
Effects of Light Orientation and Mechanical Damage to Leaves on Isoflavone Accumulation in Soybean Seeds



Abstract
:1. Introduction
2. Materials and Methods
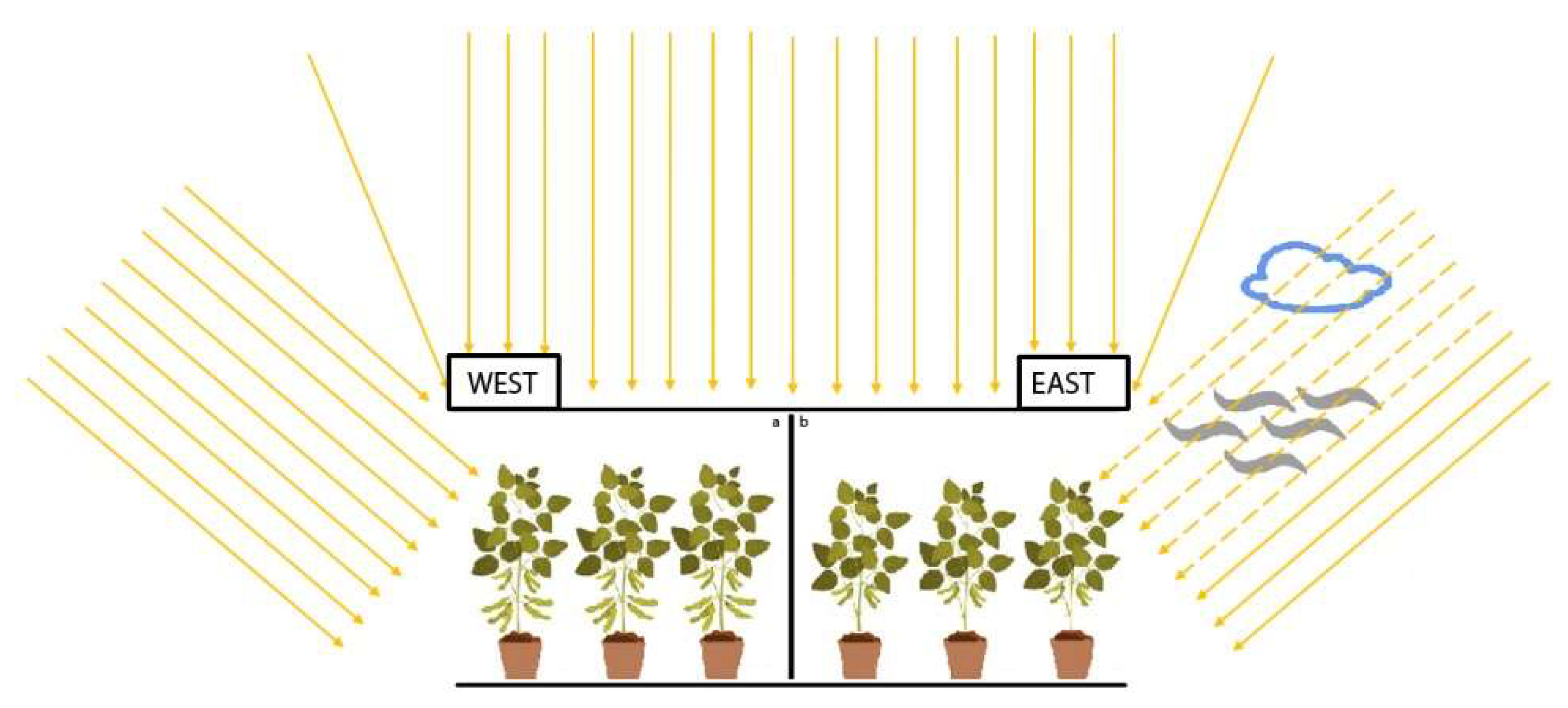
2.1. Experimental Set-Up
2.2. Plant Analysis
2.3. Statistical Analysis
3. Results
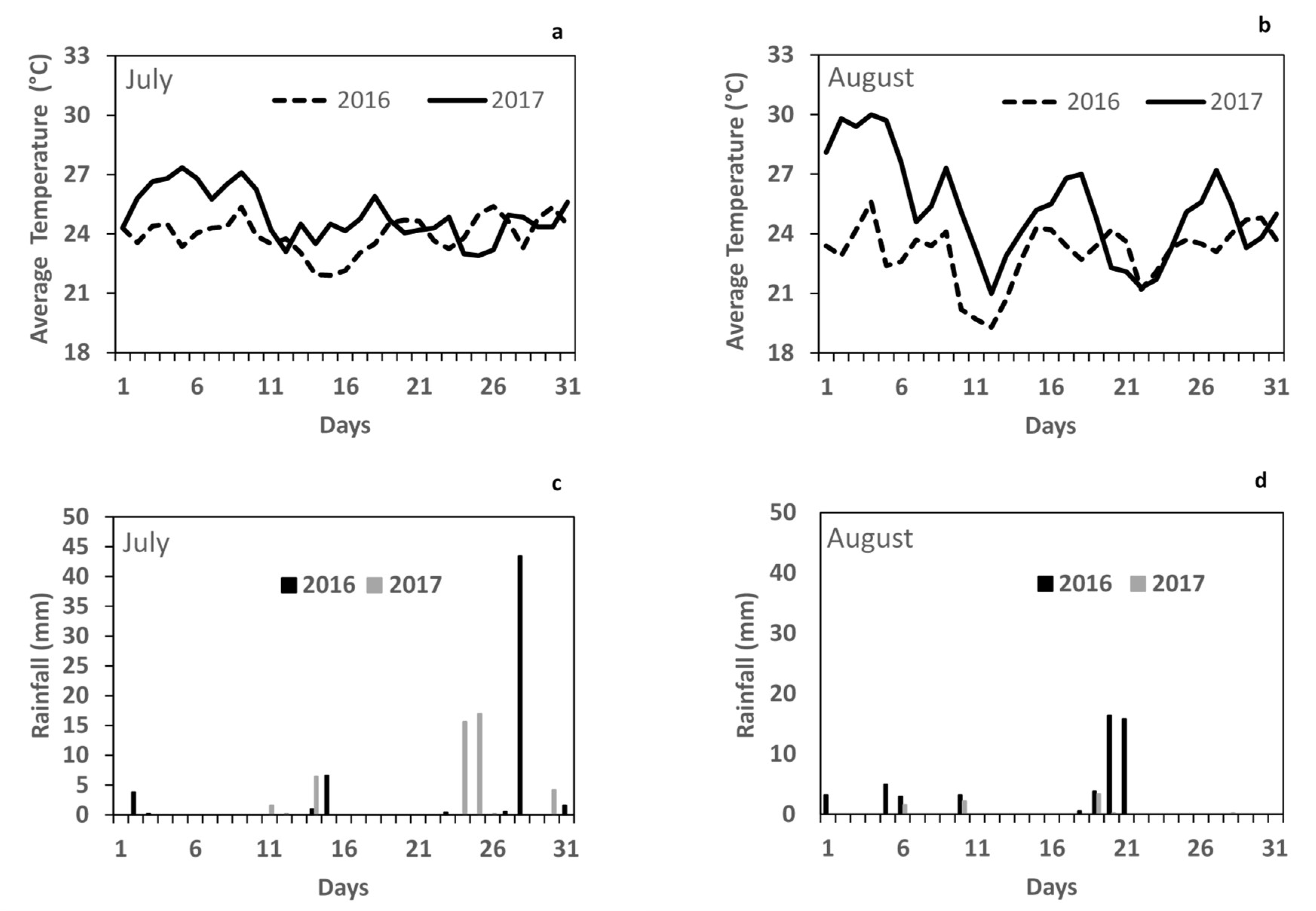
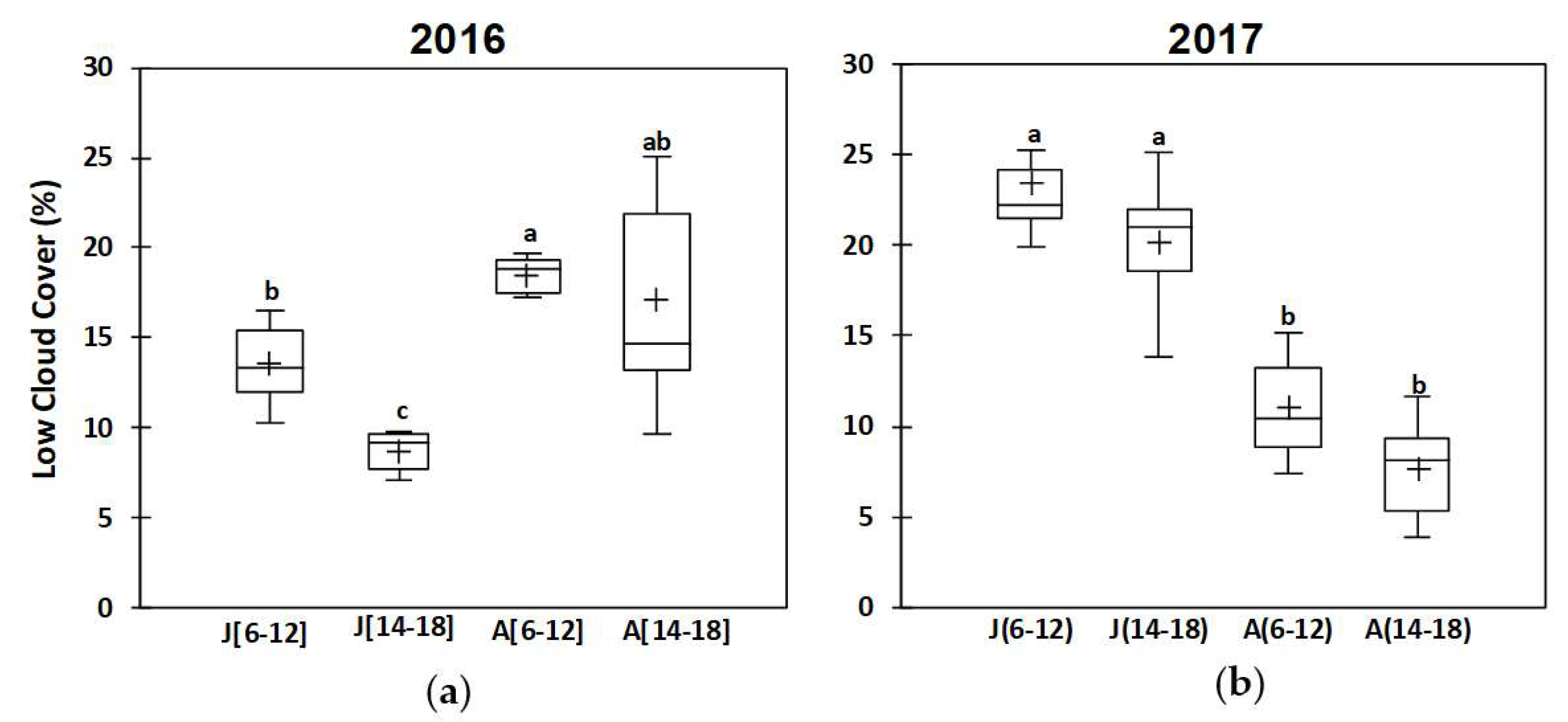
3.1. Climate Parameters
3.2. Plant Biomass and Yield
3.3. Quality Parameters
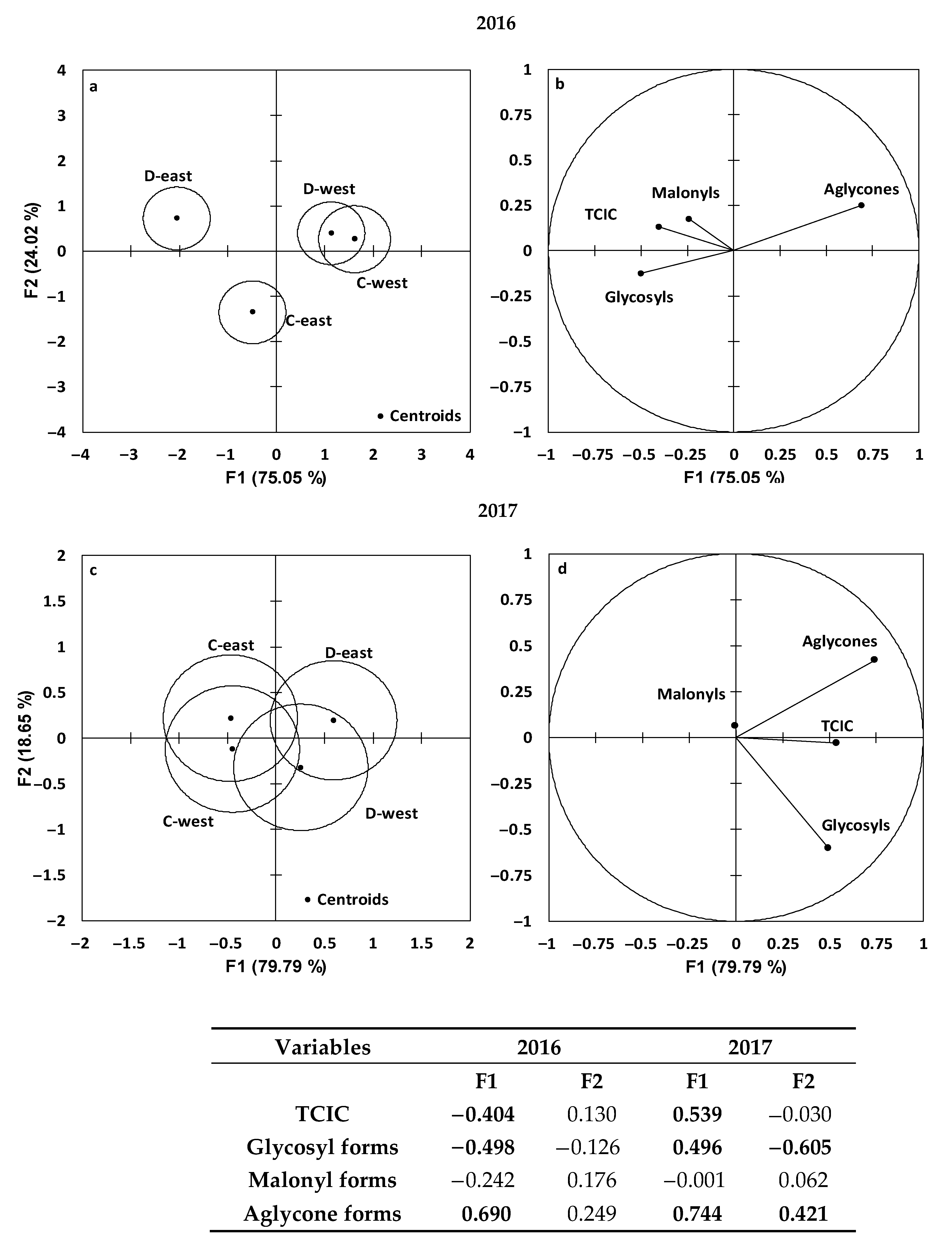
3.4. PCA and MDA on Isoflavone Concentration and Profile within Treatments
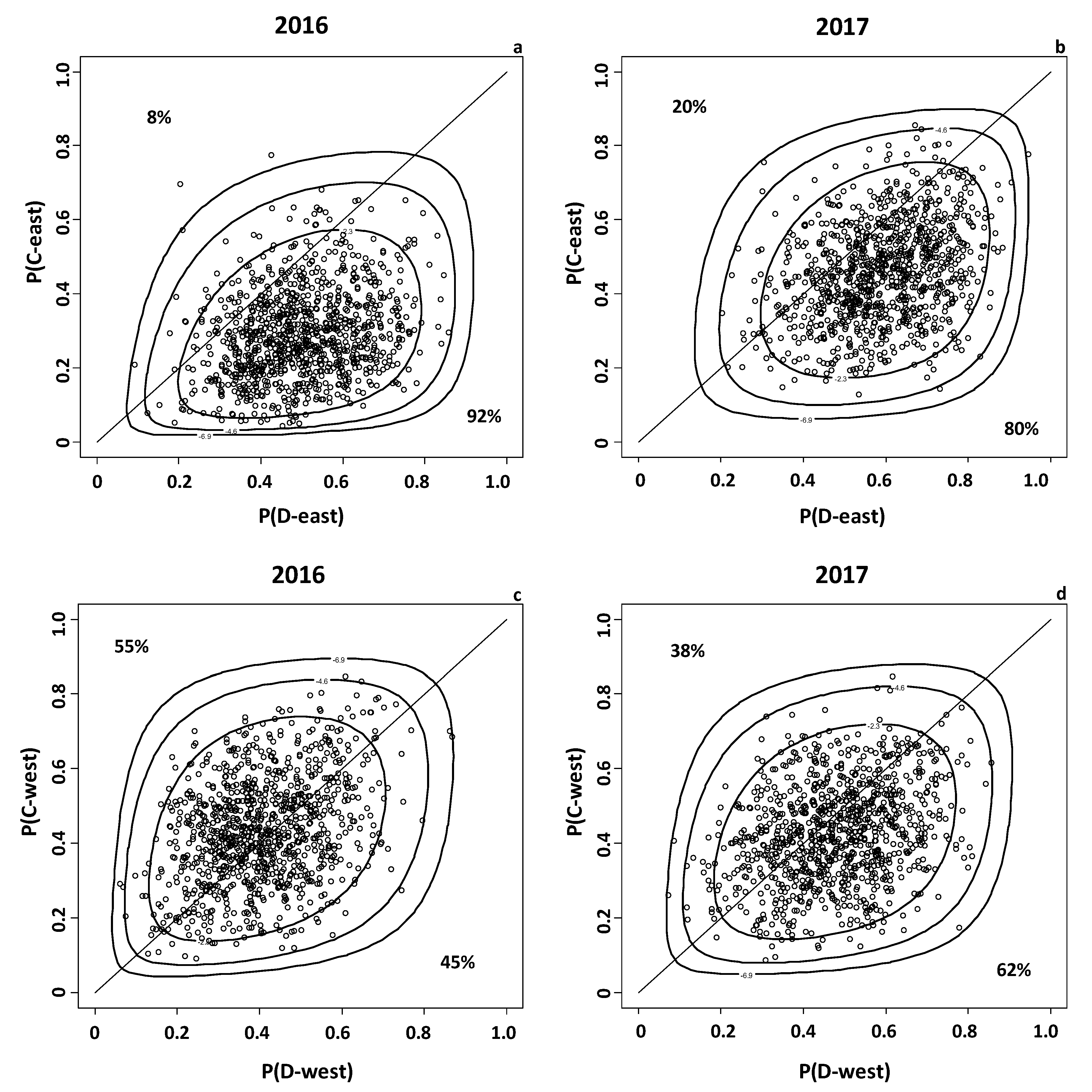
3.5. Monte Carlo Analysis on the Probability to Maximize TCIC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sarkar, F.H.; Li, Y. Soy isoflavones and cancer prevention: Clinical science review. Cancer Investig. 2003, 21, 744–757. [Google Scholar] [CrossRef] [PubMed]
- Sathyapalan, T.; Aye, M.; Rigby, A.; Thatcher, N.; Dargham, S.; Kilpatrick, E.; Atkin, S. Soy isoflavones improve cardiovascular disease risk markers in women during the early menopause. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 691–697. [Google Scholar] [CrossRef]
- Barion, G.; Mosca, G.; Vamerali, T. Estimation of cotyledon isoflavone abundance by a grey luminance-based model in variously hilum-coloured soybean varieties. J. Sci. Food Agric. 2016, 96, 4126–4134. [Google Scholar] [CrossRef]
