
Potassium Application Positively Modulates Physiological Responses of Cocoa Seedlings to Drought Stress



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Experimental Design and Treatments
2.2.1. Potassium Treatments
2.2.2. Water Stress Treatments
2.3. Data Collection
2.4. Statistical Analyses
3. Results
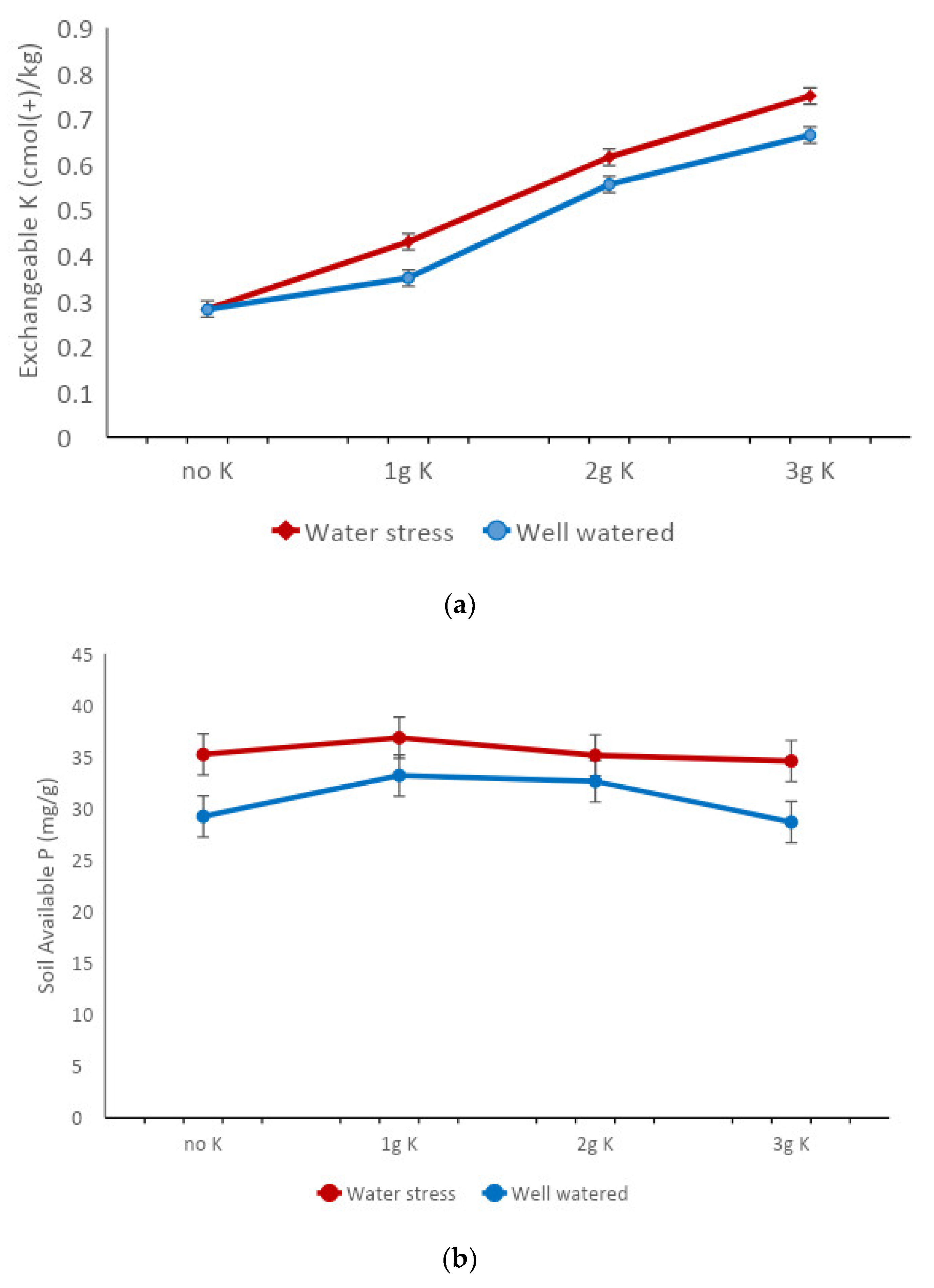
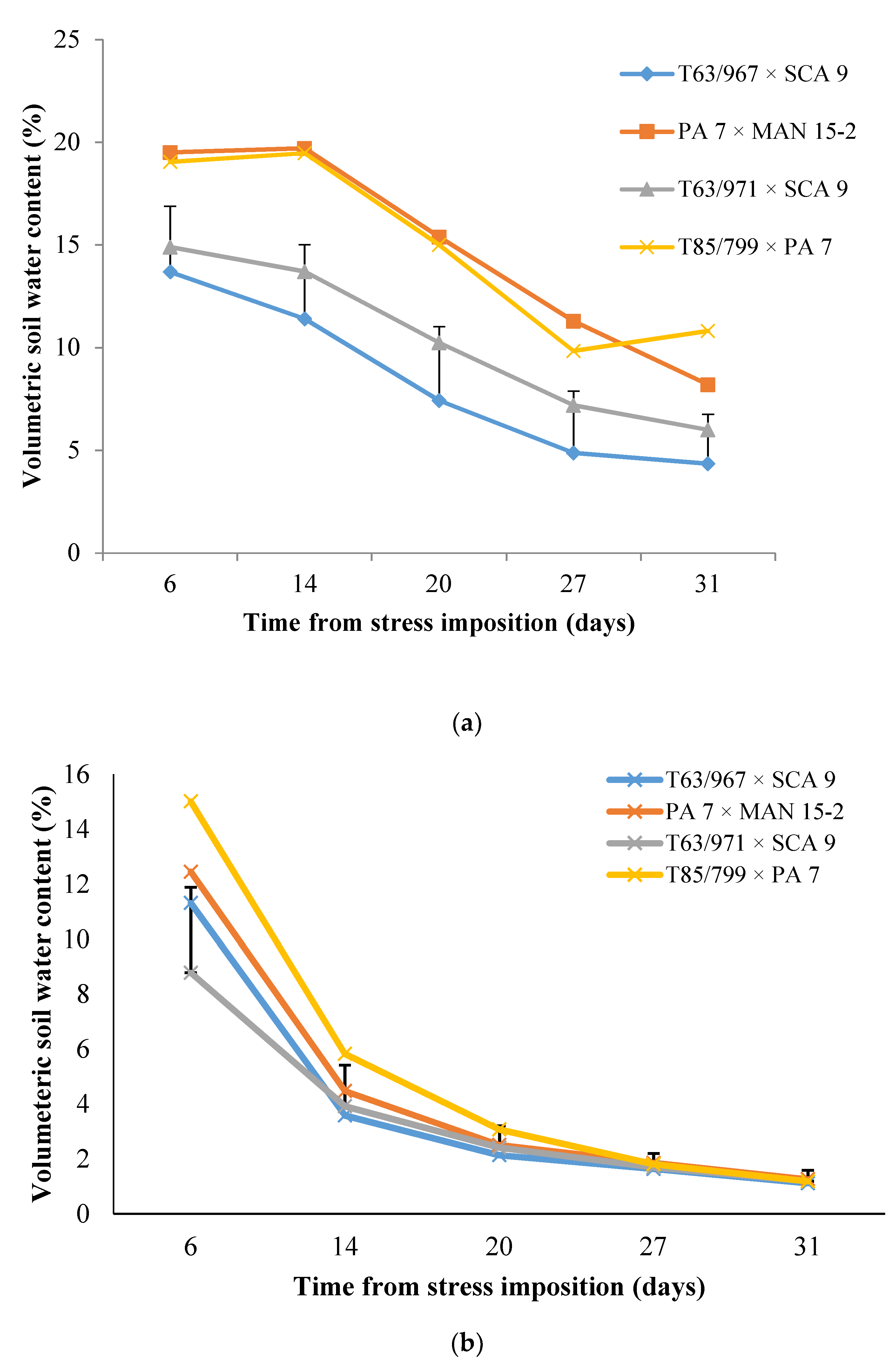
3.1. Effects of Variety and Water Stress Treatments on Contents of Soil Nutrients and Water
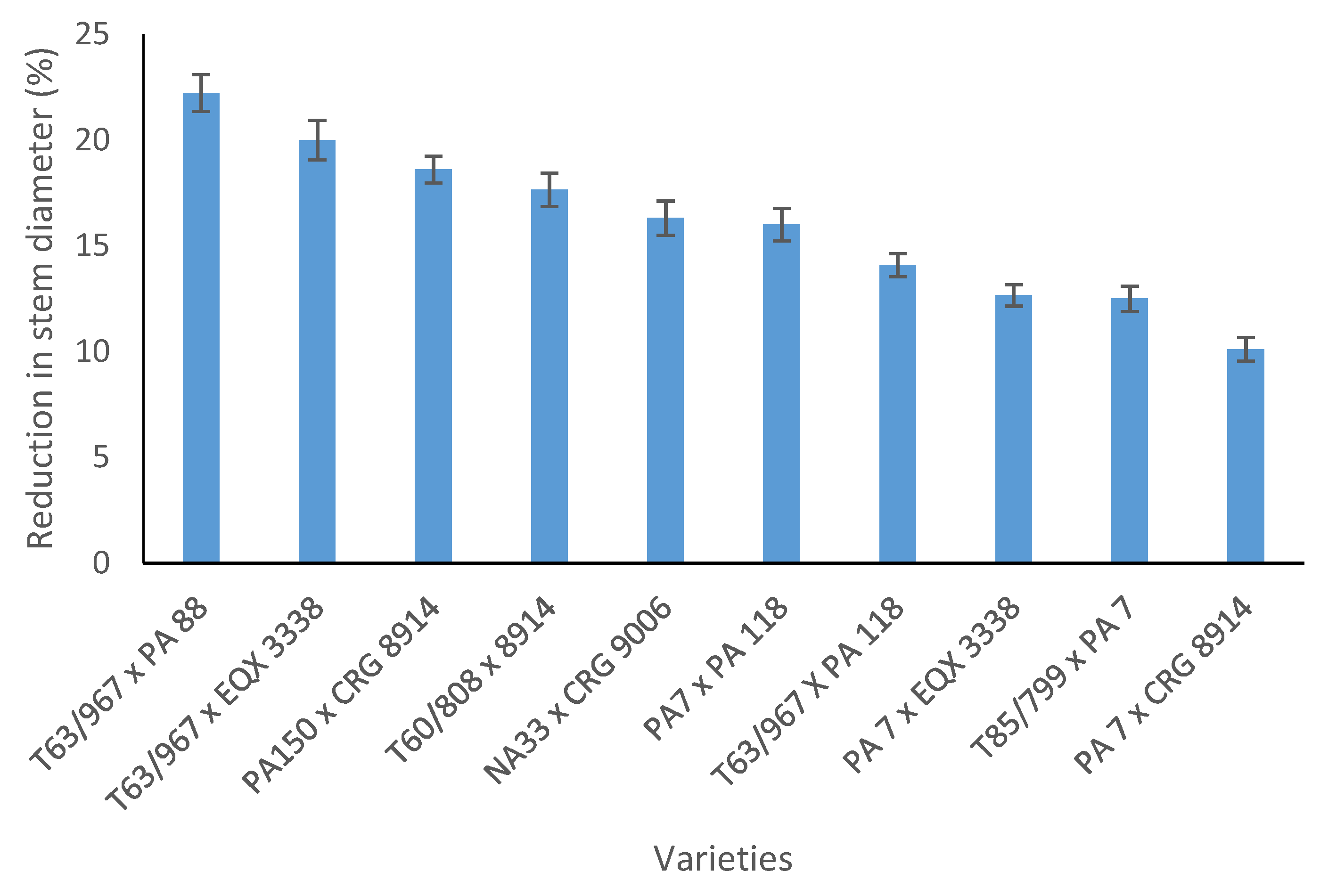
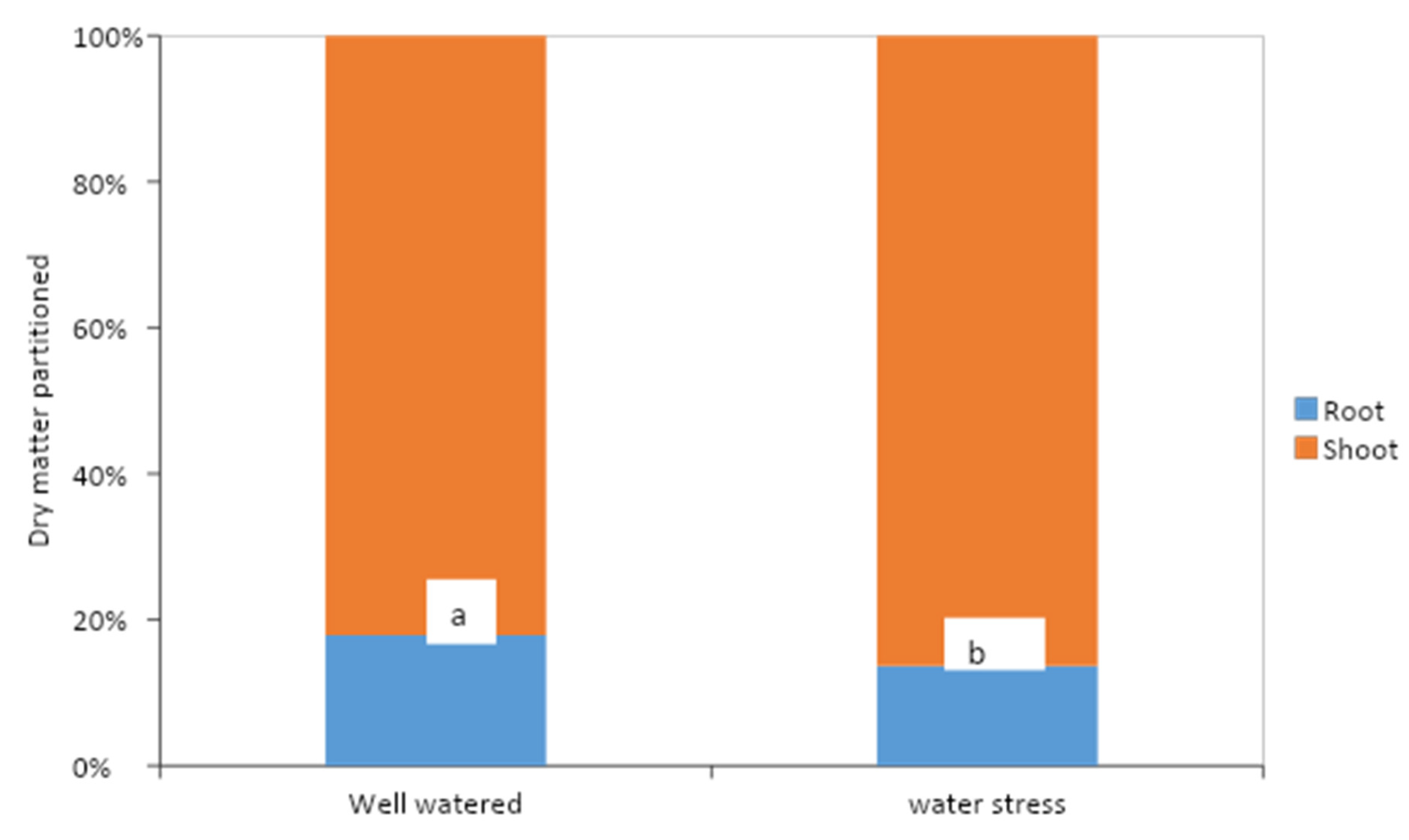
3.2. Effects of Water Stress and Potassium Application on Allometric Relationships of Cocoa Seedlings
