
Do Crop Rotations in Rice Reduce Weed and Echinochloa spp. Infestations? Recommendations for Integrated Weed Control


Abstract
:1. Introduction
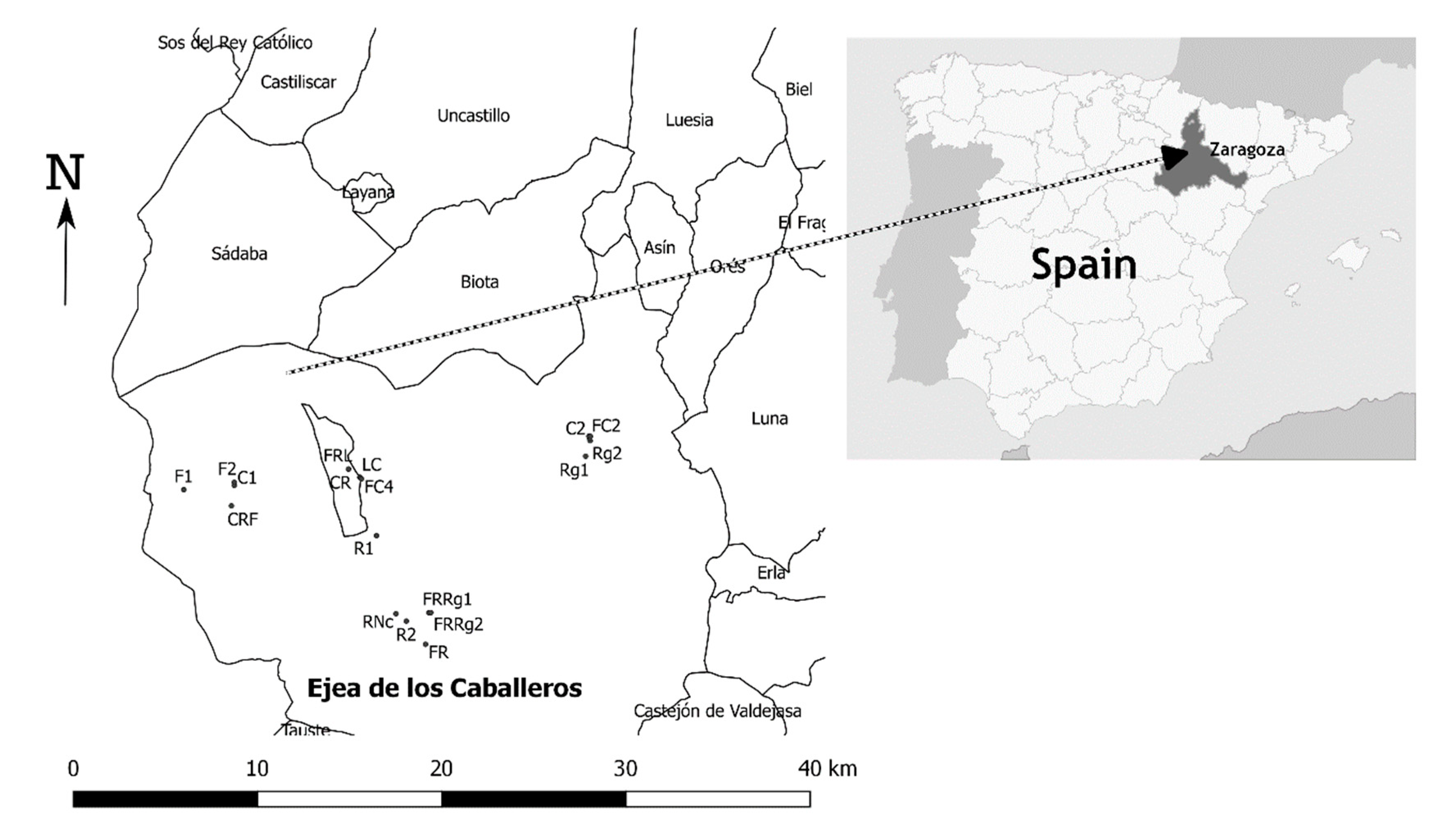
2. Materials and Methods
2.1. Weed Survey
2.2. Soil Analysis
2.3. Data Analysis
3. Results
3.1. General Results
3.2. Soil Samples
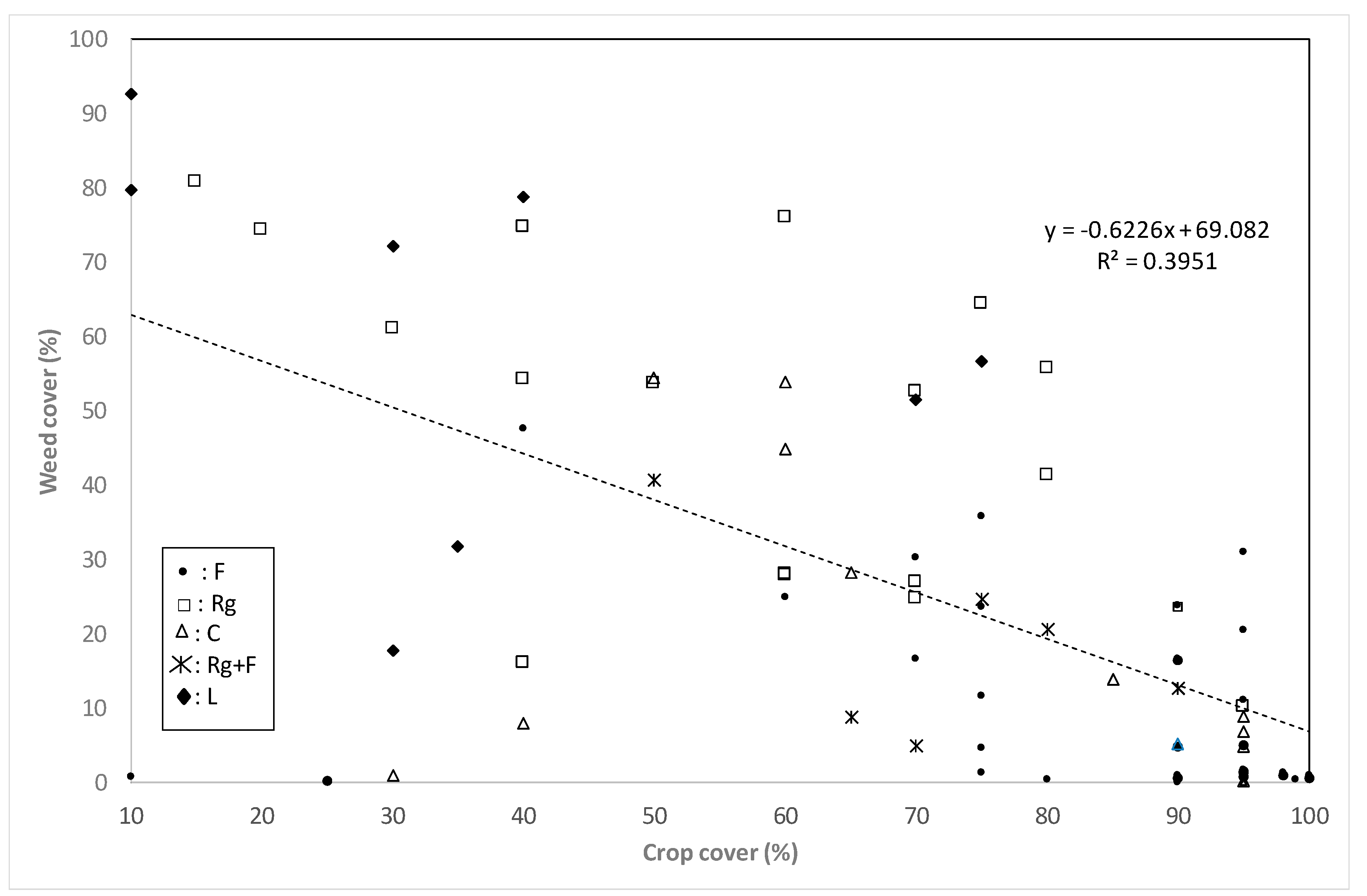
3.3. Crop Cover
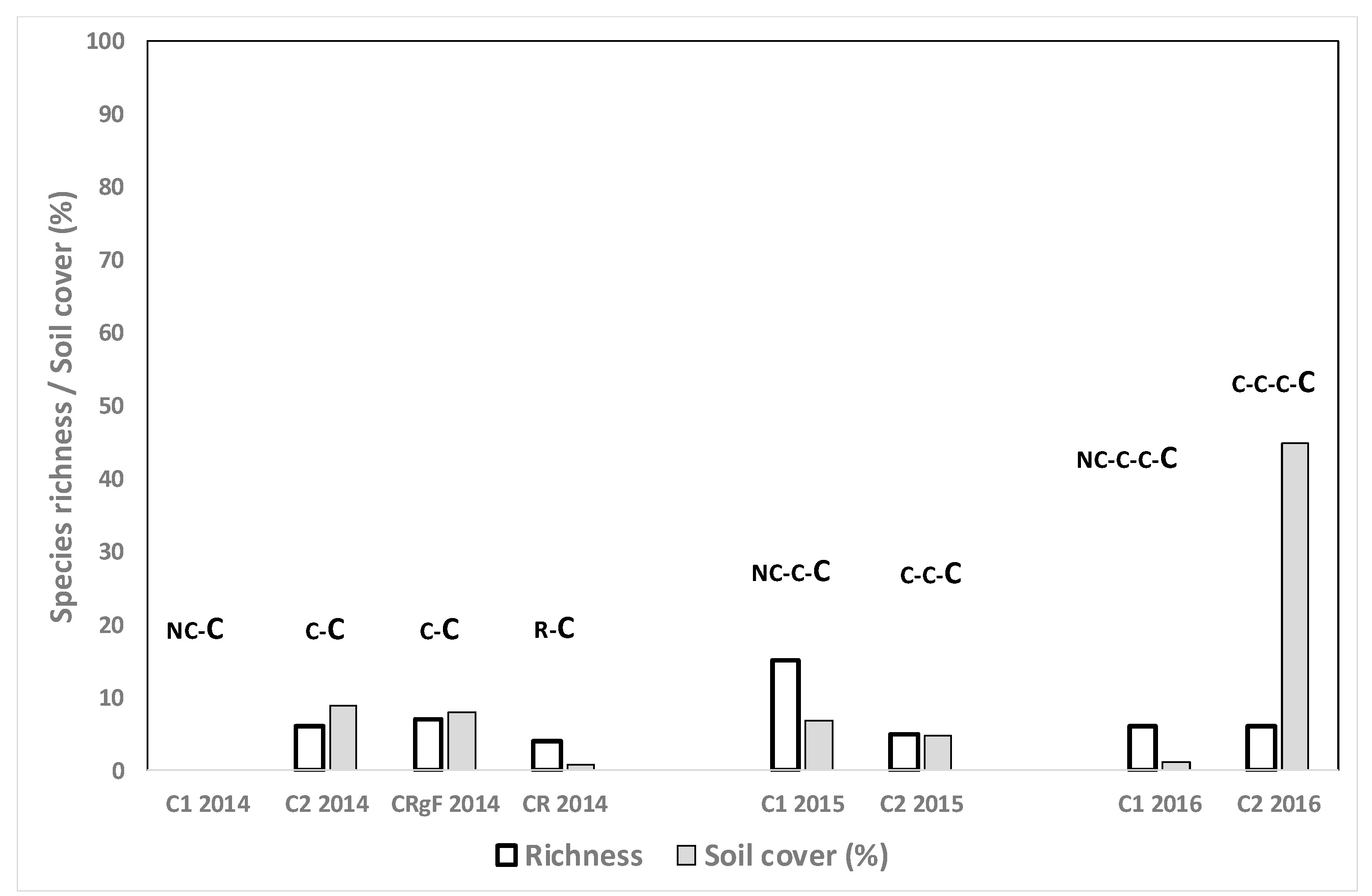
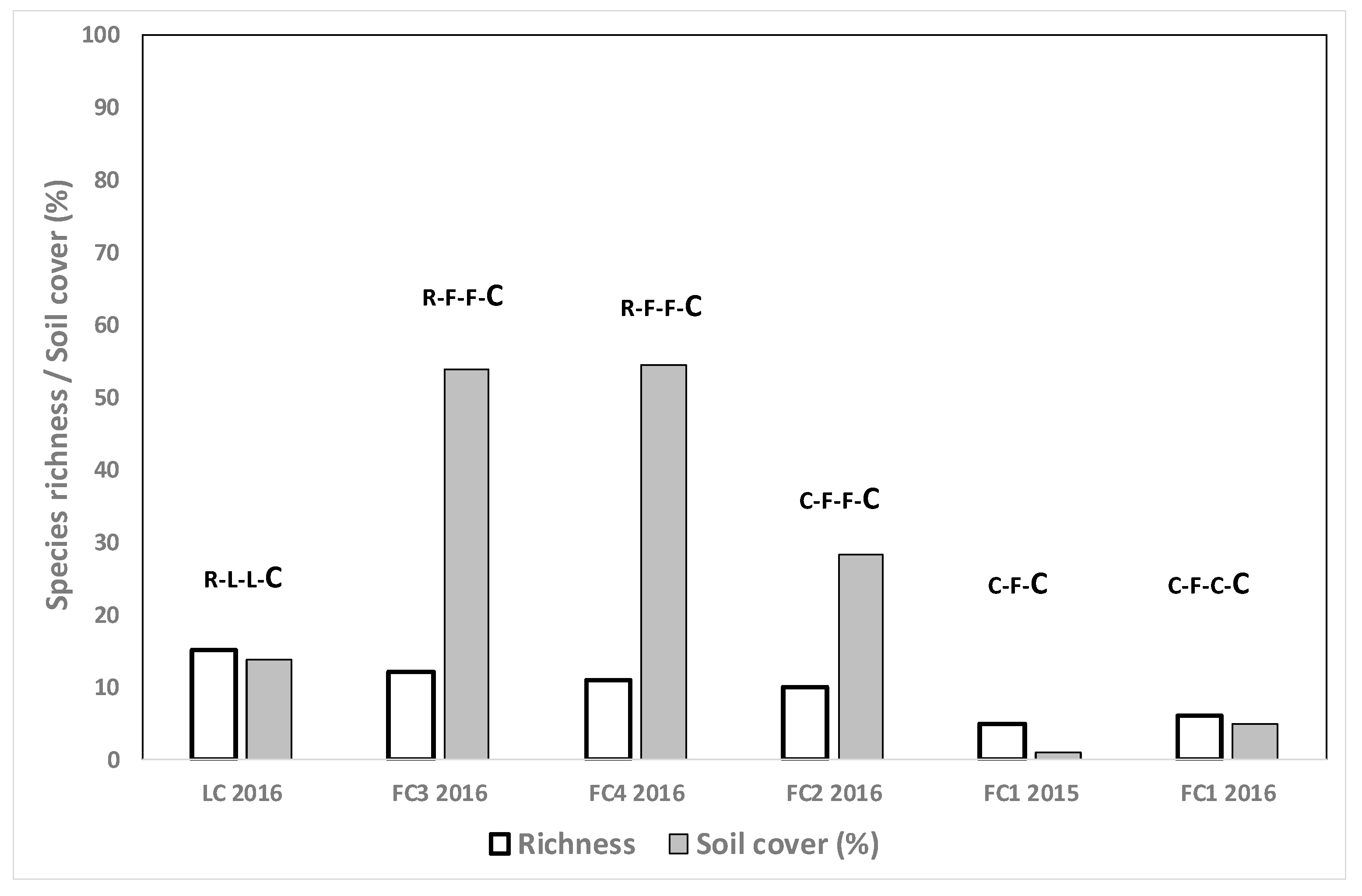
3.4. Species Richness and Weed Cover in Different Crop Situations
