
Influence of the Presence of Grape Skins during White Wine Alcoholic Fermentation





Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Analytical Methodology
2.3. Sensory Analysis
2.4. Statistical Analysis
3. Results and Discussion
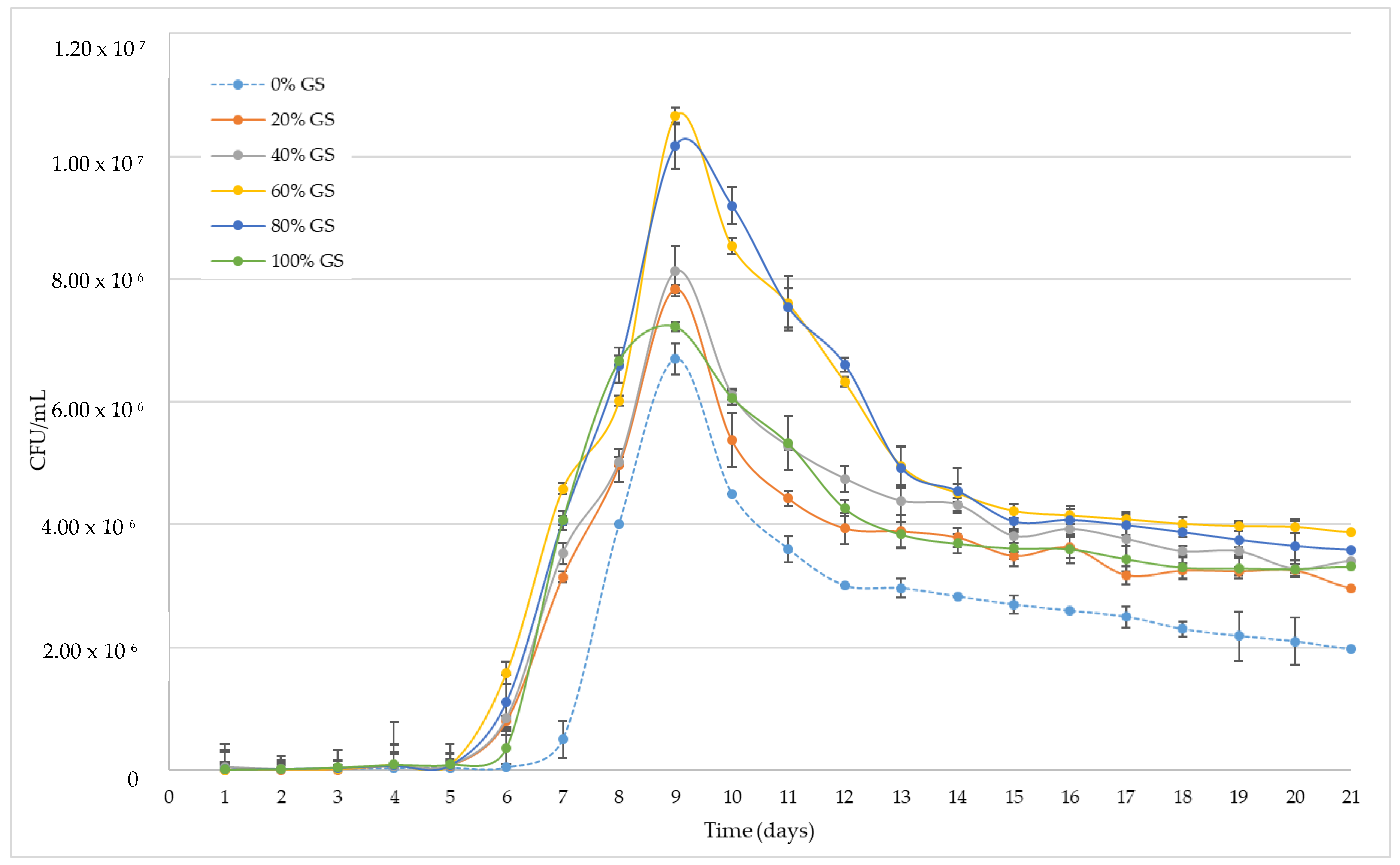
3.1. Grape Skin Effect on Yeast Populations during Alcoholic Fermentation
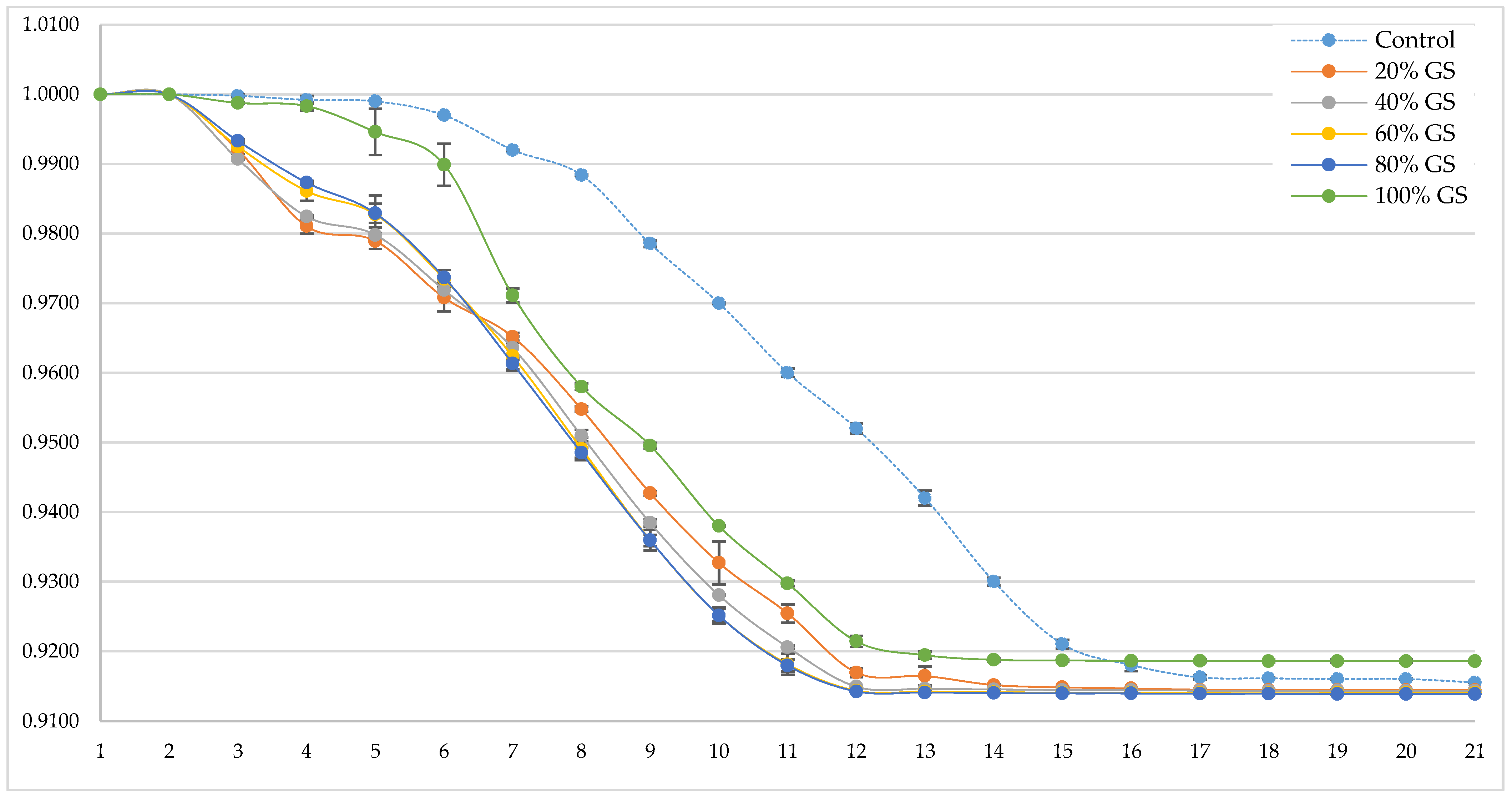
3.2. Grape Skin Effect on Alcoholic Fermentation Kinetics.
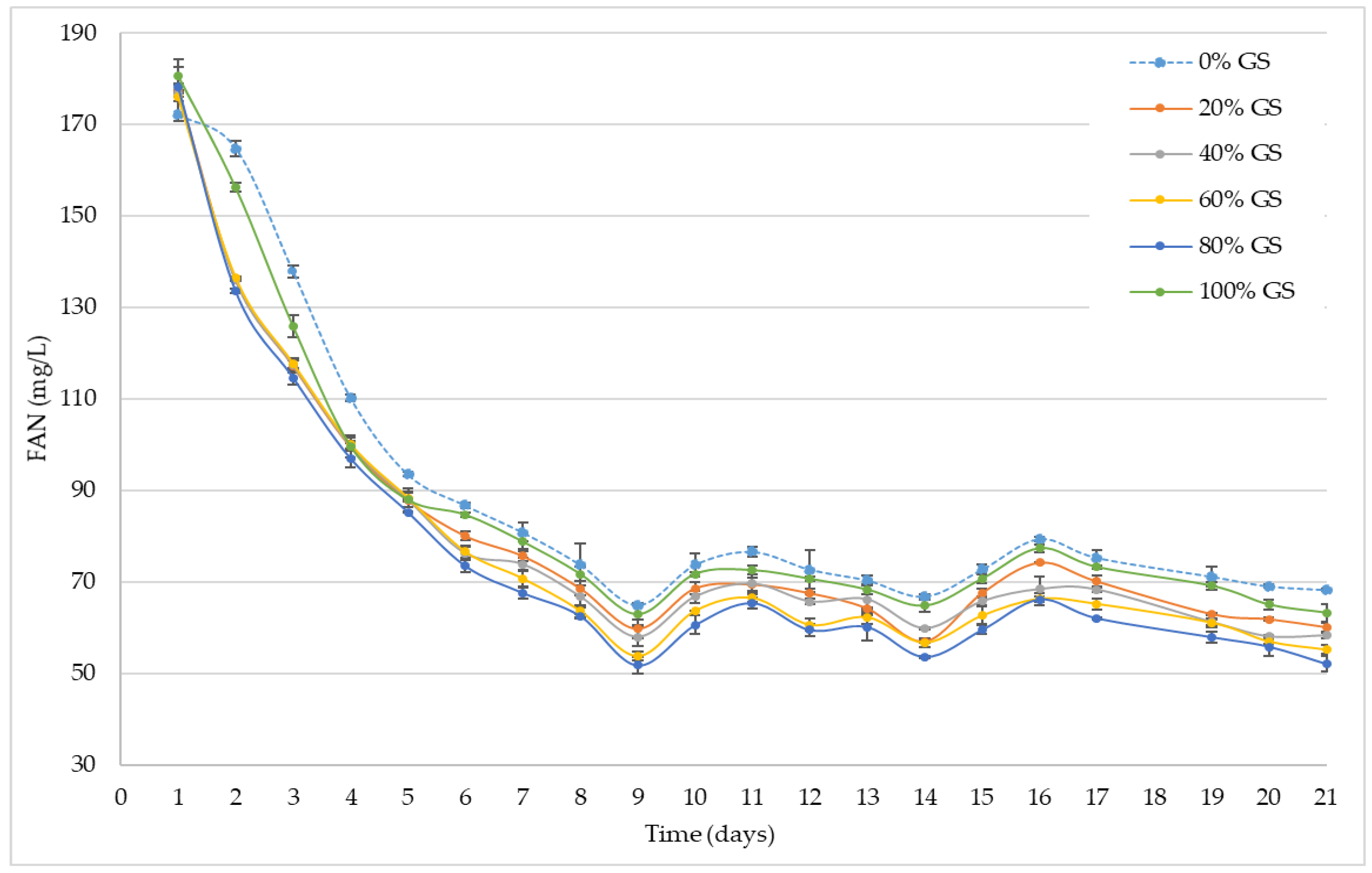
3.3. GS Influence on Free Amino Nitrogen (FAN) during Alcoholic Fermentation
