
Morphophysiological, Enzymatic, and Elemental Activity in Greenhouse Tomato Saladette Seedlings from the Effect of Plant Growth-Promoting Rhizobacteria


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Inoculation in Seed and Production of Seedlings
2.2. Irrigation and Preparation of Nutrient Solution
2.3. Variables Evaluated In Vivo in Greenhouse Tomato Seedlings
2.3.1. Seedling Height; Stem Diameter; Root Displaced Volume; Fresh and Dry Weight (Dry Matter) of Leaves, Stem, and Root; Leaf Area
2.3.2. Enzymatic Activity
2.3.3. Plant Tissue Elemental Analysis
2.3.4. Microbiological Analysis
2.4. Experimental Design
3. Results
3.1. Morphophysiological Responses, Enzymatic Activity, and Mineral–Elemental Response in Seedlings, Due to the Inoculation of PGPR Associated with Distichlis Spicata
3.1.1. Seedling Height (PTH)
3.1.2. Stem Diameter (SDM)
3.1.3. Root Displaced Volume (RDV)
3.1.4. Leaf Area (LA)
3.1.5. Plant Fresh Weight and Plant Dry Matter
3.1.6. Enzymatic Activity—Nitrate Reductase Analysis: (1) Endogenous Reaction (ENDO), (2) Induced Reaction with NO3− Substrate (NSB), (3) Reaction with Mo as Cofactor (CMO), and (4) Substrate and Cofactor NO3− + Mo (MNR)
3.1.7. Plant Tissue Elemental Analysis
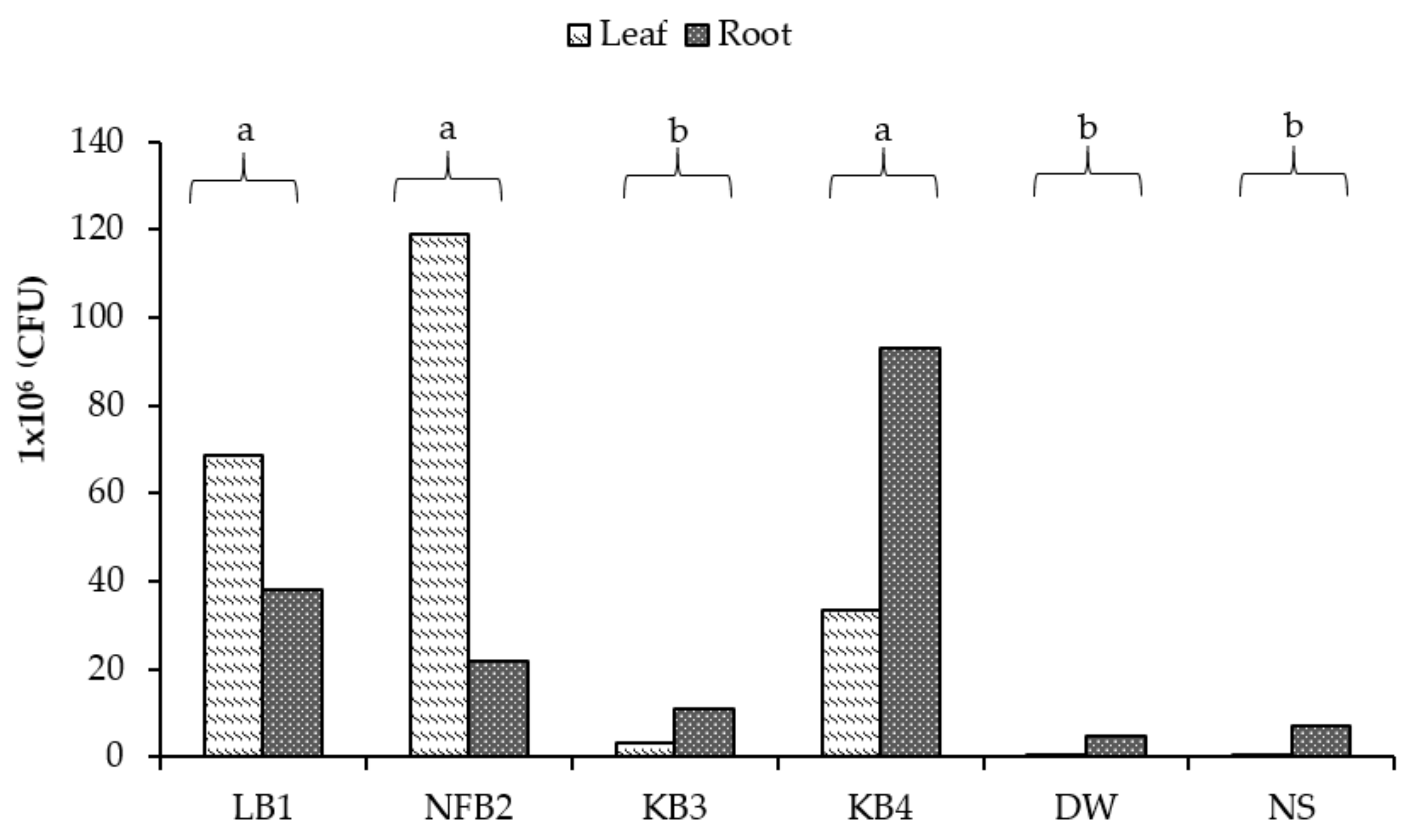
3.1.8. Microbiological Analysis (CFU)
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT, Food and Agriculture Organization Corporate Statistical Database. Available online: http://www.fao.org/faostat/es/#data/QC (accessed on 9 January 2021).
- SIAP-SAGARPA. Servicio de Información Agroalimentaria y Pesquera—Secretaria de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. Available online: http://infosiap.siap.gob.mx/aagricola_siap_gb/ientidad/index.jsp (accessed on 25 January 2021).