- Lee, C.H.; Yang, L.; Xu, J.Z.; Yeung, S.Y.V.; Huang, Y.; Chen, Z.Y. Relative antioxidant activity of soybean isoflavones and their glycosides. Food Chem. 2005, 90, 735–741. [Google Scholar] [CrossRef]
- Arai, Y.; Uehara, M.; Sato, Y.; Kimira, M.; Eboshida, A.; Adlercreutz, H.; Watanabe, S. Comparison of isoflavones among dietary intake, plasma concentration and urinary excretion for accurate estimation of phytoestrogen intake. J. Epidemiol. 2000, 10, 127–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skalicky, M.; Kubes, J.; Hejnak, V.; Tumova, L.; Martinkova, J.; Martin, J.; Hnilickova, H. Isoflavones production and possible mechanism of their exudation in Genista tinctoria L. suspension culture after treatment with vanadium compounds. Molecules 2018, 23, 1619. [Google Scholar] [CrossRef] [Green Version]
- Rasolohery, C.A.; Berger, M.; Lygin, A.V.; Lozovaya, V.V.; Nelson, R.L.; Daydé, J. Effect of temperature and water availability during late maturation of the soybean seed on germ and cotyledon isoflavone content and composition. J. Sci. Food Agric. 2008, 88, 218–228. [Google Scholar] [CrossRef]
- Zhu, H.; Li, X.; Zhai, W.; Liu, Y.; Gao, Q.; Liu, J.; Zhu, Y. Effects of low light on photosynthetic properties, antioxidant enzyme activity, and anthocyanin accumulation in purple pak-choi (Brassica campestris ssp. Chinensis Makino). PLoS ONE 2017, 12, e0179305. [Google Scholar] [CrossRef]
- Mosquera-Losada, M.R.; Santiago-Freijanes, J.J.; Rois-Díaz, M.; Moreno, G.; Den Herder, M.; Aldrey-Vázquez, J.A.; Rigueiro-Rodríguez, A. Agroforestry in Europe: A land management policy tool to combat climate change. Land Use Policy 2018, 78, 603–613. [Google Scholar] [CrossRef]