3.4. Final Wines Physicochemical Characterisation
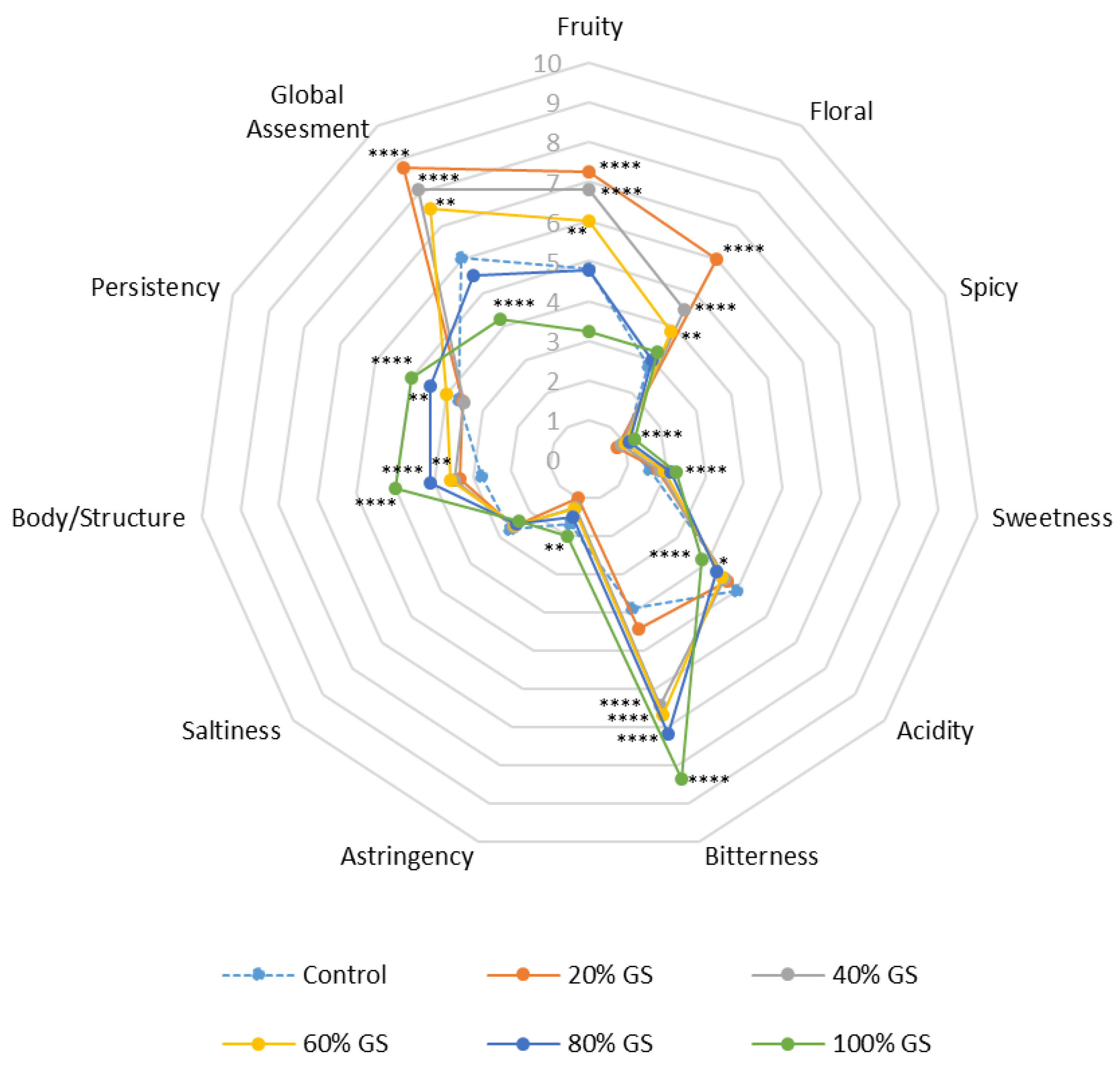
3.5. Sensory Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Marsal, G.; Mateo-Sanz, J.M.; Canals, J.M.; Zamora, F.; Fort, F. SSR analysis of 338 accessions planted in Penedes (Spain) reveals 28 unreported molecular profiles of Vitis vinifera L. Am. J. Enol. Vitic 2016, 67, 466–470. [Google Scholar] [CrossRef]
- International Organization of Vine and Wine (OIV). State of the Vitiviniculture World Market: State of the