- Diver, S.; Greer, L. Sustainable Small-Scale Nursery Production; National Center for Appropriate Technology: Butte, MT, USA, 2008; p. 28. Available online: http://www.microfarms.com/technical/greenhousecd/greenhouse/nursery/nursery.pdf (accessed on 10 March 2021).
- Grageda-Cabrera, O.A.; Díaz-Franco, A.; Peña-Cabriales, J.J.; Vera-Nuñez, J.A. Impacto de los biofertilizantes en la agricultura. Rev. Mex. Cienc. Agríc. 2012, 3, 1261–1274. [Google Scholar] [CrossRef] [Green Version]
- Escobar, C.; Horna, Y.; Carreño, C.; Mendoza, G. Caracterización de cepas nativas de Azotobacter spp. y su efecto en el desarrollo de Lycopersicon esculentum Mill. “tomate” en Lambayeque. Sci. Agropecu. 2011, 2, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Armenta-Bojórquez, A.D.; García-Gutiérrez, C.; Camacho-Báez, J.R.; Apodaca-Sánchez, M.Á.; Gerardo-Montoya, L.; Nava-Pérez, E. Biofertilizantes en el desarrollo agrícola de México. Ra Ximhai 2010, 6, 51–56. [Google Scholar] [CrossRef]
- Lara, C.; Avila, L.; Peñata, J. Native phosphate solubilizing bacteria to increase the crops in the department of Cordova-Colombia. Biotecnol. Sect. Agropecu. Agroind. 2011, 9, 114–120. [Google Scholar]
- Afzal, M.; Yousaf, S.; Reichenauer, T.G.; Sessitsch, A. The Inoculation Method Affects Colonization and Performance of Bacterial Inoculant Strains in the Phytoremediation of Soil Contaminated with Diesel Oil. Int. J. Phytoremediat. 2012, 14, 35–47. [Google Scholar] [CrossRef]
- Jasso, C.C.; Martínez, G.M.A.; Chávez, V.J.R.; Ramírez, T.J.A.; Garza, E. Guía Para Cultivar Jitomate en Condiciones de Malla Sombra en San Luís Potosí; Folleto técnico: SAGARPA, Mexico. 2012, pp. 9–54. Available online: http://www.inifapcirne.gob.mx/Biblioteca/Publicaciones/905.pdf (accessed on 12 January 2021).