- Rosenstock, T.S.; Wilkes, A.; Jallo, C.; Namoi, N.; Bulusu, M.; Suber, M.; Mboi, D.; Mulia, R.; Simelton, E.; Richards, M. Making trees count: Measurement and reporting of agroforestry in UNFCC national communications of non-annex countries. Agric. Ecosyst. Environ. 2019, 284, 106569. [Google Scholar] [CrossRef]
- Jose, S. Agroforestry for ecosystem services and environmental benefits: An overview. Agrofor. Syst. 2009, 76, 1–10. [Google Scholar] [CrossRef]
- Dupraz, C.; Blitz-Frayret, C.; Lecomte, I.; Molto, Q.; Reyes, F.; Gosme, M. Influence of latitude on the light availability for intercrops in an agroforestry alley-cropping system. Agrofor. Syst. 2018, 92, 1019–1033. [Google Scholar] [CrossRef]
- Cano-Cruz, E.M.; López-Orozco, F. Design and development of a low-cost and portable meteorological system: Meteoblue. Adv. Sci. Lett. 2015, 21, 83–87. [Google Scholar] [CrossRef]
- Barion, G.; Hewidy, M.; Mosca, G.; Vamerali, T. Intraspecific variability for soybean cotyledon isoflavones in different cropping and soil conditions. Eur. J. Agron. 2010, 33, 63–73. [Google Scholar] [CrossRef]
- Hubert, J.; Berger, M.; Daydé, J. Use of a simplified hplc-UV analysis for soyasaponin b determination: Study of saponin and isoflavone variability in soybean cultivars and soy-based health food products. J. Agric. Food Chem. 2005, 53, 3923–3930. [Google Scholar] [CrossRef] [PubMed]