Sector in 2019. Available online: http://www.oiv.int/public/medias/6679/en-oiv-state-of-the-vitiviniculture-world-market-2019.pdf (accessed on 2 November 2020).
- Conferencia Española de Consejos Reguladores vitivinícolas. El Sector en Cifras. Available online: https://vinosdo.wine/sala-de-prensa/el-sector-en-cifras/ (accessed on 2 November 2020).
- Pszczólkowski, P. Manual de Vinificación: Guía Práctica Para la Elaboración de Vinos, 1st ed.; Ediciones UC: Santiago de Chile, Chile, 2015. [Google Scholar]
- Tomasevic, M.; Lisjak, K.; Vanzo, A.; Basa-Cesnik, H.; Grancin, L.; Curko, N.; Kovacevic-Ganic, K. Change in the compositions of aroma and phenolic compounds induced by different enological practices of Croatian white wine. Polish J. Food Nutr. Sci. 2019, 69, 343–358. [Google Scholar] [CrossRef]
- Del Pozo Bayón, M. Descifrando las Claves Químicas que Explican el Aroma del Vino. Consejo Superior de Investigaciones Científicas (CSIC). Available online: http://www.quimica2011.es/sites/default/files/claves_quimicas_del_aroma_del_vino_0.pdf (accessed on 4 November 2020).