- Carrillo, A.; Puente, M.; Castellanos, T.; Bashan, Y. Aplicaciones Biotecnologicas de Ecología Microbiana. Manual de Laboratorio Pontificia Universidad Javeriana; Centro de Investigaciones Biológicas del Noroeste: La Paz, Baja California Sur, Mexico, 1998; p. 51. [Google Scholar]
- Palacio-Rodríguez, R.; Coria-Arellano, J.L.; López-Bucio, J.; Sánchez-Salas, J.; Muro-Pérez, G.; Castañeda-Gaytán, G.; Sáenz-Mata, J. Halophilic rhizobacteria from Distichlis spicata promote growth and improve salt tolerance in heterologous plant hosts. Symbiosis 2017, 73, 179–189. [Google Scholar] [CrossRef]
- Richards, L.A. Diagnosis and Improvement of Saline and Alkali Soils; U.S. Department Agriculture Handbook 60: Washington, DC, USA, 1954. [Google Scholar]
- Santamaría-César, J.; Figueroa-Viramontes, U.; Medina-Morales, M.d.C. Productividad de la alfalfa en condiciones de salinidad en el Distrito de Riego 017, Comarca Lagunera. Terra Latinoam. 2004, 22, 343–349. [Google Scholar]
- Jaworski, E.G. Nitrate reductase assay in intact plant tissues. Biochem. Biophys. Res. Commun. 1971, 43, 1274–1279. [Google Scholar] [CrossRef]
- Mauriño, S.G.; Echevarria, C.; Mejias, J.A.; Vargas, M.A.; Maldonado, J.M. Properties of the in vivo Nitrate Reductase Assay in Maize, Soybean, and Spinach Leaves. J. Plant Physiol. 1986, 124, 123–130. [Google Scholar] [CrossRef]
- Hageman, R.H.; Hucklesby, D.P. Nitrate reductase from higher plants. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1971; Volume 23, pp. 491–503. [Google Scholar]
- Fernández, V.; Del Río, V.; Abadía, J.; Abadía, A. Foliar Iron Fertilization of Peach (Prunus persica L.) Batsch: Effects of Iron Compounds, Surfactants and Other Adjuvants. Plant Soil 2006, 289, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Wacal, C.; Ogata, N.; Basalirwa, D.; Handa, T.; Sasagawa, D.; Acidri, R.; Ishigaki, T.; Kato, M.; Masunaga, T.; Yamamoto, S.; et al. Growth, Seed Yield, Mineral Nutrients and Soil Properties of Sesame (Sesamum indicum L.) as Influenced by Biochar Addition on Upland Field Converted from Paddy. Agronomy 2019, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Helrich, K. Official Methods of Analysis of the Association of Official Analytical Chemists, 15th ed.; Arlington, VA Association of Official Analytical Chemists United States: Arlington, VA, USA, 1990; pp. 40–49. [Google Scholar]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SAS (Statistical Analysis System). SAS/STAT 9.2 User’s Guide; SAS: Cary, NC, USA, 2016; Available online: https://support.sas.com/documentation/cdl/en/statugintroduction/61750/PDF/default/statugintroduction.pdf (accessed on 10 January 2021).