- Podani, J. PCA. Ordinamento. In Analisi ed Esplorazione Multivariate dei Dati in Ecologia e Biologia; Liguori Editore: Napoli, Italy, 2007; pp. 333–334. [Google Scholar]
- Ihaka, R.; Gentleman, R. R: A language for data analysis and graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- Albert, J. Multiparameter models. In Bayesian Computation with R; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009; pp. 76–79. [Google Scholar]
- Howard, J. The 2 × 2 table: A discussion from a bayesian viewpoint. Stat. Sci. 1998, 351–367. [Google Scholar]
- Song, Y.H.; Kubota, A.; Kwon, M.S.; Covington, M.F.; Lee, N.; Taagen, E.R.; Laboy Cintrón, D.; Hwang, D.Y.; Akiyama, R.; Hodge, S.K.; et al. Molecular basis of flowering under natural long-day conditions in Arabidopsis. Nat. Plants 2018, 4, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Beć, K.B.; Huck, C.W. Breakthrough potential in near-infrared spectroscopy: Spectra simulation. A review of recent developments. Front. Chem. 2019, 7, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorrain, S.; Allen, T.; Duek, P.D.; Whitelam, G.C.; Fankhauser, C. Phytochrome-mediated inhibition of shade avoidance involves degradation of growth-promoting bhlh transcription factors. Plant J. 2008, 53, 312–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Il fitocromo e lo sviluppo delle piante controllato dalla luce. In Fisiologia Vegetale; Piccin Nuova Libraria S.p.A.: Padova, Italy, 2013; pp. 713–714. [Google Scholar]