- Schreier, P.; Jennings, W.G. Flavor composition of wines: A review. Crit. Rev. Food Sci. 1979, 12, 59–111. [Google Scholar] [CrossRef]
- Rapp, A.; Mandery, H. Wine aroma. Cell. Mol. Life Sci. 1986, 42, 873–884. [Google Scholar] [CrossRef]
- González-Barreiro, C.; Rial-Otero, R.; Cancho-Grande, B.; Simal-Gándara, J. Wine Aroma Compounds in Grapes: A Critical Review. Crit. Rev. Food Sci. 2014, 55, 202–218. [Google Scholar] [CrossRef] [PubMed]
- Darias-Martin, J.J.; Rodríguez, O.; Díaz, E.; Lamuela-Raventos, R.M. Effect of skin contact on the antioxidant phenolic in white wine. Food Chem. 2000, 7, 483–487. [Google Scholar] [CrossRef]
- Aleixandre-Tudo, J.L.; Weightman, C.; Panzeri, V.; Nieuwoudt, H.H.; du Toit, W.J. Effect of skin contact before and during alcoholic fermentation on the chemical and sensory profile of South African Chenin blanc white wines. S. Afr. J. Enol. Vitic. 2015, 36, 366–377. [Google Scholar] [CrossRef]
- Selli, S.; Canbas, A.; Cabaroglu, T.; Erten, H.; Gunata, Z. Aroma components of cv. Muscat of Bornova wines and influence of skin contact treatment. Food Chem. 2006, 94, 319–326. [Google Scholar] [CrossRef]
- Cheynier, V.; Rigaud, J.; Souquet, J.M.; Barillere, J.M.; Moutonet, M. Effect of pomace contact and hiperoxidation on the phenolic composition and quality of Grenache and Chardonnay wines. Am. J. Enol. Vitic. 1989, 40, 36–42. [Google Scholar]
- Palomo, E.S.; Pérez-Coello, M.S.; Díaz-Maroto, M.C.; González-Viñas, M.A.; Cabezudo, M.D. Contribution of free and glycosicadilly-bound volatile compounds to the aroma of muscat “a petit grains” wines and effect of skin contact. Food Chem. 2006, 95, 279–289. [Google Scholar] [CrossRef]
- Singleton, V.L.; Zaya, J.; Trousdale, E. White table wine quality and polyphenol composition as affected by must SO2 content and pomace contact time. Am. J. Enol. Vitic. 1980, 31, 14–20. [Google Scholar]
- Sánchez-Palomo, E.; González-Viñas, M.A.; Díaz-Maroto, M.C. Aroma potential of Albillo wines and effect of skin-contact treatment. Food Chem. 2007, 103, 631–640. [Google Scholar] [CrossRef]
- Barisashvili, G. Making Wine in Qvevri: A Unique Georgian Tradition. Available online: https://www.qvevriproject.org/Files/2011.00.00_Elkana_Barisashvili_MakingWineInQvevri_eng.pdf (accessed on 4 November 2020).
- Bene, Z.; Kállay, M. Polyphenol contents of skin-contact fermented white wines. Acta Aliment. 2019, 48, 515–524. [Google Scholar] [CrossRef]
- Lorteau, S. A comparative legal analysis of skin-contact wine definitions in Ontario and South Africa. J. Wine Res. 2018, 29, 265–277. [Google Scholar] [CrossRef]
- López, I.; Morales, J.; Ramirez, P.; (Instituto de Formación Agraria y Pesquera, IFAPA. Consejería de Agricultura y Pesca, Junta de Andalucía); Palencia, L.; Romero, C.; (Sociedad Cooperativa AECOVI, Jerez de la Frontera). Personal communication, 2007.