- Njira, K. A review of effects of nutrient elements on crop quality. African Journal of Food, Agriculture. Nutr. Dev. 2015, 15, 9777–9783. [Google Scholar]
- Puente, M.E.; Holguin, G.; Glick, B.R.; Bashan, Y. Root-surface colonization of black mangrove seedlings by Azospirillum halopraeferens and Azospirillum brasilense in seawater. FEMS Microbiol. Ecol. 1999, 29, 283–292. [Google Scholar] [CrossRef]
- Rozema, J. The influence of salinity, inundation and temperature on the germination of some halophytes and non-halophytes. Oecol. Plant 1995, 10, 341. [Google Scholar]
- Goodfriend, W.L.; Olsen, M.W.; Frye, R.J. Soil microfloral and microfaunal response to Salicornia bigelovii planting density and soil residue amendment. Plant Soil 2000, 223, 23–32. [Google Scholar] [CrossRef]
- Díaz-Vargas, P.; Ferrera-Cerrato, R.; Almaraz-Suárez, J.J.; Alcántar-González, G. Inoculación de bacterias promotoras de crecimiento en lechuga. Terra Latinoam. 2001, 19, 327–335. [Google Scholar]
- Arsac, J.F.; Lamothe, C.; Mulard, D.; Fages, J. Growth enhancement of maize (Zea mays L.) through Azospirillum lipoferum inoculation: Effect of plant genotype and bacterial concentration. Agronomie 1990, 10, 640–654. [Google Scholar] [CrossRef]
- Puente, M.; Bashan, Y.J.S. Effect of Inoculation with Azospirillum brasilense Strains on the Germination and Seedlings Growth of the Giant Columnar Cardon Cactus (Pachycereus pringlei). Symbiosis 1993, 15, 49–60. [Google Scholar]
- Haahtela, K.; Ronkko, R.; Laaskso, T.; Williams, P.; Korhonen, T. Root-associated Enterobacter and Klebsiella in Poa pratensis: Characterization of an iron-scavening system and a subtance stimulating root hair production. Mol. Plant Microbe Interact. 1990, 3, 358–365. [Google Scholar] [CrossRef]
- Turyanitsa, A.; Petak, A.; Nichik, M.; Petrosova, V. Capacity of Klebsiella bacteria to fix atmospheric nitrogen and to produce plant growth hormones. Mikrobiol 1995, 57, 28–34. [Google Scholar]
- El-Khawas, H.; Adachi, K. Identification and quantification of auxins in culture media of Azospirillum and Klebsiella and their effect on rice roots. Biol. Fertil. Soils 1999, 28, 377–381. [Google Scholar] [CrossRef]
- Zulueta, R.; Hernandez-Montiel, L.G.; Amador, B.; Rueda-Puente, E.; Capistrán, L.; Guez, E.; Cordoba, M.; Yu, F.-H.; Jokela, E. Effect of Hydropriming and Biopriming on Seed Germination and Growth of Two Mexican Fir Tree Species in Danger of Extinction. Forests 2015, 6, 3109–3122. [Google Scholar] [CrossRef] [Green Version]
- Noh Medina, J.; Yam Chimal, C.; Borges Gómez, L.; Zúñiga Aguilar, J.J.; Godoy Hernández, G. Aislados bacterianos con potencial biofertilizante para plántulas de tomate. Terra Latinoam. 2014, 32, 273–281. [Google Scholar]
- Aguado-Santacruz, G.; Moreno-Gómez, B.; Jimenez, B.; Moya, E.; Preciado, E. Impact of the microbial siderophores and phytosiderophores on the iron assimilation by plants: A synthesis. Rev. Fitotec. Mex. 2012, 35, 9–21. [Google Scholar]