- Xu, X.; Kathare, P.K.; Pham, V.N.; Bu, Q.; Nguyen, A.; Huq, E. Reciprocal proteasome-mediated degradation of pifs and hfr1 underlies photomorphogenic development in Arabidopsis. Development 2017, 144, 1831–1840. [Google Scholar] [CrossRef] [Green Version]
- Dittes, L.; Rissland, I.; Mohr, H. On the regulation of enzyme levels (phenylalanine ammonia-lyase) in different organs of a plant (sinapis alba l.). Z. Nat. B 1971, 26, 1175–1180. [Google Scholar] [CrossRef]
- Beggs, C.J.; Kuhn, K.; Böcker, R.; Wellmann, E. Phytochrome-induced flavonoid biosynthesis in mustard (sinapis alba L.) cotyledons. enzymic control and differential regulation of anthocyanin and quercetin formation. Planta 1987, 172, 121–126. [Google Scholar] [CrossRef]
- Brödenfeldt, R.; Mohr, H. Time courses for phytochrome-induced enzyme levels in phenylpropanoid metabolism (phenylalanine ammonia-lyase, naringenin-chalcone synthase) compared with time courses for phytochrome-mediated end-product accumulation (anthocyanin, quercetin). Planta 1988, 176, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Pham, V.N.; Kathare, P.K.; Huq, E. Phytochromes and phytochrome interacting factors. Plant Physiol. 2018, 176, 1025–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, H.; Matsuoka, M.; Steber, C.M. A role for the ubiquitin–26S-proteasome pathway in gibberellin signaling. Trends Plant Sci. 2003, 8, 492–497. [Google Scholar] [CrossRef]
