland | B0 | 2.07 ± 0.04 | 1.12 ± 0.3 ab | 1.77 ± 0.27 ab | 1.06 ± 0.2 | 0.86 ± 0.79 |
| B5 | 2.09 ± 0.19 | 1.31 ± 0.25 a | 1.88 ± 0.13 a | 1.18 ± 1.05 | 1.16 ± 0.15 | ||
| B20 | 1.72 ± 0.29 | 1.14 ± 0.28 ab | 1.21 ± 0.28 b | 0.82 ± 0.76 | 0 ± 0 | ||
| B40 | 1.84 ± 0.30 | 0.32 ± 0.56 b | 1.75 ± 0.25 ab | 0.96 ± 0.83 | 0.76 ± 0.68 | ||
| Flooded | B0 | 1.92 ± 0.15 | 0 ± 0 | 0.37 ± 0.63 | 0.89 ± 0.83 | 0.37 ± 0.63 | |
| B5 | 1.87 ± 0.04 | 0.85 ± 0.74 | 0.95 ± 0.86 | 1.33 ± 0.16 | 0 ± 0 | ||
| B20 | 1.80 ± 0.19 | 0.36 ± 0.62 | 0.45 ± 0.78 | 0.9 ± 0.8 | 0 ± 0 | ||
| B40 | 1.83 ± 0.22 | 0 ± 0 | 0.82 ± 0.71 | 0.34 ± 0.59 | 0.63 ± 0.55 | ||
| Week 3 | Upland | B0 | 2.67 ± 0.16 | 2.00 ± 0.24 | 2.26 ± 0.07 b | 1.90 ± 0.12 | 1.58 ± 0.28 |
| B5 | 2.59 ± 0.11 | 2.14 ± 0.18 | 2.69 ± 0.12 a | 1.77 ± 0.37 | 1.59 ± 0.28 | ||
| B20 | 2.62 ± 0.10 | 2.02 ± 0.52 | 2.44 ± 0.18 ab | 1.87 ± 0.09 | 1.75 ± 0.07 | ||
| B40 | 2.86 ± 0.13 | 2.21 ± 0.41 | 2.27 ± 0.03 b | 1.93 ± 0.23 | 1.59 ± 0.30 | ||
| Flooded | B0 | 1.66 ± 0.19 | 0 ± 0 | 0.8 ± 0.72 | 1.14 ± 0.21 | 0.26 ± 0.45 | |
| B5 | 1.18 ± 1.03 | 0.71 ± 0.62 | 0.81 ± 0.7 | 0.8 ± 0.69 | 0 ± 0 | ||
| B20 | 1.92 ± 0.13 | 1.03 ± 0.90 | 1.39 ± 0.28 | 0.29 ± 0.51 | 0.75 ± 0.65 | ||
| B40 | 1.84 ± 0.07 | 1.27 ± 0.24 | 0.81 ± 0.70 | 0.47 ± 0.82 | 0.33 ± 0.56 | ||
| Week 4 | Upland | B0 | 2.84 ± 0.12 | 2.28 ± 0.13 | 2.42 ± 0.03 b | 1.64 ± 0.16 | 1.18 ± 1.03 |
| B5 | 2.79 ± 0.03 | 2.22 ± 0.12 | 2.75 ± 0.07 a | 1.81 ± 0.22 | 1.60 ± 0.25 | ||
| B20 | 2.79 ± 0.08 | 2.15 ± 0.12 | 2.62 ± 0.1 a | 1.69 ± 0.17 | 1.69 ± 0.17 | ||
| B40 | 2.96 ± 0.05 | 2.19 ± 0.17 | 2.44 ± 0.0 b | 1.72 ± 0.23 | 1.77 ± 0.07 | ||
| Flooded | B0 | 1.94 ± 0.12 | 1.10 ± 0.01 | 0.93 ± 0.85 | 0 ± 0 | 0.37 ± 0.64 | |
| B5 | 1.84 ± 0.24 | 0.37 ± 0.64 | 1.35 ± 0.23 | 0 ± 0 | 0.37 ± 0.64 | ||
| B20 | 2.11 ± 0.18 | 0.52 ± 0.89 | 0.76 ± 0.65 | 0.84 ± 0.75 | 0.74 ± 0.64 | ||
| B40 | 1.64 ± 0.44 | 0.93 ± 0.81 | 0.99 ± 0.86 | 0.37 ± 0.64 | 0 ± 0 | ||
| Week 8 | Upland | B0 | 2.59 ± 0.12 | 2.13 ± 0.15 | 2.85 ± 0.05 ab | 1.62 ± 0.21 | 1.38 ± 0.25 |
| B5 | 2.37 ± 0.16 | 2.17 ± 0.24 | 3.08 ± 0.11 a | 1.98 ± 0.16 | 1.31 ± 0.21 | ||
| B20 | 2.35 ± 0.07 | 1.72 ± 0.17 | 2.69 ± 0.15 b | 1.97 ± 0.45 | 0.88 ± 0.81 | ||
| B40 | 2.47 ± 0.10 | 1.75 ± 0.15 | 2.25 ± 0.05 c | 1.70 ± 0.33 | 0.92 ± 0.86 | ||
| Flooded | B0 | 1.43 ± 0.43 | 0 ± 0 | 0.32 ± 0.55 | 0.32 ± 0.55 | 0 ± 0 | |
| B5 | 1.22 ± 0.21 | 0.28 ± 0.48 | 0.34 ± 0.59 | 0.62 ± 0.54 | 0 ± 0 | ||
| B20 | 1.48 ± 0.41 | 0.38 ± 0.66 | 0 ± 0 | 0.45 ± 0.78 | 0 ± 0 | ||
| B40 | 1.70 ± 0.30 | 0.47 ± 0.81 | 0 ± 0 | 0 ± 0 | 0 ± 0 | ||
| p | B | ns | ns | ns | ns | ns | |
| W | *** | *** | *** | *** | *** | ||
| T | *** | *** | *** | ns | *** | ||
| BxW | ns | ns | ns | ns | ns | ||
| BxT | ns | * | ns | ns | ns | ||
| WxT | *** | ** | *** | *** | ** | ||
| BxWxT | ns | ns | ns | ns | ns | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Sinh, N.; Kato, R.; Linh, D.T.T.; Phuong, N.T.K.; Toyota, K. Influence of Rice Husk Biochar on Soil Nematode Community under Upland and Flooded Conditions: A Microcosm Experiment. Agronomy 2022, 12, 378. https://doi.org/10.3390/agronomy12020378
Van Sinh N, Kato R, Linh DTT, Phuong NTK, Toyota K. Influence of Rice Husk Biochar on Soil Nematode Community under Upland and Flooded Conditions: A Microcosm Experiment. Agronomy. 2022; 12(2):378. https://doi.org/10.3390/agronomy12020378
Chicago/Turabian StyleVan Sinh, Nguyen, Risako Kato, Doan Thi Truc Linh, Nguyen Thi Kim Phuong, and Koki Toyota. 2022. "Influence of Rice Husk Biochar on Soil Nematode Community under Upland and Flooded Conditions: A Microcosm Experiment" Agronomy 12, no. 2: 378. https://doi.org/10.3390/agronomy12020378
APA StyleVan Sinh, N., Kato, R., Linh, D. T. T., Phuong, N. T. K., & Toyota, K. (2022). Influence of Rice Husk Biochar on Soil Nematode Community under Upland and Flooded Conditions: A Microcosm Experiment. Agronomy, 12(2), 378. https://doi.org/10.3390/agronomy12020378