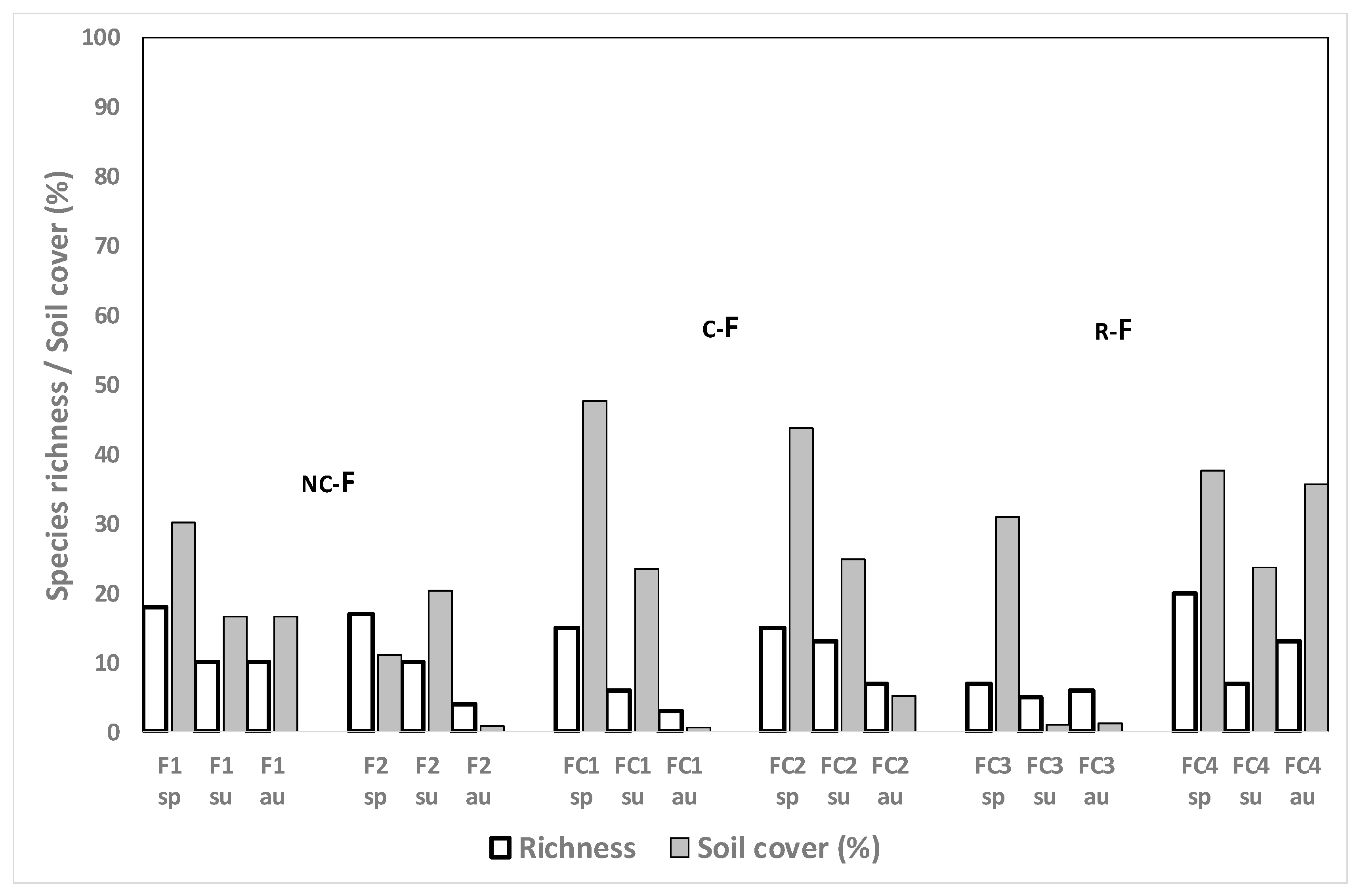
3.4.1. Winter Cereal Fields, Spring Evaluation
3.4.2. Winter Cereal Fields, Summer and Autumn Evaluations
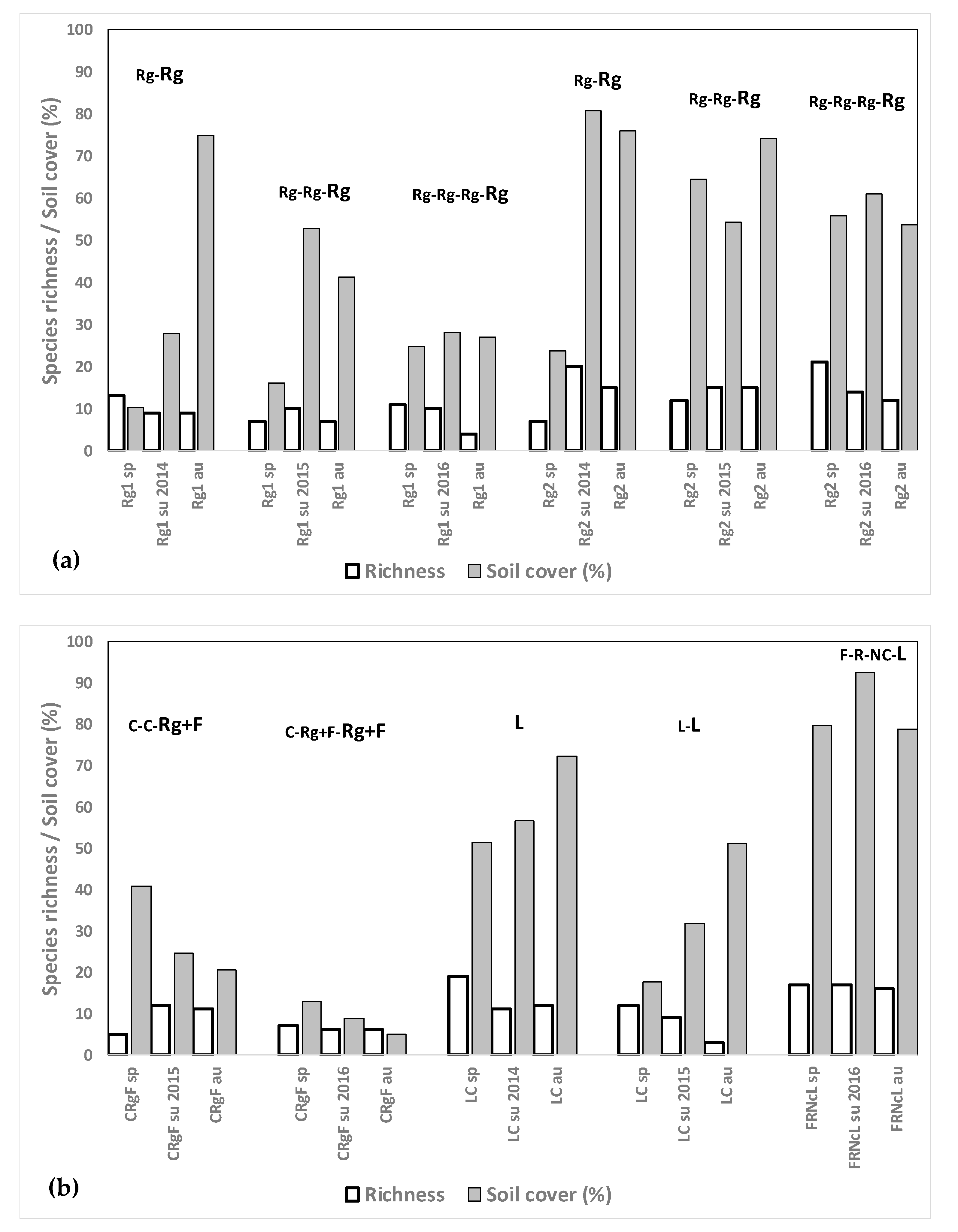
3.4.3. Fescue Fields after Rice
3.4.4. Forage Crops in Other Positions of the Rotation
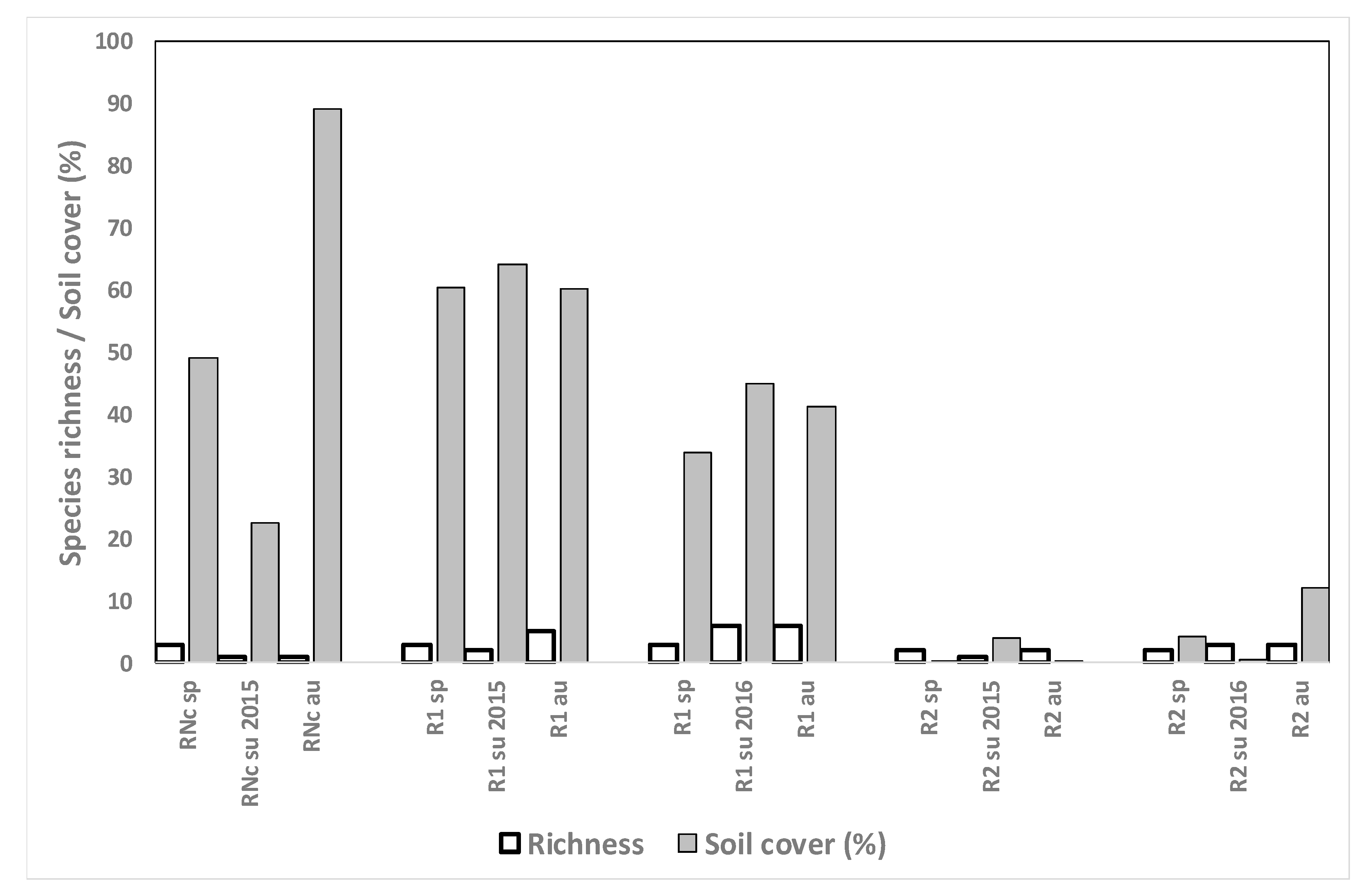
3.4.5. Monocrop Rice Fields
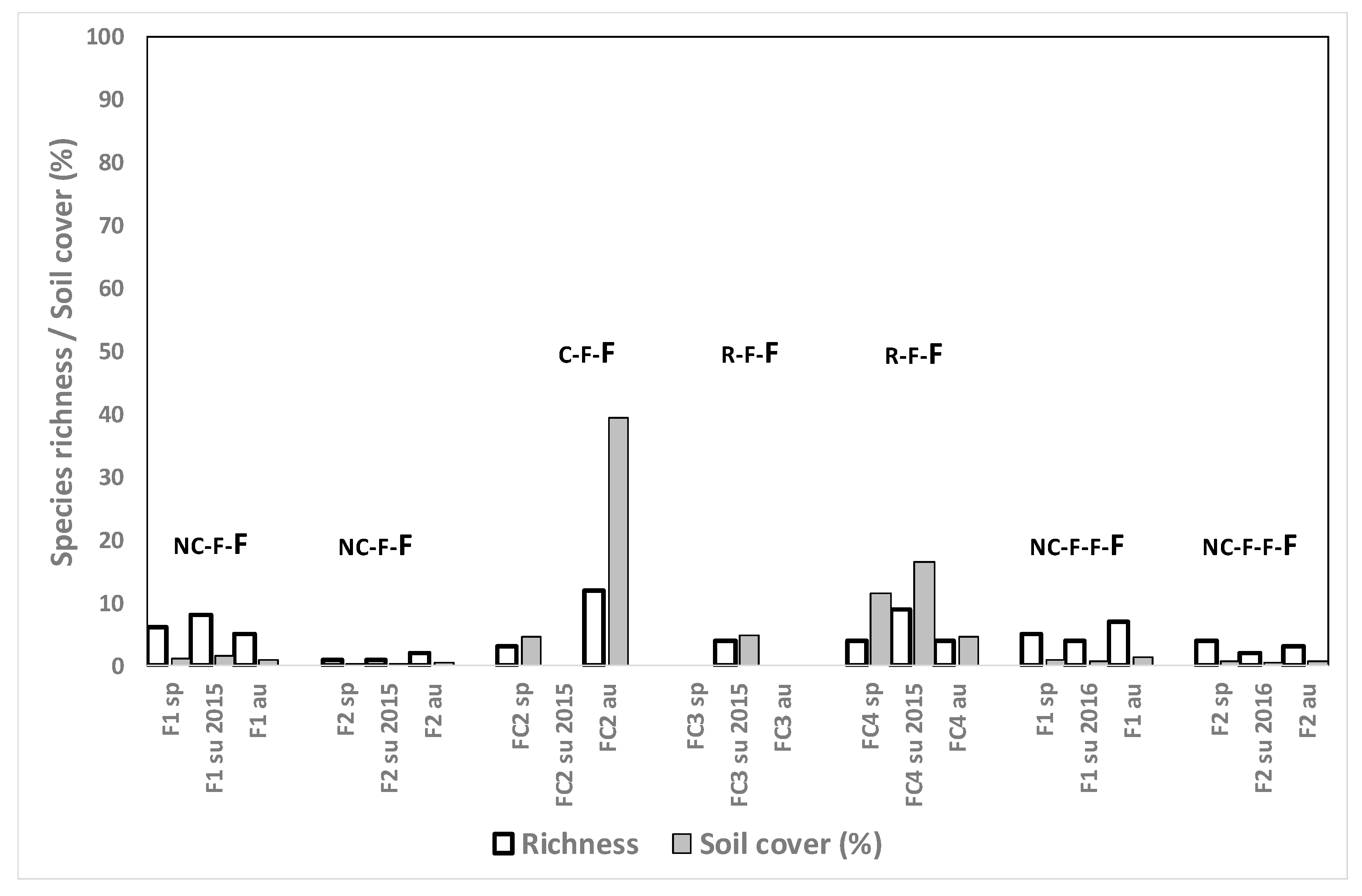
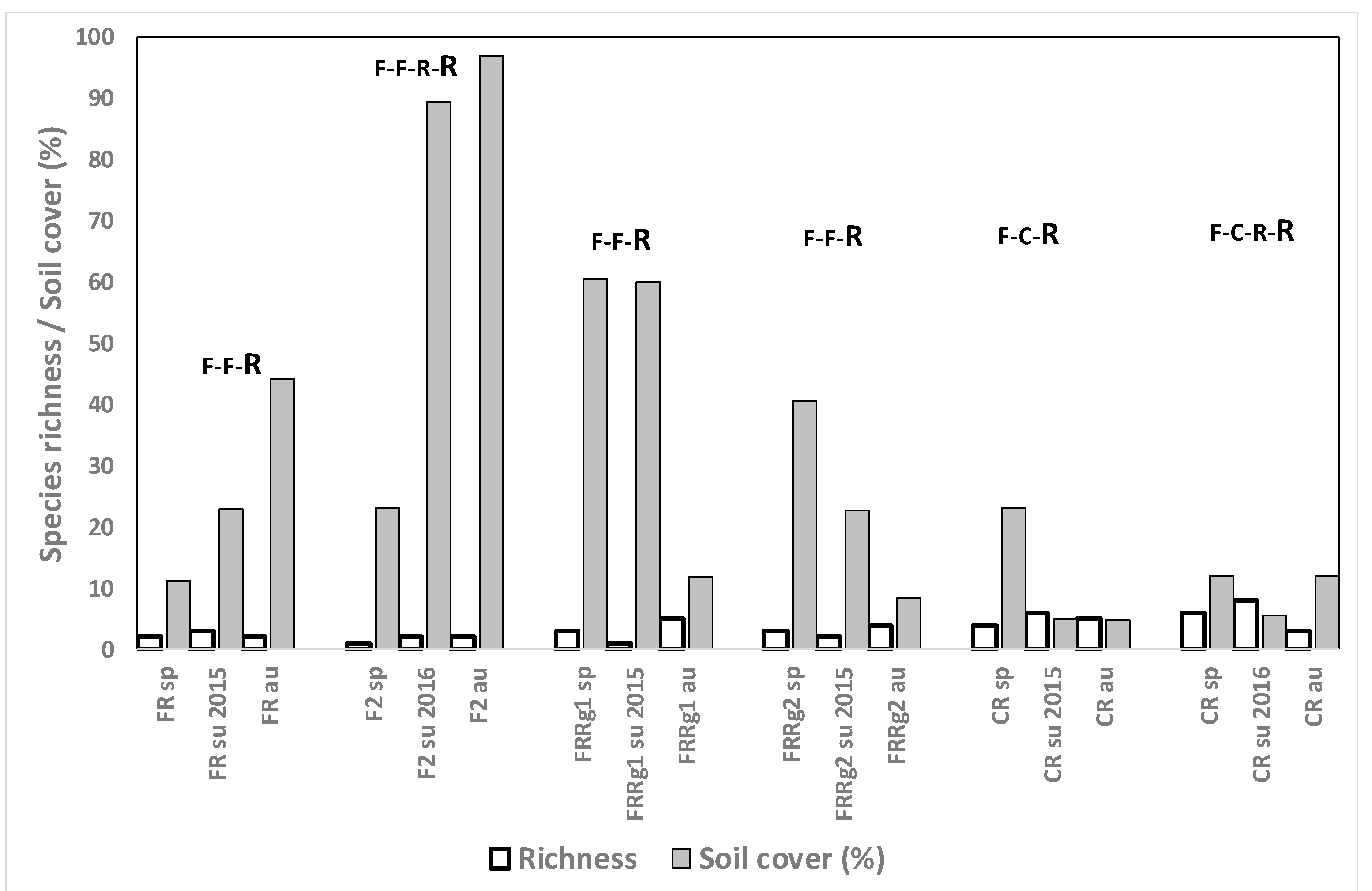
3.4.6. Rice after Growing Other Crops
4. Discussion
4.1. General Results