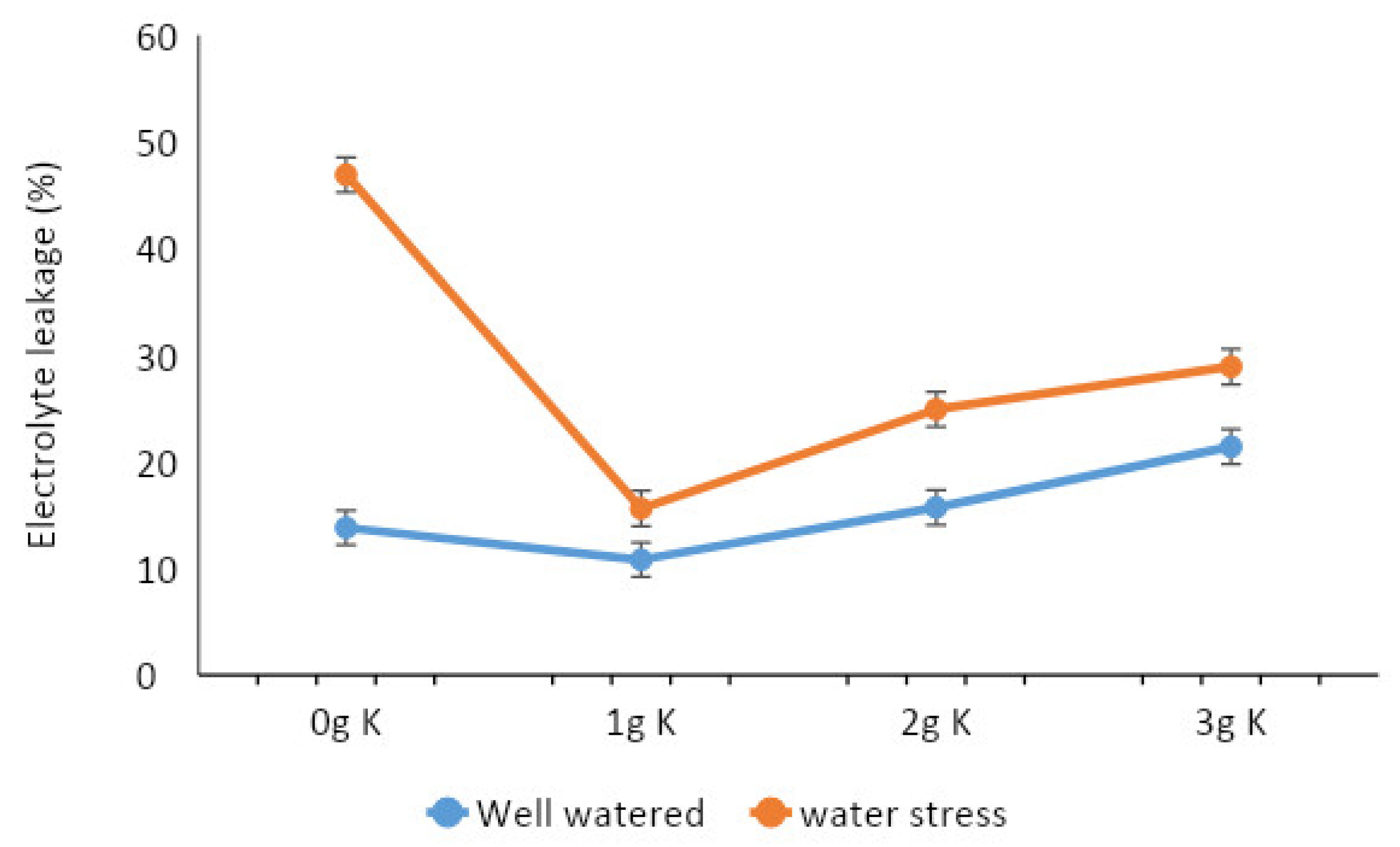
3.3. Effects of Potassium and Watering Regime on the Physiological Functions of Cocoa Varieties
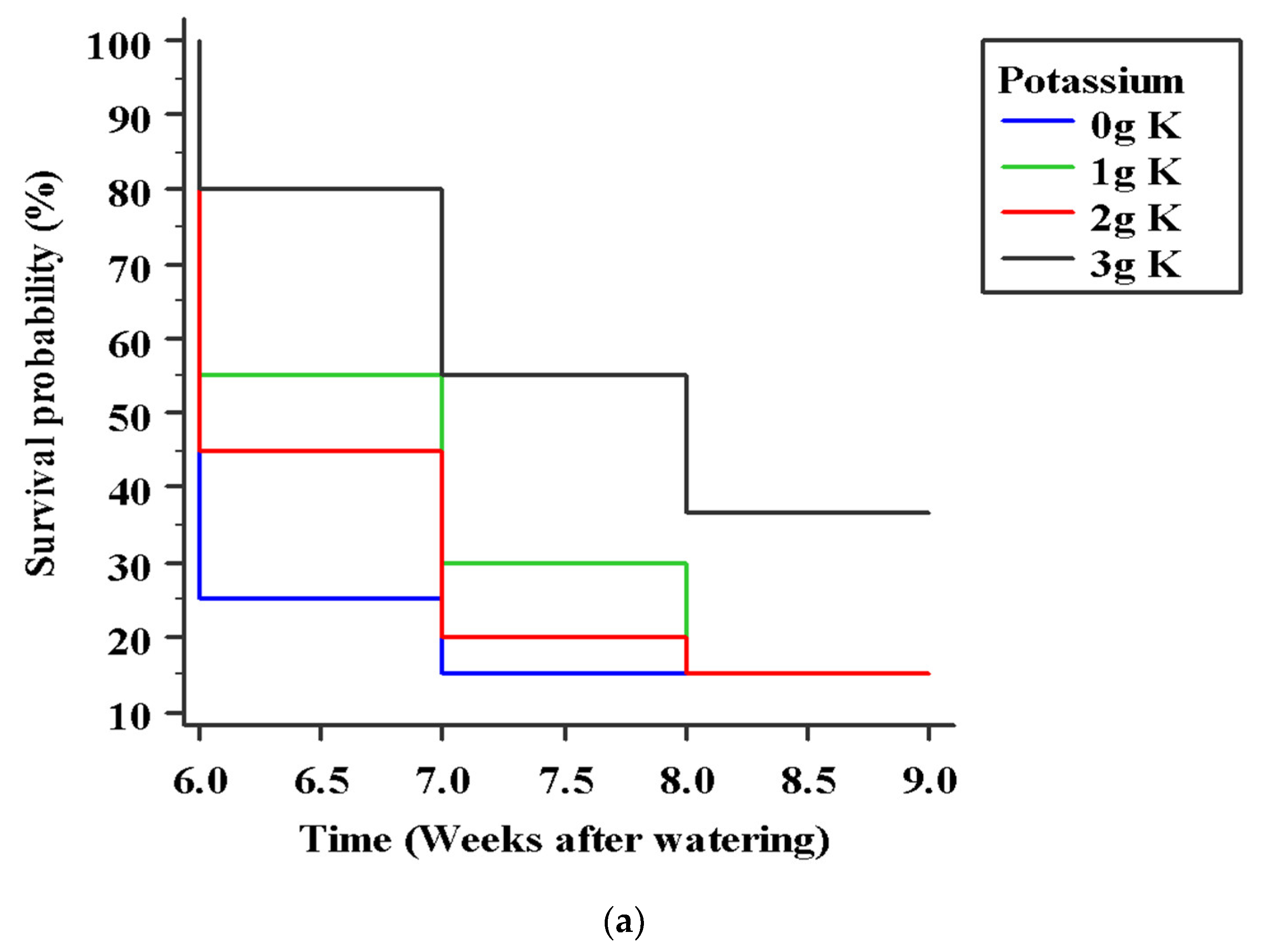
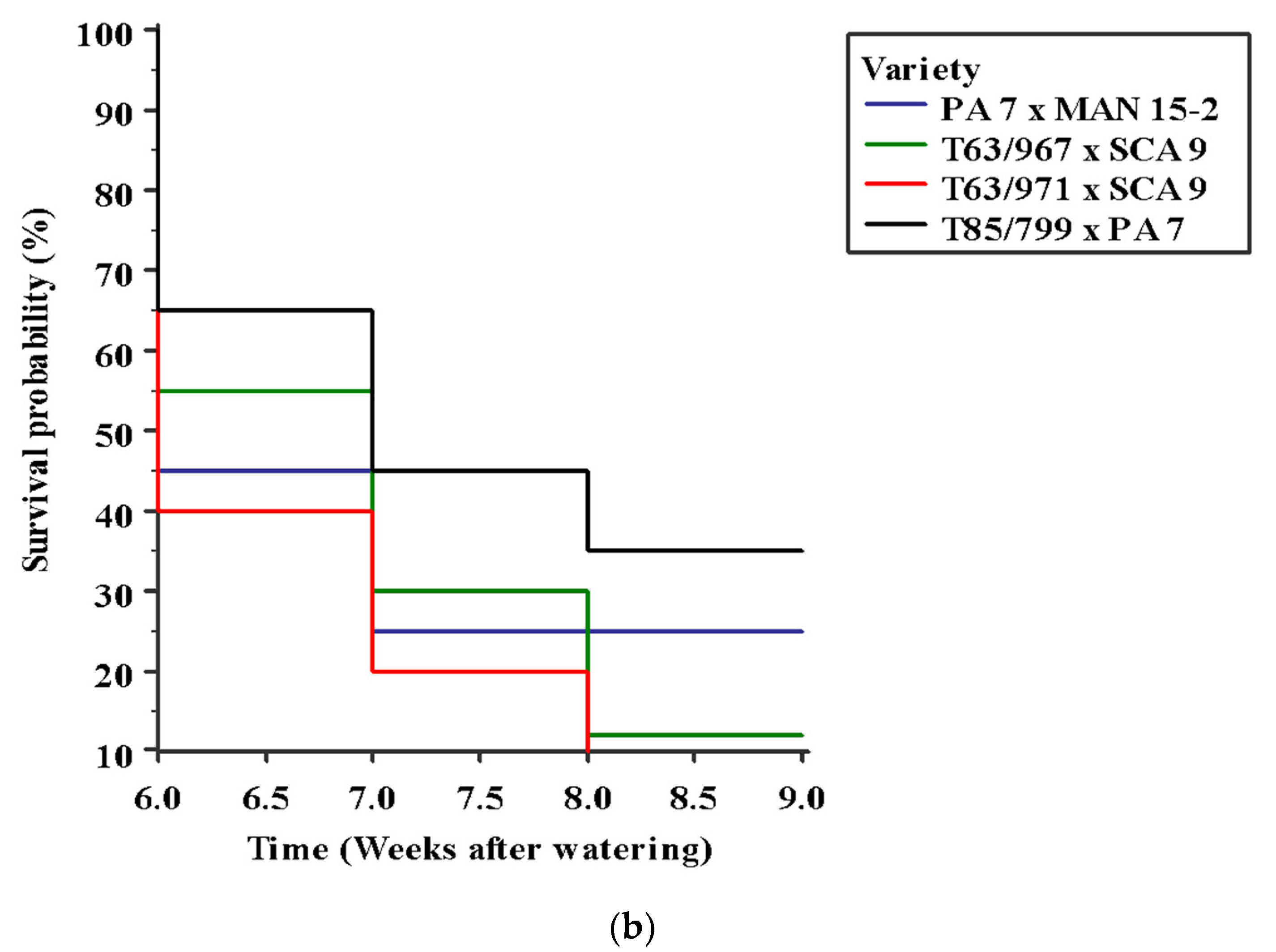
3.4. Effects of Variety and Potassium Application on Cocoa Seedling Survival under Water Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anim-Kwapong, G.J.; Frimpong, E.B. Vulnerability of Agriculture to Climate Change–Impact of Climate Change on Cocoa. 2005. Available online: https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.494.4508&rep=rep1&type=pdf (accessed on 1 May 2017).