- Palacios, V.; Roldán, A.; Jiménez-Cantizano, A.; Amores-Arrocha, A. Physicochemical and microbiological characterization of the sensory deviation responsible for the origin of the special sherry wines “palo cortado” type. PLoS ONE 2018, 13, e0208330. [Google Scholar] [CrossRef] [Green Version]
- Cejudo-Bastante, M.J.; Castro-Vázquez, L.; Hermosín-Gutiérrez, I.; Pérez-Coello, M.S. Combined Effects of Prefermentative Skin Maceration and Oxygen Addition of Must on Color-Related Phenolics, Volatile Composition, and Sensory Characteristics of Airén White Wine. J. Agric. Food Chem. 2011, 59, 12171–12182. [Google Scholar] [CrossRef] [PubMed]
- Olejar, K.J.; Fedrizzi, B.; Kilmartin, P.A. Enhancement of chardonnay antioxidant activity and sensory perception through maceration technique. LWT 2016, 65, 152–157. [Google Scholar] [CrossRef]
- Albisu-Aguado, L.M.; Zeballos, M.G. Consumo de vino en España. Tendencias y comportamiento del consumidor. In La Economía del Vino en España y en el Mundo, 1st ed.; Compés-López, R., Castillo-Valero, J.S., Eds.; Monografías Cajamar: Murcia, España, 2014; pp. 99–140. [Google Scholar]
- Casassa, F.; Beaver, C.W.; Mireles, M.S.; Harbertson, J.F. Effect of extended maceration and ethanol concentration on the extraction and evolution of phenolics, colour components and sensory attributes of Merlot wines. Aust. J. Grape Wine Res. 2013, 19, 25–39. [Google Scholar] [CrossRef]
- OIV Office International de la Vigne et du Vin. Recuéil des Methods Internationals D’analyse des Vins et des Moûts; Edition Oficielle: Paris, France, 2014. [Google Scholar]
- Sancho-Galán, P.; Amores-Arrocha, A.; Jiménez-Cantizano, A.; Palacios, V. Use of Multiflora Bee Pollen as a Flor Velum Yeast Growth Activator in Biological Aging Wines. Molecules 2019, 24, 1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amores-Arrocha, A.; Sancho-Galán, P.; Jiménez-Cantizano, A.; Palacios, V. Bee Pollen as Oenological Tool to Carry out Red Winemaking in Warm Climate Conditions. Agronomy 2020, 10, 634. [Google Scholar] [CrossRef]
- Abernathy, D.G.; Spedding, G.; Starcher, B. Analysis of Protein and Total Usable Nitrogen in Beer and Wine Using a Microwell Ninhydrin Assay. J. Inst. Brew 2009, 115, 122–127. [Google Scholar] [CrossRef]
- Gonçalves, C.; Rodriguez-Jasso, R.M.; Gomes, N.; Teixeira, J.A.; Belo, I. Adaptation of dinitrosalycilic acid method to microtiter plates. Anal. Methods 2010, 2, 2046–2048. [Google Scholar] [CrossRef] [Green Version]
- Almela, L.; Javaloy, S.; Fernández-López, J.A.; López-Roca, J.M. Varietal classification of young red wines in terms of chemical and colour parameters. J. Sci. Food Agric. 1996, 70, 173–180. [Google Scholar] [CrossRef]
- Heredia, F.J.; Troncoso, A.M.; Guzmán-Chozas, M. Multivariate characterization of aging status in red wines based on chromatic parameters. Food Chem. 1997, 60, 103–108. [Google Scholar] [CrossRef]
- Pérez-Magariño, S.; González-San José, M.L. Polyphenols and colour variability of red wines made from grapes harvested at different ripeness grade. Food Chem. 2006, 96, 197–208. [Google Scholar] [CrossRef]
- ISO 3591. Sensory Analysis -Apparatus- Wines- Tasting Glass; International Organization for Standardization: Genève, Switzerland, 1997. [Google Scholar]
- Jackson, R.S. Wine Tasting: A Professional Handbook; Academic Press: London, UK, 2009. [Google Scholar]
- Carbonell-Bejerano, P.; Martínez-Zapater, J.M. Estructura y Composición de la Uva y Su Contribución al Vino. SEBBM, Bioquímica del Vino. Available online: https://www.sebbm.es/revista/articulo.php?id=212&url=estructura-y-composicion-de-la-uva-y-su-contribucion-al-vino (accessed on 9 November 2020).