4.2. Crop and Weed Cover
4.3. Winter Cereal Fields
4.4. Fescue, Ryegrass and Lucerne Fields
4.5. Monocrop Rice Fields
4.6. Rice after Growing Other Crops
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kraehmer, H.; Jabran, K.; Mennan, H.; Chauhan, B.S. Global distribution of rice weeds—A review. Crop Prot. 2016, 80, 73–86. [Google Scholar] [CrossRef]
- Kraehmer, H.; Thomas, C.; Vidotto, F. Rice production in Europe. In Rice Production Worldwide; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Bagavathiannan, M.V.; Norsworthy, J.K.; Smith, K.L.; Burgos, N. Seedbank size and emergence pattern of barnyardgrass (Echinochloa crus-galli) in Arkansas. Weed Sci. 2011, 59, 359–365. [Google Scholar] [CrossRef]
- Ruiz-Santaella, J.P.; Fisher, A.J.; De Prado, R. Alternative control of two biotypes of Echinochloa phyllopogon susceptible and resistant to fenoxaprop-ethyl. Commun. Agric. Appl. Biol. Sci. 2003, 68, 403–407. [Google Scholar]
- Lin, D.; Tsuzuki, E.; Dong, Y.; Terao, H.; Xuan, T.D. Potential biological control of weeds in rice fields by allelopathy of dwarf lilyturf plants. BioControl 2004, 49, 187–196. [Google Scholar] [CrossRef]
- Wang, X.L.; Zhang, Z.Y.; Xu, X.M.; Li, G. The density of barnyard grass affects photosynthesis and physiological characteristics of rice. Photosynthetica 2019, 57, 705–711. [Google Scholar] [CrossRef] [Green Version]
- Heap, I. International Survey of Herbicide Resistant Weeds. Available online: http://www.weedscience.org/Summary/Species.aspx (accessed on 30 May 2019).