- Cisneros-Rojas, C.A.; Sánchez-de Prager, M.; Menjivar-Flores, J.C. Efecto de bacterias solubilizadoras de fosfatos sobre el desarrollo de plántulas de café. Agron. Mesoam. 2017, 28, 149–158. [Google Scholar] [CrossRef] [Green Version]
- El-Shatnawi, M.K.J.; Makhadmeh, I.M. Ecophysiology of the Plant–Rhizosphere System. J. Agron. Crop Sci. 2001, 187, 1–9. [Google Scholar] [CrossRef]
- Espinosa Palomeque, B.; Moreno Reséndez, A.; Cano Ríos, P.; Álvarez Reyna, V.d.P.; Sáenz Mata, J.; Sánchez Galván, H.; González Rodríguez, G. Inoculación de rizobacterias promotoras del crecimiento vegetal en tomate (Solanum lycopersicum L.) cv. afrodita en invernadero. Terra Latinoam. 2017, 35, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Rojas-Solís, D.; Hernández-Pacheco, C.E.; Santoyo, G. Evaluation of Bacillus and Pseudomonas to colonize the rhizosphere and their effect on growth promotion in tomato (Physalis ixocarpa Brot. ex Horm.). Revista Chapingo. Ser. Hortic. 2016, 22, 45–58. [Google Scholar]
- Geisseler, D.; Horwath, W.R.; Joergensen, R.G.; Ludwig, B. Pathways of nitrogen utilization by soil microorganisms—A review. Soil Biol. Biochem. 2010, 42, 2058–2067. [Google Scholar] [CrossRef]
- Méndez-Gómez, M.; Castro-Mercado, E.; García-Pineda, E. Azospirillum una rizobacteria con uso potencial en la agricultura. Biológicas 2014, 16, 11–18. [Google Scholar]
- Coppens, J.; Grunert, O.; van den Hende, S.; Vanhoutte, I.; Boon, N.; Haesaert, G.; Gelder, L. The use of microalgae as a high-value organic slow-release fertilizer results in tomatoes with increased carotenoid and sugar levels. J. Appl. Phycol. 2016, 28. [Google Scholar] [CrossRef]
- Sánchez-López, D.B.; Gómez-Vargas, R.M.; Garrido Rubiano, M.F.; Bonilla Buitrago, R.R. Inoculación con bacterias promotoras de crecimiento vegetal en tomate bajo condiciones de invernadero. Rev. Mex. Cienc. Agríc. 2012, 3, 1401–1415. [Google Scholar]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Zahir, Z.A.; Arshad, M.; Frankenberger, W.T. Plant Growth Promoting Rhizobacteria: Applications and Perspectives in Agriculture. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2003; Volume 81, pp. 97–168. [Google Scholar]
- Esquivel-Cote, R.; Ramírez-Gama, R.M.; Tsuzuki-Reyes, G.; Orozco-Segovia, A.; Huante, P. Azospirillum lipoferum strain AZm5 containing 1-aminocyclopropane-1-carboxylic acid deaminase improves early growth of tomato seedlings under nitrogen deficiency. Plant Soil 2010, 337, 65–75. [Google Scholar] [CrossRef]
- Ribaudo, C.M. Mecanismos Bioquímicos y Moleculares Desencadenados en la Interacción Bacterias Promotoras de Crecimiento Vegetal y Plantas de Interés Agronómico. Ph.D. Thesis, Universidad de Buenos Aires, Facultad de Ciencias Exactas y Naturales, Buenos Aires, Argentina, 2003. Available online: http://hdl.handle.net/20.500.12110/tesis_n5372_Ribaudo (accessed on 17 April 2021).