- Flamini, R.; De Rosso, M. Mass spectrometry in the analysis of grape and wine proteins. Expert Rev. Proteomic 2006, 3, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Bardi, L.; Cocito, C.; Marzona, M. Saccharomyces cerevisiae cell fatty acid composition and reléase during fermentation without aireation and in absence of exogenous lipids. Int. J. Food Microbiol. 1999, 47, 133–140. [Google Scholar] [CrossRef]
- Lambrechts, M.; Pretorius, I.S. Yeast and its importance to wine aroma. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Viegas, C.A.; Rosa, M.F.; Sa-Correia, I.; Novais, J.M. Inhibition of yeast growth by actanoic and decanoic acids produced during ethanolic fermentation. Appl. Environ. Microbiol. 1989, 55, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Torija-Martínez, M.J. Ecología de Levaduras: Selección y Adaptación a Fermentaciones Vínicas. Ph.D. Thesis, Universitat Rovira i Virgili, Tarragona, Spain, 2002. [Google Scholar]
- Nguela, J.M.; Vernhet, A.; Julien-Ortiz, A.; Sieczkowski, N.; Mouret, J.-R. Effect of grape must polyphenols on yeast metabolism during alcoholic fermentation. Food Res. Int. 2019, 121, 161–175. [Google Scholar] [CrossRef]
- Li, J.Y.; Du, G.; Yang, X.; Huang, W.D. Effect of proanthocyanidins on yeast metabolism, H + -ATPase activity, and wine fermentation. Am. J. Enol. Vitic. 2011, 62, 512–518. [Google Scholar] [CrossRef]
- Henschke, P.A.; Jiranek, V. Yeasts—Metabolism of nitrogen compounds. In Wine Microbiology and Biotechnology, 1st ed.; Fleet, G.H., Ed.; CRC Press: Boca Raton, FL, USA, 1993; pp. 77–164. [Google Scholar]
- Barre, P.; Blondin, P.; Dequin, S.; Feuillat, M.; Sablayrolles, J.M.; Salmon, J.M. La levure de Fermentation Alcoolique; Oenologie Technique et Documentation; Lovisier: Paris, France, 1998; pp. 414–495. [Google Scholar]
- Dizy, M.; Polo, M.C. Changes in concentration of nitrogenous compounds during fermentation of white grape musts at pilot plant scale. Food Sci. Technol. Int. 1996, 2, 87–93. [Google Scholar] [CrossRef]
- Fornairon-Bonneford, C.; Camarasa, C.; Moutounet, M.; Salmon, J.M. New Trends on yeast autolysis and wine ageing on lees: A bibliographical review. Int. J. Vine Wine Sci. 2001, 36, 49–69. [Google Scholar]
- Moreno-Arribas, M.V.; Polo, M.C. Wine Chemistry and Biochemistry; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Pérez-Serradilla, J.A.; Luque de Castro, M.D. Role of lees in wine production: A review. Food Chem. 2008, 111, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Ough, C.; Davenport, M.; Joseph, K. Effects of Certain Vitamins on Growth and Fermentation Rate of Several Commercial Active Dry Yeasts. Am. J. Enol. Vitic. 1989, 40, 208–213. [Google Scholar]
- Olejar, K.J.; Fedrizzi, B.; Kilmartin, P.A. Antioxidant activity and phenolic profiles of Sauvignon Blanc wines made by various maceration techniques. Aust. J. Grape Wine Res. 2015, 21, 57–68. [Google Scholar] [CrossRef]
- Ough, C.S. Substances extracted during skin contact with white musts. I. General wine composition and quality changes with contact time. Am. J. Enol. Vitic. 1969, 37, 93–100. [Google Scholar]
- Test, S.L.; Noble, A.C.; Schmidt, J.O. Effect of pomace contact on chardonnay musts and wines. Am. J Enol. Vitic. 1986, 37, 133–136. [Google Scholar]
- Gomez, E.; Laencina, J.; Martínez, A. Vinification effects on changes in colatile compounds of wine. J. Food Sci. 1994, 59, 406–409. [Google Scholar] [CrossRef]
- Liu, S.Q. Malolactic fermentation in wine—Beyond deaidification. J. Appl. Microbiol. 2002, 92, 589–601. [Google Scholar] [CrossRef]
- Guzzon, R.; Malacarne, M.; Larcher, R.; Franciosi, E.; Toffanin, A. The impact of grape processing and carbonic maceration on the microbiota of early stages of winemaking. J. Appl. Microbiol. 2019, 128, 209–224. [Google Scholar] [CrossRef] [Green Version]
- Amores-Arrocha, A.; Roldán, A.; Jiménez-Cantizano, A.; Caro, I.; Palacios, V. Effect on White Grape Musts of Multiflora Bee Pollen Addition during the Alcoholic Fermentation Process. Molecules 2018, 23, 1321. [Google Scholar] [CrossRef] [Green Version]