- Giannopolitis, C.N.; Vassiliou, G. Propanil tolerance in Echinochloa crus-galli (l.) Beauv. Trop. Pest Manag. 1989, 35, 6–7. [Google Scholar] [CrossRef]
- Romano, Y.; Mendoza, F.; Palmerín, J.A.; Quiles, J.; Amaro, I.; Osuna, M.D. Distribución de poblaciones resistentes de Echinochloa spp. a herbicidas inhibidores de la ALS y ACCasa en Extremadura. In Proceedings of the XVI Congreso de la Sociedad Española de Malherbología, Pamplona, Spain, 25–27 October 2017; Royuela, M., Zabalza, A., Eds.; Universidad Pública de Navarra: Navarra, Spain, 2017; pp. 269–271. [Google Scholar]
- Pardo, G.; Marí, A.; Fernández-Cavada, S.; García-Floria, C.; Hernández, S.; Zaragoza, C.; Cirujeda, A. Alternatives to penoxsulam to control Echinochloa spp. and cyperaceous weeds in rice crop in NE Spain. ITEA Inf. Tec. Econ. Agrar. 2015, 111, 295–309. [Google Scholar] [CrossRef]
- Farrell, T.C.; Fox, K.M.; Williams, R.L.; Fukai, S.; Lewin, L.G. Minimising cold damage during reproductive development among temperate rice genotypes. II. Genotypic variation and flowering traits related to cold tolerance screening. Aust. J. Agric. Res. 2006, 57, 98–100. [Google Scholar] [CrossRef]
- Anderson, R.L. Managing weeds with a dualistic approach of prevention and control. A review. Agron. Sustain. Dev. 2007, 27, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Adeux, G.; Munier-Jolain, N.; Meunier, D.; Farcy, P.; Carlesi, S.; Barberi, P.; Cordeau, S. Diversified grain-based cropping systems provide long-term weed control while limiting herbicide use and yield losses. Agron. Sustain. Dev. 2019, 39, 42. [Google Scholar] [CrossRef]
- Burnside, O.C.; Wilson, R.G.; Weisberg, S.; Hubbard, K.G. Seed longevity of 41 weed species buried 17 years in eastern and western Nebraska. Weed Sci. 1996, 44, 74–86. [Google Scholar] [CrossRef]
- Gardarin, A.; Dürr, C.; Mannino, M.R.; Busset, H.; Colbach, N. Seed mortality in the soil is related to seed coat thickness. Seed Sci. Res. 2010, 20, 243–256. [Google Scholar] [CrossRef]
- Liebman, M.; Gallandt, E.R. Many Little Hammers: Ecological Management of Crop-Weed Interactions. In Ecology in Agriculture; Liebman, E.M., Gallandt, E.R., Eds.; Academic Press: Cambridge, MA, USA, 1997; pp. 291–343. [Google Scholar] [CrossRef]
- Bosnic, A.C.; Swanton, C.J. Influence of barnyardgrass (Echinochloa crus-galli) time of emergence and density on corn (Zea mays). Weed Sci. 1997, 45, 276–282. [Google Scholar] [CrossRef]
- Awan, T.H.; Chauhan, B.S. Effect of emergence time, inter- and intra-specific competition on growth and fecundity of Echinochloa crus-galli in dry-seeded rice. Crop Prot. 2016, 87, 98–107. [Google Scholar] [CrossRef]
- Awan, T.H.; Saleem, M.U.; Hussain, S.; Ahmed, S.; Ali, H.H. Integrated weeds management in dry-seeded basmati rice. J. Anim. Plant Sci. 2019, 29, 1299–1307. [Google Scholar]
- Wei, H.; Bai, W.; Zhang, J.; Chen, R.; Xiang, H.; Quan, G. Integrated rice-duck farming decreases soil seed bank and weed density in a paddy field. Agronomy 2019, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- MAPA. Anuarios Estadísticos. Available online: https://www.mapa.gob.es/es/estadistica/temas/publicaciones/anuario-de-estadistica/default.aspx (accessed on 18 January 2021).
- Cirujeda, A.; Aibar, J.; Zaragoza, C. Remarkable changes of weed species in Spanish cereal fields from 1976 to 2007. Agron. Sustain. Dev. 2011, 31, 675–688. [Google Scholar] [CrossRef]
- Marnotte, P. Influence des facteurs agroécologiques sur le developpement des mauvaises herbes en climat tropical humide. In Proceedings of the 1984 7ème Colloque International Ecologie, Biologie et Systematique des Mauvaises Herbes, Paris, France, 9–11 October 1984; COLUMA/EWRS: Paris, France, 1984; pp. 183–188. [Google Scholar]
- Alberti, J. L’arròs. Principals Malalties, Plagues i Males Herbes; BASF España: Barcelona, Spain, 1999. [Google Scholar]
- Puente, J. Guía de La Flora de La Depresión Del Ebro. Serie Difusión; Consejo Protección de la Naturaleza de Aragón, Gobierno de Aragón: Zaragoza, Spain, 2004. [Google Scholar]
- Recasens, J.; Conesa, J.A. Malas Hierbas en Plántula. Guía de Identificación; Universitat de Lleida and Bayer CropScience: Lleida, Spain, 2009. [Google Scholar]
- R Core Development Team. R: A Language and Environment for Statistical Computing; R Core Development Team: Vienna, Austria, 2019. [Google Scholar]
- Pardo, G.; Marí, A.I.; Aibar, J.; Vilaplana, L.; Cirujeda, A. Bastard Cabbage (Rapistrum rugosum L.) resistance to tribenuron-methyl and iodosulfuron-methyl-sodium in Spain and alternative herbicides for its control. Agronomy 2019, 9, 492. [Google Scholar] [CrossRef] [Green Version]
- MAAMA. Guía de Gestión Integrada de Plagas. Cereal. Available online: https://www.mapa.gob.es/es/agricultura/temas/sanidad-vegetal/GUIAdeCEREALESWEB_tcm30-57956.pdf (accessed on 18 October 2018).
- Vasconcelos, T.; Forte, P. Weeds in the rice fields of Sado valley. In Proceedings of the VII Congreso de la Sociedad Española de Malherbología, Logroño, Spain, 23–25 November 1999; Suso Martínez de Bujo, M., Pardo Iglesias, A., Eds.; Sociedad Española de Malherbologia: Logroño, Spain, 1999; pp. 394–403. [Google Scholar]
- Golmohammadi, M.J.; Mohammaddoust Chamanabad, H.R.; Yaghoubi, B.; Oveisi, M. Rice weed community composition and richness in northern Iran: A temperate rainy area. Appl. Ecol. Environ. Res. 2018, 16, 4605–4617. [Google Scholar] [CrossRef]
- FAO. Annexe 1. Crop Salt Tolerance Data. Available online: http://www.fao.org/3/y4263e/y4263e0e.htm (accessed on 21 October 2020).