- Läderach, P.; Martinez, A.; Schroth, G.; Castro, N. Predicting the future climatic suit-ability for cocoa farming of the world’s leading producer countries, Ghana and Côte d’Ivoire. Clim. Chang. 2013, 119, 841–854. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, I.C.; De Almeida, A.-A.F.; Anhert, D.; Da Conceição, A.S.; Pirovani, C.P.; Pires, J.L.; Valle, R.R.; Baligar, V.C. Molecular, Physiological and Biochemical Responses of Theobroma cacao L. Genotypes to Soil Water Deficit. PLoS ONE 2014, 9, e115746. [Google Scholar] [CrossRef] [Green Version]
- Ruf, F.; Schroth, G.; Doffangui, K. Climate change, cocoa migrations and deforestation in West Africa: What does the past tell us about the future? Sustain. Sci. 2015, 10, 101–111. [Google Scholar] [CrossRef]
- Carr, M.K.V.; Lockwood, G. The water relations and irrigation requirements of cocoa (Theobroma cacao L.): A review. Exp. Agric. 2011, 47, 653–676. [Google Scholar] [CrossRef]
- Schroth, G.; Läderach, P.; Martinez-Valle, A.I.; Bunn, C.; Jassogne, L. Vulnerability to climate change of cocoa in West Africa: Patterns, opportunities and limits to adaptation. Sci. Total. Environ. 2016, 556, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.B.; Gifford, R.M. Soil carbon stocks and land use change: A meta analysis. Glob. Chang. Biol. 2002, 8, 345–360. [Google Scholar] [CrossRef]
- Anim-Kwapong, G.J.; Frimpong, E.B. Vulnerability of agriculture to climate change- impact of climate change on cocoa production. In Report on Vulnerability and Adaptation Assessment under The Netherlands Climate Change Studies Assistance Programme Phase 2 (NCCSAP2); Cocoa Research Institute of Ghana: New Tafo–Akim, Ghana, 2006; p. 44. [Google Scholar]
- Oppong, F.; Opoku-Ameyaw, K.; Osei-Bonsu, K.; Amoah, F.; Brew, K.; Acheampong, K. The effect of time of planting at stake on cocoa seedling survival. Ghana J. Agric. Sci. 1999, 32, 79–86. [Google Scholar] [CrossRef]
- Oppong, F.; Osei-Bonsu, K.; Amoah, F.; Acheampong, K. Potential use of Flemingia macrophylla as mulch for managing weeds in young cocoa in Ghana. Ghana J. Agric. Sci. 1998, 31, 67–72. [Google Scholar] [CrossRef]
- Padi, F.K.; Adu-Gyamfi, P.; Akpertey, A.; Arthur, A.; Ofori, A. Differential response of cocoa families to field establishment stress. Plant Breed. 2013, 132, 229–236. [Google Scholar] [CrossRef]
- Ofori, A.; Padi, F.K.; Acheampong, K.; Lowor, S. Genetic variation and relationship of traits related to drought tolerance in cocoa (Theobroma cacao L.) under shade and no-shade conditions in Ghana. Euphytica 2014, 201, 411–421. [Google Scholar] [CrossRef]
- Asare, R.; Ræbild, A. Tree diversity and canopy cover in cocoa systems in Ghana. New For. 2016, 47, 287–302. [Google Scholar]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Rodriguez, D.; Andrade, F.; Goudriaan, J. Effects of phosphorus nutrition on tiller emergence in wheat. Plant Soil 1999, 209, 283–295. [Google Scholar] [CrossRef]
- Jensen, P. Effects of interrupted K+ supply on growth and uptake of K+, Ca2+, Mg2+ and Na+ in spring wheat. Physiol. Plant. 1982, 56, 259–265. [Google Scholar] [CrossRef]
- Elumalai, R.P.; Nagpal, P.; Reed, J.W. A Mutation in the Arabidopsis KT2/KUP2 Potassium Transporter Gene Affects Shoot Cell Expansion. Plant Cell 2002, 14, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egilla, J.N.; Davies, F.T.; Boutton, T.W. Drought stress influences leaf water content, photosynthesis, and water-use efficiency of Hibiscus rosa-sinensis at three potassium concentrations. Photosynthetica 2005, 43, 135–140. [Google Scholar] [CrossRef]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001. [Google Scholar]
- De la Guardia, M.D.; Benlloch, M. Effects of potassium and gibberellic acid on stem growth of whole sunflower plants. Physiol. Plant. 1980, 49, 443–448. [Google Scholar] [CrossRef]
- Blanchet, R.; Studer, R.; Chaumont, C. Quelques aspects des interactions entre l’alimentation potassique et l’alimentation hydrique des plantes. Ann. Agron. 1962, 13, 93. [Google Scholar]
- Benlloch-Gonzalez, M.; Arquero, O.; Fournier, J.M.; Barranco, D.; Benlloch, M. K+ starvation inhibits water-stress-induced stomatal closure. J. Plant Physiol. 2008, 165, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, G.; Gyamfi, M.M.O. The CRIG Cocoa Germplasm Collection with Notes on Codes Used in the Breeding Programme at Tafo and Elsewhere; Technical Bulletin, No. 10; Cocoa Research Institute of Ghana: Tafo, Ghana, 1979; p. 62. [Google Scholar]
- Padi, F.K.; Ofori, A.; Takrama, J.; Djan, E.; Opoku, S.Y.; Dadzie, A.M.; Bhattacharjee, R.; Motamayor, J.C.; Zhang, D. The impact of SNP fingerprinting and parentage analysis on the effectiveness of variety recommendations in cacao. Tree Genet. Genomes 2015, 11, 44. [Google Scholar] [CrossRef]
- Eskes, A.B. Collaborative and participatory approaches to cocoa variety improvement. In Final Report of the CFC/ICCO/Bioversity Project on Cocoa Productivity and Quality Improvement: A Participatory Approach (2004–2010); CFC: Amsterdam, The Netherlands; ICCO: London, UK; Bioversity International: Rome, Italy, 2011; p. 205. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining organic carbon in soils: Effect of variations in digestion conditions and of inorganic soil constituents. Soil Sci. 1934, 63, 251–263. [Google Scholar] [CrossRef]