- Kasperbauer, M.J. Far-red light reflection from green leaves and effects on phytochrome- mediated assimilate partitioning under field conditions. Plant Physiol. 1987, 85, 350–354. [Google Scholar] [CrossRef] [Green Version]
- Van Gelderen, K.; Kang, C.; Paalman, R.; Keuskamp, D.; Hayes, S.; Pierik, R. Far-red light detection in the shoot regulates lateral root development through the hy5 transcription factor. Plant Cell 2018, 30, 101–116. [Google Scholar] [CrossRef] [Green Version]
- Hitz, T.; Hartung, J.; Graeff-Hönninger, S.; Munz, S. Morphological response of soybean (glycine max (L.) merr.) cultivars to light intensity and red to far-red ratio. Agron 2019, 9, 428. [Google Scholar] [CrossRef] [Green Version]
- Vamerali, T.; Barion, G.; Hewidy, M.; Mosca, G. Soybean isoflavone patterns in main stem and branches as affected by water and nitrogen supply. Eur. J. Agron. 2012, 41, 1–10. [Google Scholar] [CrossRef]
- Reynolds, P.E.; Simpson, J.A.; Thevathasan, N.V.; Gordon, A.M. Effects of tree competition on corn and soybean photosynthesis, growth, and yield in a temperate tree-based agroforestry intercropping system in southern Ontario, Canada. Ecol. Eng. 2007, 29, 362–371. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, Y.; Cai, J.; Jiang, Z.; Zhang, S. Photosynthesis, growth and yield of soybean and maize in a tree-based agroforestry intercropping system on the Loess Plateau. Agrofor. Syst. 2009, 76, 569–577. [Google Scholar] [CrossRef]