- Yuan, Z.Q.; Yu, K.L.; Wang, B.X.; Zhang, W.Y.; Zhang, X.L.; Siddique, K.H.M.; Stefanova, K.; Turner, N.C.; Li, F.M. Cutting improves the productivity of lucerne-rich stands used in the revegetation of degraded arable land in a semi-arid environment. Sci. Rep. 2015. [Google Scholar] [CrossRef] [Green Version]
- Carretero, J.L. Flora Arvense Española. Las Malas Hierbas de Los Cultivos Españoles; Phytoma-España: Valencia, Spain, 2004. [Google Scholar]
- Hutchinson, I.; Colosi, J.; Lewin, R.A. The biology of Canadian weeds. 63. Sonchus asper (L.) Hill. Biol. Can. Weeds 1984, 64, 731–744. [Google Scholar] [CrossRef] [Green Version]
- Hanslin, H.M.; Eggen, T. Salinity tolerance during germination of seashore halophytes and salt-tolerant grass cultivars. Seed Sci. Res. 2005, 15, 43–50. [Google Scholar] [CrossRef]
- Pastor Piñero, J.; Hernández, A.J. Old landfills as emergent vulnerable ecosystems to salinization: Soil characteristics and response of the plant species to increasing Na and anion contents. In Proceedings of the Global Forum on Salinization and Climate Change (GFSCC2010), Valencia, Spain, 25–29 October 2010; pp. 239–248. [Google Scholar]
- Lombardi, T.; Fochetti, T.; Onnis, A. Comparative salt tolerance of two wild Hordeum species (H. maritimum With. and H. murinum L.) from the coast of Tuscany (Italy). Plant Biosyst. 2000, 134, 333–339. [Google Scholar] [CrossRef]
- Wattl, S.; Vilar, L. A comparative study of the vegetation at Aiguamolls de l’Empordà wetlands (NE Iberian Peninsula). Sci. Gerundensis 1997, 23, 19–154. [Google Scholar]
- Turki, Z.; Sheded, M. Some observations on the weed flora of rice fields in the Nile Delta, Egypt. Feddes Repert. 2002, 113, 394–403. [Google Scholar] [CrossRef]
- Recasens, J.; Calvet, V.; Cirujeda, A.; Conesa, J.A. Phenological and demographic behaviour of an exotic invasive weed in agroecosystems. Biol. Invasions 2005, 7, 17–27. [Google Scholar] [CrossRef]
- Osca, J. Estudio de las malas hierbas más problemáticas de los arrozales valencianos. In Herbologia e Biodiversidade numa Agricultura Sustentável, Proceedings of the XII Congreso da Sociedad Española de Malherbología (SEMh), Lisboa, Portugal, 10–13 November 2009; ISA Press: Lisboa, Portugal, 2009; pp. 289–290. [Google Scholar]
- Lewin, R.A. Biological flora of the British Isles. J. Ecol. 1948, 36, 203–233. [Google Scholar] [CrossRef]
- Barrett, S.C.H.; Wilson, B.F. Colonizing ability in the Echinochloa crus-galli complex (barnyard grass). I. Variation in life history. Can. J. Bot. 1981, 59, 1844–1860. [Google Scholar] [CrossRef]
- Travlos, I.S.; Economou, G.; Kanatas, P.J. Corn and barnyardgrass competition as influenced by relative time of weed emergence and corn hybrid. Agron. J. 2011, 103, 1–6. [Google Scholar] [CrossRef]
- Osca, J.M. Expansion of Leptochloa fusca ssp. uninervia and Leptochloa fusca ssp. fascicularis in rice fields in Valencia, eastern Spain. Weed Res. 2013, 53, 479–488. [Google Scholar] [CrossRef]
- Vladimirov, V.; Delcheva, M. First record of the alien Diplachne fascicularis (Poaceae) in Bulgaria. Flora Mediterr. 2016, 26. [Google Scholar] [CrossRef]
- Lloveras, J.; Melines, M.A. La calidad en la alfalfa. Posibles clasificaciones. Vida Rural 2015, 212, 36–40. [Google Scholar]
- MAAMA. Guía de Gestión Integrada de Plagas. Arroz. Available online: https://www.mapa.gob.es/es/agricultura/temas/sanidad-vegetal/guia_arroz_webprotegida_tcm30-434395.pdf (accessed on 18 October 2018).
- Reddy, T.Y.; Reddi, G.H.S. Principles of Agronomy, 3rd ed.; Kalyani Publishers: New Delhi, India, 2002. [Google Scholar]
- Dass, A.; Shekhawat, K.; Choudhary, A.K.; Sepat, S.; Rathore, S.S.; Mahajan, G.; Chauhan, B.S. Weed management in rice using crop competition—A review. Crop Prot. 2017. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Name | 2012 (Ns) | 2013 (Ns) | 2014 | 2015 | 2016 |
| Plots with the same crop in 2014–2016 | |||||
| F1 (Fescue 1) | Rice | Nc | Fescue | Fescue | Fescue |
| F2 (Fescue 2) | Rice | Nc | Fescue | Fescue | Fescue |
| Rg1 (Ryegrass 1) | Rice | Ryegrass | Ryegrass | Ryegrass | Ryegrass |
| Rg2 (Ryegrass 2) | Nc | Ryegrass | Ryegrass | Ryegrass | Ryegrass |
| C1 (Cereal 1) | Rice | Nc | Barley | Barley | Wheat |
| C2 (Cereal 2) | Nc | Wheat | Wheat | Wheat | Wheat |
| R1 (Rice 1) | Rice | Rice | Rice (Ns) | Rice | Rice |
| R2 (Rice 2) | Rice | Rice | Rice (Ns) | Rice | Rice |
| Plots with different crops in 2014–2016 | |||||
| CRg+F (Cereal-ryegrass+fescue) | Rice | Barley | Barley | Ryegrass and fescue | Ryegrass and fescue |
| FC1 (Fescue-cereal 1) | Nc | Wheat | Fescue | Wheat | Wheat |
| FC2 (Fescue-cereal 2) | Nc | Wheat | Fescue | Fescue | Wheat |
| FC3 (Fescue-cereal 3) | Nc | Rice | Fescue | Fescue | Wheat |
| FC4 (Fescue-cereal 4) | Nc | Rice | Fescue | Fescue | Wheat |
| LC (Lucerne-cereal) | Nc | Rice | Lucerne | Lucerne | Wheat |
| CR (Cereal-rice) | Nc | Rice | Wheat | Rice | Rice |
| FRNcL (Fescue-rice-Nc–lucerne) | Nc | Fescue | Rice | Nc | Lucerne |
| FR (Fescue-rice) | Nc | Fescue | Fescue (Ns) | Rice | Rice |
| FRRg1 (Fescue-rice-ryegrass 1) | Rice | Fescue | Fescue (Ns) | Rice | Nc, ryegrass |
| FRRg2 (Fescue-rice-ryegrass 2) | Nc | Fescue | Fescue (Ns) | Rice | Nc, ryegrass |
| RNc (Rice-Nc) | Rice | Rice | Rice (Ns) | Rice | Nc |
| Plot */year | 2014 | 2015 | 2016 |
| C1 | pinoxaden (barley) | - | - |
| R1 | - | Oxadiazon + penoxsulam + azimsulfuron | Oxadiazon + penoxsulam + azimsulfuron |
| R2 | - | Oxadiazon + penoxsulam + azimsulfuron | Oxadiazon + penoxsulam + azimsulfuron |