- Van Reeuwijk, L.P. Procedures for Soil Analysis, 3rd ed.; Technical Paper 9; International Soil Reference and Information Centre: Wageningen, The Netherlands, 1992. [Google Scholar]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen-Total. In Methods of Soil Analysis. Part 2, 2nd ed.; Agronomy Monograph. 9; Page, A.L., Miller, R.H., Eds.; ASA: Madison, WI, USA; SSSA: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar]
- Truog, E. The Determination of the Readily Available Phosphorus of Soils 1. Agron. J. 1930, 22, 874–882. [Google Scholar] [CrossRef] [Green Version]
- Thomas, G.W. Exchangeable cations. In Methods of Soil Analysis, Part 2; Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; Soil Science Society of America, Madison Wisconsin USA: Madison, WL, USA, 1982. [Google Scholar]
- Yamasaki, S.; Dillenburg, L.R. Measurements of leaf relative water Content in Araucaria angustifolia. Rev. Bras. Fisiol. Veg. 1999, 11, 69–75. [Google Scholar]
- Ragazzi, E.; Veronese, G. Quantitative analysis of phenolic compounds after thin-layer chromatographic separation. J. Chromatogr. A 1973, 77, 369–375. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Gattward, J.N.; Almeida, A.-A.F.; Souza, J.O.; Gomes, F.P.; Kronzucker, H.J. Sodium–potassium synergism in Theobroma cacao: Stimulation of photosynthesis, water-use efficiency and mineral nutrition. Physiol. Plant. 2012, 146, 350–362. [Google Scholar] [CrossRef]
- Orchard, J.E. Effect of K on transpiration, leaf diffusive resistance and growth in seedlings of Theobroma cacao L. In Technical Report; Cepec/Ceplac: Ilhéus, Brazil, 1978; pp. 61–64. [Google Scholar]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; pp. 178–189. [Google Scholar]
- Romheld, V.; Kirkby, E.A. Research on potassium in agriculture: Needs and prospects. Plant Soil 2010, 335, 155–180. [Google Scholar] [CrossRef]
- Ahenkorah, Y.; Akrofi, G.S. Amazon cacao (Theobroma cacao L.) shade and manurial experiment (K2- 01) at the Cocoa Research Institute of Ghana. I. First five years. Agron. J. 1968, 60, 591–594. [Google Scholar] [CrossRef]
- Wessel, M. Fertilizer requirements of cacao (Theobroma cacao L.) in South-Western Nigeria. In Communication 61; Koninklijk Instituut voor de Tropen: Amsterdam, The Netherlands, 1971; p. 104. [Google Scholar]
- Ahenkorah, Y.; Halm, B.J.; Appiah, M.R.; Akrofi, G.S. Fertilizer use on cacao rehabilitation projects in Ghana. In Proceeding of the 8th International Cocoa Research Conference, Cartagena, Colombia, 18–23 October 1981; pp. 165–170. [Google Scholar]
- Kemp, W.P.; Moody, U.L. Relationships between Regional Soils and Foliage Characteristics and Western Spruce Budworm (Lepidoptera: Tortricidae) Outbreak Frequency. Environ. Entomol. 1984, 13, 1291–1297. [Google Scholar] [CrossRef]
- Díaz, E.; Roldan, A. Effects of reforestation techniques on the nutrient content, photosynthetic rate and stomatal conductance of Pinus halepensis seedlings under semiarid conditions. Land Degrad. Dev. 2000, 11, 475–486. [Google Scholar] [CrossRef]
- Steudle, E. Water uptake by roots: Effects of water deficit. J. Exp. Bot. 2000, 51, 1531–1542. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Sun, S.; Yang, L.; Li, M.; Ma, F.; Zou, Y. Potassium Uptake and Transport in Apple Roots Under Drought Stress. Hortic. Plant J. 2019, 5, 10–16. [Google Scholar] [CrossRef]
- Ávila-Lovera, E.; Coronel, I.; Jaimez, R.E.; Urich, R.; Pereyra, G.; Araque, O.; Chacón, I.; Tezara, W. Ecophysiological traits of adult trees of Criollo cocoa cultivars (Theobroma cacao L.) from a germplasm bank in Venezuela. Exp. Agric. 2015, 52, 137–153. [Google Scholar] [CrossRef]
- De Almeida, A.A.F. Some water relations aspects of Theobroma cocoa clones. In Proceedings of the 13th International Cocoa Research Conference, Kota Kinabalu, Malaysia, 8–14 October 2000; pp. 349–363. [Google Scholar]
- Rolny, N.; Costa, L.; Carrión, C.; Guiamet, J.J. Is the electrolyte leakage assay an unequivocal test of membrane deterioration during leaf senescence? Plant Physiol. Biochem. 2011, 49, 1220–1227. [Google Scholar] [CrossRef]
- Percival, G.C.; Keary, I.P.; Al-Habsi, S. An assessment of the drought tolerance of Fraxinus genotypes for urban landscape plantings. Urban For. Urban Green. 2006, 5, 17–27. [Google Scholar] [CrossRef]
- Banks, J.M. Chlorophyll fluorescence as a tool to identify drought stress in Acer genotypes. Environ. Exp. Bot. 2018, 155, 118–127. [Google Scholar] [CrossRef]
- Lim, P.O.; Kim, H.J.; Gil Nam, H. Leaf Senescence. Annu. Rev. Plant Biol. 2007, 58, 115–136. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.-H.; Zhu, J.-K. Phenotypic Analysis of Arabidopsis Mutants: Electrolyte Leakage after Freezing Stress. Cold Spring Harb. Protoc. 2010, 2010, 4970. [Google Scholar] [CrossRef]