- Mantino, A.; Volpi, I.; Micci, M.; Pecchioni, G.; Bosco, S.; Dragoni, F.; Ragaglini, G. Effect of tree presence and soil characteristics on soybean yield and quality in an innovative alley-cropping system. Agronomy 2020, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Carrier, M.; Gonzalez, F.A.R.; Cogliastro, A.; Olivier, A.; Vanasse, A.; Rivest, D. Light availability, weed cover and crop yields in second generation of temperate tree-based intercropping systems. Field Crops Res. 2019, 239, 30–37. [Google Scholar] [CrossRef]
- Ceron-Garcia, A.; Vargas-Arispuro, I.; Aispuro-Hernandez, E.; Martinez-Tellez, M.A. Oligoglucan elicitor effects during plant oxidative stress. In Cell Metabolism. Cell Homeostasis and Stress Response; Bubulya, P., Ed.; IntechOpen: Apodaca, México, 2012. [Google Scholar]
- Delledonne, M.; Zeier, J.; Marocco, A.; Lamb, C. Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response. Proc. Natl. Acad. Sci. USA 2001, 98, 13454–13459. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | 2016 | 2017 | ||
|---|---|---|---|---|
| West | East | West | East | |
| Shoot biomass (g) | ||||
| Foliar damage (D) | 71.7 a | 58.9 ab | 68.5 a | 56.3 ab |
| Control (C) | 59.6 ab | 42.1 b | 55.1 a | 45.8 b |
| Grain yield (g plant−1) | ||||
| Foliar damage (D) | 39.5 a | 32.1 ab | 25.6 a | 19.7 a |
| Control (C) | 32.4 ab | 22.1 b | 16.7 b | 15.3 b |
| Nb of pods/plant | ||||
| Foliar damage (D) | 84.7 a | 69.9 a | 66.2 a | 55.2 a |