| CR | - | Profoxidim + penoxsulam | profoxdim + penoxsulam |
| FR | - | Oxadiazon + penoxsulam + bispyribac | penoxsulam |
| FRRg1 | - | Oxadiazon + cihalofop-p-butil + penoxsulam + bispyribac | - |
| FRRg1 | - | Oxadiazon + cihalofop-p-butil + penoxsulam + bispyribac | - |
| RNc | cihalofop-p-butil + penoxsulam + bispyribac | cihalofop-p-butil + penoxsulam + bispyribac | - |
| Plot * | Sand (%) | Coarse Loam (%) | Fine Loam (%) | Clay (%) | Texture | pH | Salinity ** (dS m−1) | Organic Matter (g/100 g) | P (mg kg−1) | K (mg kg−1) | Mg (mg kg−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| F1 | 10.43 | 6.42 | 43.03 | 40.12 | silty clay | 8.6 | 0.2 | 3.46 | 20 | 162 | 232 |
| F2 | 4.69 | 11.37 | 59.92 | 24.02 | silt loam | 8.8 | 0.2 | 1.88 | 4 | 96 | 226 |
| Rg1 | 22.68 | 11.49 | 35.81 | 30.02 | clay loam | 8.8 | 0.3 | 2.94 | 5 | 212 | 340 |
| Rg2 | 8.73 | 8.93 | 47.82 | 34.52 | silty clay loam | 8.9 | 0.3 | 2.15 | 7 | 158 | 274 |
| C1 | 5.19 | 8.48 | 55.29 | 31.04 | silty clay loam | 8.4 | 0.3 | 2.29 | 20 | 262 | 220 |
| C2 | 11.98 | 10.5 | 44.16 | 33.36 | silty clay loam | 8.6 | 0.4 | 2.24 | 6 | 292 | 270 |
| R1 | 20.06 | 7.39 | 37.82 | 34.73 | clay loam | 8.4 | 0.4 | 2.88 | 32 | 254 | 311 |
| R2 | 4.23 | 4.79 | 44.76 | 46.27 | silty clay | 8.4 | 0.3 | 3.38 | 32 | 348 | 478 |
| CRgF | 3.6 | 4.4 | 54.9 | 37.1 | silty clay loam | 8.5 | 0.6 | 2.59 | 6 | 236 | 401 |
| FC1 | 10.7 | 8.33 | 45.79 | 35.18 | silty clay loam | 8.8 | 0.3 | 2.22 | 12 | 158 | 264 |
| FC2 | 11.67 | 10.27 | 45.16 | 32.9 | silty clay loam | 8.4 | 0.9 | 2.01 | 8 | 182 | 259 |
| FC3 | 32.74 | 8.16 | 29.6 | 29.5 | clay loam | 8.6 | 0.3 | 2.51 | 9 | 148 | 242 |
| FC4 | 35.72 | 6.47 | 26.37 | 31.44 | clay loam | 8.5 | 0.3 | 2.95 | 19 | 268 | 192 |
| LC | 32.94 | 5.77 | 30.75 | 30.54 | clay loam | 8.6 | 0.3 | 2.50 | 10 | 248 | 304 |
| CR | 8.64 | 5.83 | 41.81 | 43.72 | silty clay | 8.6 | 0.3 | 3.53 | 39 | 328 | 354 |
| FRNcL | 8.27 | 6.57 | 43.63 | 41.53 | silty clay | 8.5 | 0.3 | 3.73 | 14 | 256 | 354 |
| FR | 4.01 | 2.1 | 36.87 | 57.02 | clay | 8.7 | 0.4 | 2.88 | 16 | 320 | 548 |
| FRRg1 | 17.8 | 7.3 | 40.6 | 34.3 | silty clay loam | 9.0 | 0.3 | 1.86 | 11 | 158 | 394 |
| FRRg2 | 36.5 | 6.87 | 27.18 | 29.35 | clay loam | 8.8 | 0.2 | 2.51 | 17 | 102 | 234 |
| RNc | 9.49 | 15.38 | 46.25 | 28.88 | silty clay loam | 8.7 | 0.2 | 2.01 | 22 | 174 | 272 |
| Plot * | Electrical Conductivity ** (dS m−1) | Saturation (%) | Ca (meql−1) | Mg (meql−1) | Na (meql−1) |
|---|---|---|---|---|---|
| C2 | 2.27 | 54.75 | 10.21 | 3.18 | 9.79 |
| R1 | 2.38 | 50.00 | 14.10 | 3.45 | 7.90 |
| CRgF | 3.62 | 60.25 | 12.52 | 5.75 | 21.17 |
| FC2 | 6.72 | 52.75 | 22.03 | 9.30 | 44.83 |
| FR | 1.52 | 69.25 | 5.56 | 2.65 | 8.30 |
| Crop | Season | 1 Mean Crop Soil Cover (%) | 1 Mean Weed Cover (%) | Mean Echinochloa spp. Cover (%) | 2 Mean Crop Soil Cover (%) | 2* Mean Weed Cover (%) | Mean Echinochloa spp. Cover (%) |
|---|---|---|---|---|---|---|---|
| Winter cereal | spring | 78 ± 6.2 A | 16 ± 5.4 A | 2 ± 1.7 | 77.9 a | 8.1 cd | 2 ± 1.7 |
| Fescue | spring | 84 a A | * 7.8 x (17 A) | 0 ± 0.0 | 81.8 a | 5.4 d | 0.5 ± 0.3 |
| summer | 82 a M | * 6.3 x (11 O) | 0.3 ± 0.3 | ||||
| autumn | 81 a X | * 2.7 x (6 Z) | 1 ± 1.1 | ||||
| Ryegrass | spring | 75 a AB | 32.5 x A | 0 ± 0.0 | 58.1 ab | 42.5 ab | 1.3 ± 1.3 |
| summer | 46 a N | 50.8 x MN | 0.1 ± 0.0 | ||||
| autumn | 53 a X | 57.8 x XY | 4 ± 2.3 | ||||
| Ryegrass+fescue | spring | 60 a ABC | 20.5 x A | 0 ± 0.0 | 62.2 ab | 13.6 bcd | 2 ± 1.2 |
| summer | 67 a MN | 13.9 x NO | 1.3 ± 1.30 | ||||
| autumn | 60 X | 12.7 x YZ | 4 ± 3.6 | ||||
| Lucerne | spring | 37 a BC | 49.6 x A | 0 ± 0.0 | 36.9 b | 55.7 a | 9 ± 5.2 |
| summer | 40 a N | 60.3 x M | 15 ± 12.7 | ||||
| autumn | 35 a X | 75.4 x X | 13 ± 9.3 | ||||
| Rice | spring | 36 b C | 28.8 x A | 12 ± 5.5 | 63.0 ab | 20.9 abc | 15 ± 4.6 |
| summer | 76 a M | 26.8 x MNO | 10 ± 7.4 | ||||
| autumn | 81 a X | 34.7 x XYZ | 21 ± 10.2 |
| 2014 | 2015 | 2016 | ||||
| Plot * | su | au | su | au | su | au |
| C | C | Rg + F | Rg + F | Rg + F | Rg + F | |
| CRF | 0.2 | 0 | 0 | 0 | 0 | 0 |
| C | C | R | R | R | R | |
| CR | 22.5 | 0.2 | 0 | 0 | 0 | 0 |
| C | C | C | C | C | C | |
| C1 | 0 | 0 | 0.2 | 0 | 0.2 | 0 |
| C2 | 0 | 0 | 0.2 | 22.5 | 0 | 0 |
| L | L | L | L | C | C | |
| LC | 0 | 0 | 0 | 0 | 11 | 0.2 |
| F | F | C | C | C | C | |
| FC1 | 0 | 0 | 4 | 4 | 0 | 0 |
| FC3 | 0 | 0 | 0 | 0 | 0 | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pardo, G.; Marí, A.I.; Aibar, J.; Cirujeda, A. Do Crop Rotations in Rice Reduce Weed and Echinochloa spp. Infestations? Recommendations for Integrated Weed Control. Agronomy 2021, 11, 454. https://doi.org/10.3390/agronomy11030454
Pardo G, Marí AI, Aibar J, Cirujeda A. Do Crop Rotations in Rice Reduce Weed and Echinochloa spp. Infestations? Recommendations for Integrated Weed Control. Agronomy. 2021; 11(3):454. https://doi.org/10.3390/agronomy11030454
Chicago/Turabian StylePardo, Gabriel, Ana Isabel Marí, Joaquín Aibar, and Alicia Cirujeda. 2021. "Do Crop Rotations in Rice Reduce Weed and Echinochloa spp. Infestations? Recommendations for Integrated Weed Control" Agronomy 11, no. 3: 454. https://doi.org/10.3390/agronomy11030454
APA StylePardo, G., Marí, A. I., Aibar, J., & Cirujeda, A. (2021). Do Crop Rotations in Rice Reduce Weed and Echinochloa spp. Infestations? Recommendations for Integrated Weed Control. Agronomy, 11(3), 454. https://doi.org/10.3390/agronomy11030454