- De Abreu, I.N.; Mazzafera, P. Effect of water and temperature stress on the con-tent of active constituents of Hypericum brasiliense Choisy. Plant Physiol. Biochem. 2005, 43, 241–248. [Google Scholar] [CrossRef]
- Jaafar, H.Z.; Ibrahim, M.H.; Fakri, N.F.M. Impact of soil field water capacity on secondary metabolites, phenylalanine ammonia-lyase (PAL), malondialdehyde (MDA) and photosynthetic responses of Malaysian Kacip Fatimah (Labisia pumila Benth). Molecules 2012, 17, 7305–7322. [Google Scholar] [CrossRef] [Green Version]
- Anjum, S.A.; Farooq, M.; Xie, X.-Y.; Liu, X.-J.; Ijaz, M.F. Antioxidant defense system and proline accumulation enables hot pepper to perform better under drought. Sci. Hortic. 2012, 140, 66–73. [Google Scholar] [CrossRef]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.R.; Chaitanyaa, K.V.; Vivekanandan, M. Drought-induced responsesof photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Hatier, J.-H.B.; Gould, K.S. Foliar anthocyanins as modulators of stress signals. J. Theor. Biol. 2008, 253, 625–627. [Google Scholar] [CrossRef]
- Agati, G.; Tattini, M. Multiple functional roles of flavonoids in photoprotection. New Phytol. 2010, 186, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Sulpice, R. Closing the yield gap: Can metabolomics be of help? J. Exp. Bot. 2019, 71, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Alvim, P.T. Las necesidades de agua del cacao. Turrialba 1960, 10, 6–16. [Google Scholar]












{kind=link}
{kind=link}
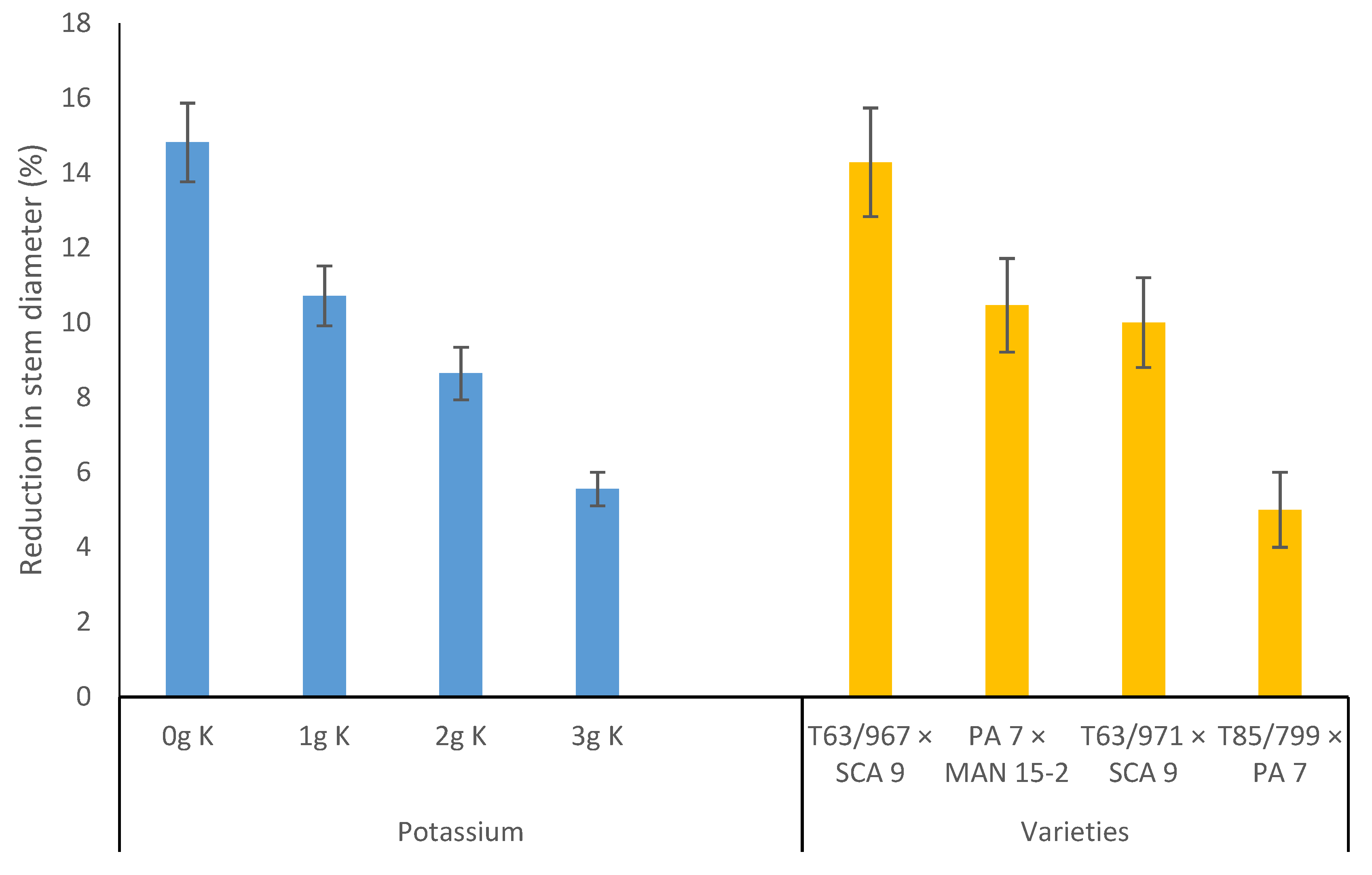{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Use in this Trial | Source/Parentage |
|---|---|---|
| T63/967 × SCA 9 | Experiment 1 | T63/967 was derived from a Nanay × Iquitos cross collected by Posnette from Trinidad in 1944 *. SCA 9 is a clone of Upper Amazon (Scavina) origin |
| T63/971 × SCA 9 | Experiment 1 | T63/971 was derived from a Nanay × Iquitos cross collected by Posnette from Trinidad in 1944. |
| PA 7 × MAN 15-2 | Experiment 1 | PA 7 is a clone of Parinari origin, belonging to the Marañón genetic group. Man 15-2 is a clone selection from an international clone trial ‡. Mainly of Purús and Amelonado origin † |
| T85/799 × PA 7 | Experiments 1&2 | T85/799 was derived from a Nanay × Iquitos (NA 34 × IMC 60) cross collected by Posnette from Trinidad in 1944. |
| NA33 × CRG 9006 | Experiment 2 | NA 33 is a clone introduction from the Nanay genetic group. CRG 9006 is clone selection made in Ghana from GU 144/C × EQX 3338. GU 144/C is a clone of Guiana origin |
| PA 7 × CRG 8914 | Experiment 2 | PA 7 belongs to the Marañón genetic group. CRG 8914 is a clone selection from a cross made in Ghana from T85/799 × CAS 3. CAS 3 is an Amelonado clone |
| PA 7 × EQX 3338 | Experiment 2 | EQX 3338 is a clone introduction originating from Ecuador. Belongs to the Trinitario genetic group † |
| PA150 × CRG 8914 | Experiment 2 | PA 150 belongs to the Marañón genetic group. CRG 8914 was derived from T85/799 × CAS 3. |
| PA 7 × PA 118 | Experiment 2 | Both PA 7 and PA 118 belong to the Marañón genetic group. |
| T60/887 × CRG 8914 | Experiment 2 | T60/887 was derived from Marañón × Nanay (PA 7 × NA 32) cross made in Trinidad and collected by Posnette in 1944. Predominantly a Nanay † |
| T63/967 × EQX 3338 | Experiment 2 | T63/967 was derived from a Nanay × Iquitos cross. EQX 3338 is a Trinitario clone |
| T63/967 × PA 118 | Experiment 2 | T63/967 was derived from a Nanay × Iquitos cross. PA 118 belongs to the Marañón genetic group. |
| T63/967 × PA 88 | Experiment 2 | T63/967 was derived from a Nanay × Iquitos cross. PA 88 belongs to the Marañón genetic group. |
| Soil Property | Initial Contents | Final Contents | ||
|---|---|---|---|---|
| Water Stress | Well-Watered | P | ||
| Organic C (%) | 1.160 ± 0.04 | 1.262 ± 0.030 | 1.237 ± 0.034 | 0.415 |
| pH | 5.90 ± 0.03 | 6.14 ± 0.027 | 6.13 ± 0.03 | 0.817 |
| Total N (%) | 0.105 ± 0.004 | 0.109 ± 0.002 | 0.107 ± 0.002 | 0.499 |
| Available P (mg/kg) | 49.46 ± 4.00 | 35.46 ± 1.38 | 30.94 ± 1.65 | 0.030 |
| Exchangeable K (cmol/kg) | 0.67 ± 0.020 | 1.164 ± 0.042 | 1.041 ± 0.033 | <0.001 |
| Ca (cmol/kg) | 4.72 ± 0.18 | 4.99 ± 0.11 | 5.12 ± 0.11 | 0.085 |
| Mg (cmol/kg) | 2.18 ± 0.11 | 1.74 ± 0.066 | 1.73 ± 0.074 | 0.779 |
| Muriate of Potassium (g/plant) | Stem Diameter (mm) | Length of Tap Root (cm) | Dry Weight of Roots (g) | Dry Weight of Stem (g) | Dry Weight of Leaves (g) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Watered | Stressed | Watered | Stressed | Watered | Stressed | Watered | Stressed | Watered | Stressed | |
| 0 | 8.1 | 6.9 | 44.9 | 29.9 | 6.9 | 2.9 | 7.3 | 5.2 | 12.5 | 8.5 |
| 1 | 8.4 | 7.5 | 33.7 | 30.8 | 5.8 | 3.1 | 8.1 | 5.3 | 12.6 | 8.9 |
| 2 | 8.1 | 7.4 | 40.9 | 32.1 | 4.7 | 3.6 | 7.5 | 5.6 | 12.9 | 9.0 |
| 3 | 9.0 | 8.5 | 36.4 | 34.5 | 5.9 | 4.4 | 8.3 | 6.4 | 13.1 | 9.0 |
| SEDdf=32 | 0.29 | 0.23 | 4.21 | 2.33 | 0.62 | 0.22 | 0.71 | 0.38 | 0.86 | 0.67 |
| Varieties | ||||||||||
| T63/967 × SCA 9 | 9.1 | 7.8 | 44.9 | 37.3 | 6.9 | 3.6 | 10.1 | 6.4 | 15.4 | 9.5 |
| T63/971 × SCA 9 | 8.0 | 7.2 | 40.9 | 29.6 | 4.7 | 2.8 | 7.3 | 5.3 | 12.7 | 9.1 |