- Soares, S.; Brandão, E.; Guerreiro, C.; Soares, S.; Mateus, N.; de Freitas, V. Tannins in Food: Insights into the Molecular Perception of Astringency and Bitter Taste. Molecules 2020, 25, 2590. [Google Scholar] [CrossRef]
- Soares, S.; Brandão, E.; Mateus, N.; de Freitas, V. Sensorial properties of red wine polyphenols: Astringency and bitternes. Crit. Rev. Food Sci. Nutr. 2017, 57, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Di Lecce, G.; Arranz, S.; Jáuregui, O.; Tresserra-Rimbau, A.; Quifer-Rada, P.; Lamuela-Raventos, R.M. Phenolic profiling of the skin, pulp and sedes of Albariño grapes using hybrid quadrupole time-of-flight and triple quadrupole mass spectrometry. Food Chem. 2014, 145, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Gawel, R.; Day, M.; Van Sluyter, S.C.; Holt, H.; Waters, E.J.; Smith, P.A. White Wine Taste and Mouthfeel as Affected by Juice Extraction and Processing. J. Agic. Food Chem. 2014, 62, 10008–10014. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | 20% GS | 40% GS | 60% GS | 80% GS | 100% GS | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TA (g/L) | 4.52 | ± | 0.02 a | 4.12 | ± | 0.0 b | 4.05 | ± | 0.02 b | 3.91 | ± | 0.02 b | 3.85 | ± | 0.03 b | 3.78 | ± | 0.02 b |
| VA (g/L) | 0.15 | ± | 0.01 a | 0.24 | ± | 0.02 b | 0.33 | ± | 0.03 c | 0.42 | ± | 0.01 d | 0.62 | ± | 0.05 e | 0.72 | ± | 0.04 f |
| % Alc. | 11.73 | ± | 0.11 a | 11.92 | ± | 0.12 a | 11.85 | ± | 0.12 a | 12.05 | ± | 0.17 a | 11.98 | ± | 0.10 a | 11.81 | ± | 0.02 a |
| RS (g/L) | 2.32 | ± | 0.14 a | 1.87 | ± | 0.03 b | 1.72 | ± | 0.02 c | 1.68 | ± | 0.04 c | 1.70 | ± | 0.02 c | 2.02 | ± | 0.06 d |
| Malic Acid (mg/L) | 329.90 | ± | 0.80 a | 440.52 | ± | 0.20 b | 463.18 | ± | 1.00 c | 472.32 | ± | 1.20 d | 480.70 | ± | 0.20 e | 487.20 | ± | 0.20 f |
| Lactic Acid (mg/L) | 104.86 | ± | 1.00 a | 70.02 | ± | 1.60 b | 92.94 | ± | 2.20 c | 136.30 | ± | 0.20 d | 136.68 | ± | 0.14 d | 161.08 | ± | 0.20 e |
| L* | 96.48 | ± | 0.34 a | 98.20 | ± | 0.23 a | 98.38 | ± | 0.3 a | 98.79 | ± | 0.25 a | 98.49 | ± | 0.48 a | 98.30 | ± | 0.15 a |
| a* | −0.50 | ± | 0.07 a | −0.57 | ± | 0.02 b | −0.60 | ± | 0.02 c | −0.63 | ± | 0.03 b | −0.58 | ± | 0.02 b | −0.60 | ± | 0.06 b |
| b* | 14.16 | ± | 0.67 a | 7.39 | ± | 0.47 b | 6.84 | ± | 0.63 b | 6.07 | ± | 0.00 b | 5.99 | ± | 0.00 b | 6.82 | ± | 0.32 b |
| H* | 92.05 | ± | 0.38 a | 94.4 | ± | 0.47 a | 95.02 | ± | 0.57 a | 95.98 | ± | 0.31 a | 95.63 | ± | 0.13 a | 94.80 | ± | 0.31 a |
| C* | 14.17 | ± | 0.67 a | 7.41 | ± | 0.47 b | 6.87 | ± | 0.63 b | 6.15 | ± | 0.24 b | 6.02 | ± | 0.01 b | 6.85 | ± | 0.32 b |
| Abs 420 nm | 0.20 | ± | 0.01 a | 0.11 | ± | 0.01 b | 0.10 | ± | 0.01 b | 0.08 | ± | 0.01 c | 0.09 | ± | 0.01 b,c | 0.10 | ± | 0.01 c |
| TPI | 0.470 a | ± | 0.068 | 0.544 | ± | 0.024 a | 0.696 | ± | 0.090 b | 0.864 | ± | 0.081 c | 0.887 | ± | 0.039 c | 0.901 | ± | 0.075 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sancho-Galán, P.; Amores-Arrocha, A.; Jiménez-Cantizano, A.; Palacios, V. Influence of the Presence of Grape Skins during White Wine Alcoholic Fermentation. Agronomy 2021, 11, 452. https://doi.org/10.3390/agronomy11030452
Sancho-Galán P, Amores-Arrocha A, Jiménez-Cantizano A, Palacios V. Influence of the Presence of Grape Skins during White Wine Alcoholic Fermentation. Agronomy. 2021; 11(3):452. https://doi.org/10.3390/agronomy11030452
Chicago/Turabian StyleSancho-Galán, Pau, Antonio Amores-Arrocha, Ana Jiménez-Cantizano, and Víctor Palacios. 2021. "Influence of the Presence of Grape Skins during White Wine Alcoholic Fermentation" Agronomy 11, no. 3: 452. https://doi.org/10.3390/agronomy11030452
APA StyleSancho-Galán, P., Amores-Arrocha, A., Jiménez-Cantizano, A., & Palacios, V. (2021). Influence of the Presence of Grape Skins during White Wine Alcoholic Fermentation. Agronomy, 11(3), 452. https://doi.org/10.3390/agronomy11030452