- Halvin, J.L.; Beaton, J.; Tisdale, S.; Nelson, W. Soil fertility and fertilizers: An Introduction to Nutrient Management; Pearson: Upper Saddle River, NJ, USA, 2005. [Google Scholar]
- Fukami, J.; Nogueira, M.; Araujo, R.; Hungria, M. Accessing inoculation methods of maize and wheat with Azospirillum brasilense. AMB Express 2016, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Hnilickova, H.; Hnilička, F.; Orsák, M.; Hejnak, V. Effect of salt stress on growth, electrolyte leakage, Na+ and K+ content in selected plant species. Plant Soil Environ. 2019, 65. [Google Scholar] [CrossRef] [Green Version]
- Shao, J.F.; Yamaji, N.; Shen, R.F.; Ma, J.F. The Key to Mn Homeostasis in Plants: Regulation of Mn Transporters. Trends Plant Sci. 2017, 22, 215–224. [Google Scholar] [CrossRef]
- Brown, P.H.; Welch, R.M.; Cary, E.E.; Checkai, R.T. Micronutrients. J. Plant Nutr. 1987, 10, 2125–2135. [Google Scholar] [CrossRef]
- Rodríguez-Jiménez, T.; Ojeda-Barrios, D.; Blanco-Macías, F.; Valdez-Cepeda, R.; Parra-Quezada, R. Ureasa y níquel en la fisiología de las plantas. Rev. Chapingo Ser. Hortic. 2016, 22, 69–81. [Google Scholar] [CrossRef]
- Liu, D.-Y.; Liu, Y.-M.; Zhang, W.; Chen, X.-P.; Zou, C.-Q. Zinc Uptake, Translocation, and Remobilization in Winter Wheat as Affected by Soil Application of Zn Fertilizer. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Muthukumararaja, T.M.; Sriramachandrasekharan, M.V. Effect of zinc on yield, zinc nutrition and zinc use efficiency of lowland rice. Int. J. Agric. Technol. 2012, 8, 551–561. [Google Scholar]
- Peña, H.; Medina-Hernandez, D.; Ghasemi, M.; Rueda-Puente, E. Salt tolerant plants as a valuable resource for sustainable food production in arid and saline coastal zones. Acta Biol. Colomb. 2020, 26. [Google Scholar] [CrossRef]
- Fages, J. An industrial view of Azospirillum inoculants: Formulation and application technology. Symbiosis 1992, 13, 15–26. [Google Scholar]


{kind=link}
| Treatments | Bacteria 1 × 108 CFU | Mode of Application | |
|---|---|---|---|
| 1 | B. paralicheniformis | LB1 | Seed Inoculation (IS) |
| 2 | B. paralicheniformis | LB1 | Seedling Spray (AP) |
| 3 | B. paralicheniformis | LB1 | Both (AP + IS) |
| 4 | A. guillouiae | NFB2 | Seed Inoculation (IS) |
| 5 | A. guillouiae | NFB2 | Seedling Spray (AP) |
| 6 | A. guillouiae | NFB2 | Both (AP + IS) |
| 7 | A. caviae | KB3 | Seed Inoculation (IS) |
| 8 | A. caviae | KB3 | Seedling Spray (AP) |
| 9 | A. caviae | KB3 | Both (AP + IS) |
| 10 | P. lini | KB4 | Seed Inoculation (IS) |
| 11 | P. lini | KB4 | Seedling Spray (AP) |
| 12 | P. lini | KB4 | Both (AP + IS) |
| 13 | Distilled water | DW | Irrigation |
| 14 | Nutritive solution | NS | Irrigation |
| NO3− | H2PO4− | K+ | Ca2+ | Mg2+ | SO4− | |
|---|---|---|---|---|---|---|
| Percentage Ratio in mol m−3 | ||||||
| Solution Steiner | 12 | 1 | 7 | 9 | 4 | 7 |
| water | 0.17 | 1.36 | 0.24 | 0.23 | ||
| Treatments | PTH | SDM | RDV | LA | R | L | S | R | L | S |
|---|---|---|---|---|---|---|---|---|---|---|