| PA 7 × MAN 15-2 | 8.6 | 7.7 | 33.7 | 31.7 | 5.8 | 4.0 | 7.2 | 5.7 | 12.0 | 9.2 |
| T85/799 × PA 7 | 8.0 | 7.6 | 36.4 | 28.7 | 5.9 | 3.8 | 6.4 | 5.1 | 11.0 | 7.5 |
| SEDdf=32 | 0.29 | 0.23 | 4.21 | 2.33 | 0.62 | 0.22 | 0.71 | 0.38 | 0.86 | 0.67 |
| Muriate of Potassium (g/plant) | Stem Diameter (mm) | Length of Tap Root (cm) | Dry Weight of Roots (g) | Dry Weight of Stem (g) | Dry Weight of Leaves (g) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Watered | Stressed | Watered | Stressed | Watered | Stressed | Watered | Stressed | Watered | Stressed | |
| 0 | 7.9 | 6.7 | 34.4 | 25.2 | 5.6 | 1.8 | 8.5 | 3.4 | 14.7 | 8.6 |
| 1 | 8.3 | 6.7 | 34.3 | 26.5 | 5.5 | 1.9 | 8.7 | 3.7 | 15.9 | 9.0 |
| 2 | 8.1 | 6.9 | 31.5 | 30.3 | 5.0 | 2.1 | 8.6 | 3.7 | 15.8 | 8.9 |
| 3 | 8.4 | 7.1 | 30.1 | 31.6 | 5.1 | 2.2 | 8.7 | 3.8 | 16.5 | 9.5 |
| SEDdf=78 | 0.14 | 0.15 | 2.85 | 2.08 | 0.33 | 0.15 | 0.51 | 0.18 | 0.50 | 0.34 |
| Varieties | ||||||||||
| NA33 × CRG 9006 | 9.2 | 7.7 | 35.0 | 29.1 | 4.8 | 2.1 | 7.5 | 4.1 | 14.5 | 9.8 |
| PA 7 × CRG 8914 | 8.9 | 8.0 | 37.1 | 32.1 | 4.8 | 2.2 | 7.2 | 4.0 | 16.5 | 10.5 |
| T63/967 × EQX 3338 | 8.0 | 6.4 | 29.3 | 25.1 | 4.4 | 2.0 | 7.2 | 3.1 | 12.8 | 8.1 |
| T85/799 × PA 7 | 8.0 | 7.0 | 33.4 | 28.9 | 6.2 | 2.3 | 9.0 | 4.1 | 16.6 | 9.0 |
| PA150 × CRG 8914 | 8.6 | 7.0 | 33.8 | 25.6 | 6.8 | 1.9 | 11.1 | 4.2 | 18.3 | 10.3 |
| PA 7 × EQX 3338 | 7.9 | 6.9 | 35.6 | 31.1 | 4.6 | 2.5 | 7.0 | 3.4 | 15.4 | 7.9 |
| T63/967 × PA 118 | 7.1 | 6.1 | 29.6 | 26.6 | 5.8 | 1.5 | 9.3 | 3.0 | 15.5 | 7.4 |
| PA7 × PA 118 | 7.5 | 6.3 | 38.1 | 33.7 | 4.3 | 1.7 | 7.0 | 2.9 | 14.9 | 7.8 |
| T60/887 × CRG 8914 | 8.5 | 7.0 | 26.4 | 26.1 | 5.5 | 1.8 | 9.2 | 3.9 | 17.3 | 9.7 |
| T63/967 × PA 88 | 8.1 | 6.3 | 28.2 | 25.5 | 5.7 | 2.0 | 11.8 | 3.8 | 15.4 | 9.6 |
| SEDdf=78 | 0.28 | 0.25 | 4.51 | 3.28 | 0.51 | 0.24 | 0.79 | 0.28 | 0.80 | 0.54 |
| Muriate of Potash (g/plant) | Leaf Chlorophyll Content (mmol/cm2) | Leaf Chlorophyll Fluorescence (Fv/Fm) | Leaf Phenol Content (mg/g) | Leaf Proline Content (µg/g) | ||||
|---|---|---|---|---|---|---|---|---|
| Watered | Stressed | Watered | Stressed | Watered | Stressed | Watered | Stressed | |
| 0 | 11.1 | 5.2 | 0.67 | 0.15 | 10.1 | 16.8 | 9.9 | 1373.4 |
| 1 | 10.5 | 4.9 | 0.65 | 0.30 | 11.1 | 19.5 | 13.1 | 1397.4 |
| 2 | 9.4 | 4.2 | 0.63 | 0.19 | 11.8 | 19.6 | 13.2 | 1202.9 |
| 3 | 10.1 | 4.3 | 0.61 | 0.29 | 11.8 | 20.7 | 12.9 | 1317.5 |
| SEDdf=32 | 0.87 | 0.72 | 0.02 | 0.07 | 0.11 | 0.22 | 0.71 | 9.92 |
| Varieties | Leaf Chlorophyll Content (mmol/cm2) | Leaf Chlorophyll Fluorescence (Fv/Fm) | Relative Water Content (%) | Electrolyte Leakage (%) | Leaf Phenol Content (mg/g) | Leaf Proline Content (µg/g) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Watered | Stressed | Watered | Stressed | Watered | Stressed | Watered | Stressed | Watered | Stressed | Watered | Stressed | |
| T63/967 × SCA 9 | 12.8 | 5.6 | 0.61 | 0.16 | 75.4 | 13.0 | 9.9 | 36.1 | 11.6 | 20.2 | 13.4 | 996.2 |
| T63/971 × SCA 9 | 10.0 | 4.7 | 0.67 | 0.17 | 73.2 | 9.3 | 7.6 | 36.4 | 12.2 | 23.9 | 5.9 | 2108.4 |
| PA 7 × MAN 15-2 | 10.0 | 4.2 | 0.61 | 0.26 | 72.0 | 9.6 | 9.0 | 27.0 | 10.4 | 15.7 | 9.7 | 1092.8 |
| T85/799 × PA 7 | 8.4 | 4.1 | 0.66 | 0.33 | 72.6 | 7.7 | 8.7 | 16.7 | 10.6 | 16.8 | 20.1 | 1094.0 |
| SEDdf=32 | 0.87 | 0.72 | 0.02 | 0.07 | 2.67 | 0.88 | 0.55 | 1.65 | 0.11 | 0.22 | 0.71 | 9.92 |
| Hazard Ratios for Varieties (95% Confidence Intervals) | Hazard Ratios for K Levels (95% Confidence Intervals) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| PA 7 × MAN 15-2 | T63/967 × SCA 9 | T63/971 × SCA 9 | T85/799 × PA 7 | 0 g K | 1 g K | 2 g K | 3 g K | ||
| PA 7 × MAN 15-2 | - | 1.05 (0.52–2.54) | 1.23 (0.60–2.55) | 0.71 (0.36–1.41) | 0 g K | - | 0.79 (0.38–1.64) | 0.87 (0.41–1.83) | 0.47 (0.23–0.95) |
| T63/967 × SCA 9 | - | 1.17 (0.58–2.40) | 0.68 (0.34–1.33) | 1 g K | - | 1.10 (0.54–2.22) | 0.59 (0.31–1.15) | ||
| T63/971 × SCA 9 | - | 0.58 (0.29–1.15) | 2 g K | - | 0.54 (0.27–1.07) | ||||
| T85/799 × PA 7 | - | 3 g K | - | ||||||
| Log-rank test | χ2df=3 = 4.31; p = 0.23 | χ2df=3 = 8.27; p = 0.04 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anokye, E.; Lowor, S.T.; Dogbatse, J.A.; Padi, F.K. Potassium Application Positively Modulates Physiological Responses of Cocoa Seedlings to Drought Stress. Agronomy 2021, 11, 563. https://doi.org/10.3390/agronomy11030563
Anokye E, Lowor ST, Dogbatse JA, Padi FK. Potassium Application Positively Modulates Physiological Responses of Cocoa Seedlings to Drought Stress. Agronomy. 2021; 11(3):563. https://doi.org/10.3390/agronomy11030563
Chicago/Turabian StyleAnokye, Esther, Samuel T. Lowor, Jerome A. Dogbatse, and Francis K. Padi. 2021. "Potassium Application Positively Modulates Physiological Responses of Cocoa Seedlings to Drought Stress" Agronomy 11, no. 3: 563. https://doi.org/10.3390/agronomy11030563
APA StyleAnokye, E., Lowor, S. T., Dogbatse, J. A., & Padi, F. K. (2021). Potassium Application Positively Modulates Physiological Responses of Cocoa Seedlings to Drought Stress. Agronomy, 11(3), 563. https://doi.org/10.3390/agronomy11030563