| Control (C) | 57.7 ab | 43.3 b | 58.1 a | 34.4 b |
| Cotyledon weight (g) | ||||
| Foliar damage (D) | 0.139 a | 0.136 a | 0.225 a | 0.279 a |
| Control (C) | 0.138 a | 0.119 a | 0.223 a | 0.183 a |
| Protein (%) | ||||
| Foliar damage (D) | 43.4 a | 43.6 a | 43.8 a | 44.0 a |
| Control (C) | 43.6 a | 43.9 a | 44.1 a | 44.2 a |
| TCIC (mg g−1) | ||||
| Foliar damage (D) | 2.60 b | 3.12 a | 1.08 b | 1.38 a |
| Control(C) | 2.49 b | 2.51 b | 1.06 b | 1.22 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barion, G.; Hewidy, M.; Panozzo, A.; Aloia, A.; Vamerali, T. Effects of Light Orientation and Mechanical Damage to Leaves on Isoflavone Accumulation in Soybean Seeds. Agronomy 2021, 11, 589. https://doi.org/10.3390/agronomy11030589
Barion G, Hewidy M, Panozzo A, Aloia A, Vamerali T. Effects of Light Orientation and Mechanical Damage to Leaves on Isoflavone Accumulation in Soybean Seeds. Agronomy. 2021; 11(3):589. https://doi.org/10.3390/agronomy11030589
Chicago/Turabian StyleBarion, Giuseppe, Mohamed Hewidy, Anna Panozzo, Andrea Aloia, and Teofilo Vamerali. 2021. "Effects of Light Orientation and Mechanical Damage to Leaves on Isoflavone Accumulation in Soybean Seeds" Agronomy 11, no. 3: 589. https://doi.org/10.3390/agronomy11030589
APA StyleBarion, G., Hewidy, M., Panozzo, A., Aloia, A., & Vamerali, T. (2021). Effects of Light Orientation and Mechanical Damage to Leaves on Isoflavone Accumulation in Soybean Seeds. Agronomy, 11(3), 589. https://doi.org/10.3390/agronomy11030589