| Tomato Seedlings | Plant Fresh Weight | Plant Dry Matter | ||||||||
| cm | mm | cm3 | cm2 | g | ||||||
| 1 | 9.45 e | 1.87 ef | 1.48 f | 10.72 b | 0.65 ed | 0.32 fe | 0.26 d | 0.05 bcd | 0.04 cdef | 0.02 ed |
| 2 | 10.62 bcd | 2.25 bc | 1.66 de | 9.80 b | 0.85 cde | 0.45 cde | 0.36 cd | 0.03 fe | 0.05 abcd | 0.02 ed |
| 3 | 10.90 bc | 2.26 bc | 1.73 cd | 8.51 bcd | 1.17 bc | 0.473 cd | 0.43 bcd | 0.07 a | 0.06 ab | 0.02 ed |
| 4 | 9.99 cde | 1.84 ef | 1.35 g | 8.70 bcd | 1.07 bcd | 0.47 cd | 0.46 bcd | 0.07 ab | 0.04 bcde | 0.03 bcd |
| 5 | 10.56 bcd | 2.10 cd | 1.61 e | 7.84 bcde | 1.08 bcd | 0.48 bcd | 0.55 bc | 0.05 bcd | 0.04 bcdef | 0.02 ed |
| 6 | 10.93 bc | 2.18 bcd | 1.83 bc | 7.44 bcde | 1.16 bcd | 0.46 cde | 0.47 bcd | 0.06 abc | 0.06 abc | 0.04 b |
| 7 | 9.61 de | 2.01 ed | 1.23 h | 4.99 ed | 0.80 cde | 0.40 def | 0.39 bcd | 0.04 cde | 0.03 efg | 0.02 ed |
| 8 | 10.09 cde | 2.20 bcd | 1.31 gh | 7.62 bcde | 1.00 bcde | 0.56 abc | 0.41 bcd | 0.04 cde | 0.04 def | 0.03 cd |
| 9 | 10.46 cde | 2.30 abc | 1.41 gf | 10.57 b | 1.19 bc | 0.546 abcd | 0.38 bcd | 0.05 bcd | 0.03 efg | 0.03 bcd |
| 10 | 10.25 cde | 2.21 bcd | 1.50 f | 5.18 cde | 0.86 cde | 0.41 cdef | 0.56 bc | 0.04 de | 0.04 cdef | 0.03 bcd |
| 11 | 10.92 bc | 2.23 bc | 1.73 cd | 9.36 bc | 1.91 a | 0.62 ab | 0.85 a | 0.04 de | 0.04 def | 0.03 bcd |
| 12 | 11.61 b | 2.34 ab | 1.96 a | 9.94 b | 1.43 ab | 0.54 abcd | 0.46 bcd | 0.04 de | 0.04 def | 0.04 bc |
| 13 | 7.89 f | 1.78 f | 0.76 i | 4.14 e | 0.48 e | 0.27767 f | 0.27 d | 0.02 e | 0.02 g | 0.01 e |
| 14 | 13.33 a | 2.48 a | 1.93 ab | 15.72 a | 1.14 bcd | 0.64 a | 0.59 b | 0.06 abcd | 0.07 a | 0.06 a |
| Treatments | ENDO | NSB | CMO | MNR |
|---|---|---|---|---|
| µMoles NO2/gpf/h | ||||
| 1 | 6.86 a | 8.26 b | 70.85 f | 120.97 bc |
| 2 | 6.46 abc | 36.89 a | 80.57 cdef | 130.68 a |
| 3 | 6.42 abcd | 37.61 a | 73.72 f | 109.57 de |
| 4 | 6.68 ab | 35.06 a | 77.10 def | 109.39 def |
| 5 | 6.59 abc | 39.84 a | 83.83 cd | 101.78 fg |
| 6 | 6.23 bcde | 7.72 b | 77.20 def | 108.44 defg |
| 7 | 6.49 abc | 38.61 a | 85.19 bc | 105.014 efg |
| 8 | 6.46 abc | 35.86 a | 77.63 def | 130.18 a |
| 9 | 6.17 bcde | 35.92 a | 94.62 a | 100.84 g |
| 10 | 6.23 bcde | 38.50 a | 73.18 f | 126.73 ab |
| 11 | 6.09 cde | 36.42 a | 91.25 ab | 128.53 ab |
| 12 | 6.16 bcde | 39.78 a | 80.79 cde | 131.86 a |
| 13 | 5.93 de | 36.99 a | 81.22 cd | 110.25 de |
| 14 | 5.79 e | 34.76 a | 81.75 cd | 115.94 cd |
| Treatments | N | P | K | Ca | Mg | Na | Mn | Ni | Fe | Zn | Cu |
|---|---|---|---|---|---|---|---|---|---|---|---|
| % | mg kg−1 | ||||||||||
| 1 | 1.50 cd | 0.24 bcde | 0.49 cd | 1.20 b | 0.16 e | 0.051 ab | 26.21 dc | 3.62 c | 64.50 def | 32.10 b | 2.62 c |
| 2 | 1.48 cd | 0.31 abcde | 0.65 bcd | 1.27 b | 0.22 cde | 0.049 abcde | 38.02 abcd | 4.21 bc | 81.50 bcd | 32.51 b | 4.01 c |
| 3 | 1.91 abcd | 0.32 ab | 0.79 bcd | 0.87 c | 0.24 cde | 0.043 abcde | 35.02 abcd | 4.71 abc | 96.72 bc | 41.74 a | 3.11 c |
| 4 | 1.45 cd | 0.22 de | 0.77 bcd | 1.12 bc | 0.24 bcde | 0.04 e | 35.82 abcd | 4.12 bc | 60.13 defg | 37.62 ab | 4.46 c |
| 5 | 1.91 abcd | 0.31 abc | 0.47 cd | 1.13 bc | 0.20 ed | 0.05 abc | 45.95 ab | 4.30 bc | 95.25 bc | 37.80 ab | 3.75 c |
| 6 | 2.26 ab | 0.28 abcde | 0.41 cd | 1.27 b | 0.26 bcde | 0.049 abcde | 46.48 ab | 4.57 abc | 165.51 a | 15.27 cd | 4.84 c |
| 7 | 1.26 cd | 0.20 e | 0.62 bcd | 0.20 g | 0.25 cde | 0.05 abc | 50.02 a | 5.02 ab | 81.01 bcd | 10.01 d | 4.61 c |
| 8 | 1.98 abc | 0.27 bcde | 1.03 b | 0.27 fg | 0.26 cde | 0.05 abc | 27.64 dc | 4.41 abc | 52.82 efgh | 11.42 d | 3.67 c |
| 9 | 1.66 bcd | 0.34 a | 0.91 bc | 0.86 cd | 0.37 bc | 0.05 abc | 22.72 d | 5.10 ab | 47.04 fgh | 13.96 cd | 8.98 b |
| 10 | 2.10 abc | 0.21 e | 0.41 cd | 0.53 ef | 0.12 e | 0.043 bcde | 25.44 d | 4.64 abc | 38.1 gh | 14.58 cd | 3.81 c |
| 11 | 2.24 ab | 0.27 abcde | 0.47 cd | 0.59 de | 0.42 b | 0.047 abcde | 31.71 bcd | 4.82 ab | 75.86 cde | 17.47 cd | 21.42 a |
| 12 | 1.98 abc | 0.32 ab | 0.47 cd | 1.04 bc | 0.68 a | 0.051 a | 43.41 abc | 5.21 ab | 103.72 b | 21.57 c | 4.63 c |
| 13 | 1.74 bcd | 0.22 cde | 0.33 d | 0.19 g | 0.18 e | 0.042 de | 22.10 d | 5.01 ab | 36.22 gh | 15.78 cd | 3.67 c |
| 14 | 2.51 a | 0.20 e | 2.09 a | 1.75 a | 0.35 bcd | 0.041 de | 50.90 a | 5.48 a | 75.42 cde | 31.07 b | 3.82 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cervantes-Vázquez, T.J.Á.; Valenzuela-García, A.A.; Cervantes-Vázquez, M.G.; Guzmán-Silos, T.L.; Fortiz, E.L.; Rangel, P.P.; Rueda-Puente, E.O. Morphophysiological, Enzymatic, and Elemental Activity in Greenhouse Tomato Saladette Seedlings from the Effect of Plant Growth-Promoting Rhizobacteria. Agronomy 2021, 11, 1008. https://doi.org/10.3390/agronomy11051008
Cervantes-Vázquez TJÁ, Valenzuela-García AA, Cervantes-Vázquez MG, Guzmán-Silos TL, Fortiz EL, Rangel PP, Rueda-Puente EO. Morphophysiological, Enzymatic, and Elemental Activity in Greenhouse Tomato Saladette Seedlings from the Effect of Plant Growth-Promoting Rhizobacteria. Agronomy. 2021; 11(5):1008. https://doi.org/10.3390/agronomy11051008
Chicago/Turabian StyleCervantes-Vázquez, Tomás Juan Álvaro, Ana Alejandra Valenzuela-García, María Gabriela Cervantes-Vázquez, Tania Lizzeth Guzmán-Silos, Erika Lagunes Fortiz, Pablo Preciado Rangel, and Edgar Omar Rueda-Puente. 2021. "Morphophysiological, Enzymatic, and Elemental Activity in Greenhouse Tomato Saladette Seedlings from the Effect of Plant Growth-Promoting Rhizobacteria" Agronomy 11, no. 5: 1008. https://doi.org/10.3390/agronomy11051008
APA StyleCervantes-Vázquez, T. J. Á., Valenzuela-García, A. A., Cervantes-Vázquez, M. G., Guzmán-Silos, T. L., Fortiz, E. L., Rangel, P. P., & Rueda-Puente, E. O. (2021). Morphophysiological, Enzymatic, and Elemental Activity in Greenhouse Tomato Saladette Seedlings from the Effect of Plant Growth-Promoting Rhizobacteria. Agronomy, 11(5), 1008. https://doi.org/10.3390/agronomy11051008