4. Discussion
Since 1970, significant efforts have been made to explore agricultural practices for reducing aggravating plant protection substances, both for consumers and for the environment, without reducing the quantity and quality of the end product. The need to reduce external inputs in agricultural systems is a challenge for breeders and growers due to reduced production costs [
58]. Thus, researchers’ interest turned to alternative production systems and methods, such as organic farming and low input agriculture, which were considered more sustainable than conventional farming [
59]. However, few studies have been published on production’s economic viability in low input systems [
60]. This information is critical for those wishing to adopt this system, especially in crops that require high inputs, such as tomato cultivation [
61]. The use of appropriate cultivars to meet specific cultivation systems is essential, considering that many cultivars which developed under high input conditions have failed to meet producers’ requirements in low input systems [
62]. This fact necessitates the creation of suitable cultivars as, according to Lammerts van Bueren, etc. [
63], more than 95% of organic farming is based on high-input cultivars. For this reason, the creation of cultivars intended for cultivation in conditions of lower inputs requires their breeding process to take place in similar conditions [
64,
65,
66]. Our study focused on the production of tomato recombinant pure lines under low input farming conditions, targeting these kind farming systems (low inputs and organic cultivation). Tomato commercial hybrids Formula F
1 and Elpida F
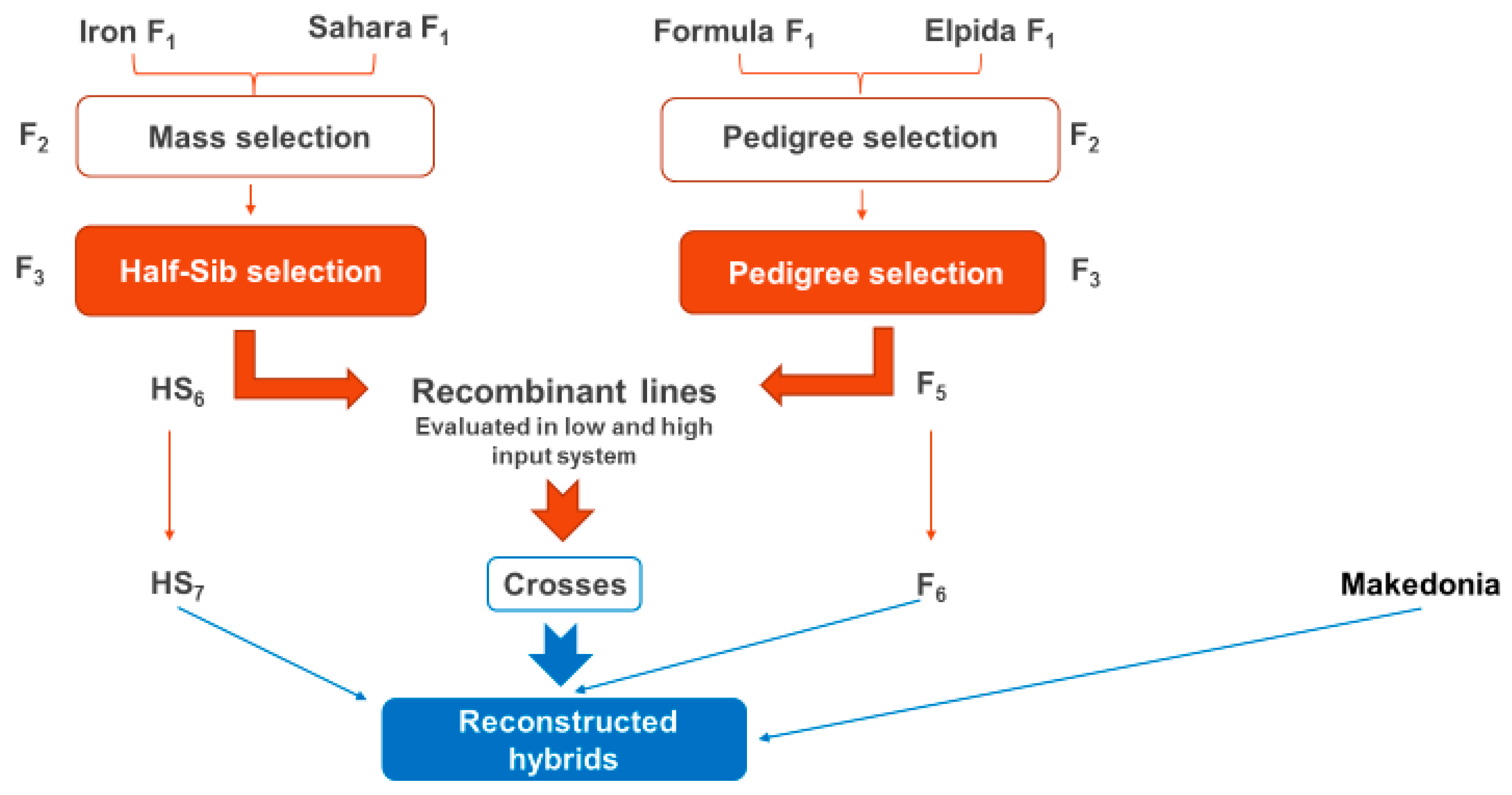
1, the recombinant lines derived from them and from the hybrids ‘Iron’ and ‘Sahara’, the cultivar ‘Makedonia’, and the second cycle reconstructed hybrids, produced after crossing these inbred lines, were evaluated under low input conditions.
Nowadays, there is a demand for the development of high-yielding tomato inbred cultivars or, in some cases, hybrids for increased yield, even in the least productive areas [
67]. Over the last few years, inbred lines’ replacement by hybrids has remarkably increased yield [
68,
69]. In general, hybrids are preferred over pure line varieties in tomato because of their superiority in marketable fruit yield, component traits, and fruit quality [
70]. The exploitation of heterosis proves an efficient approach for the improvement of tomato, with the tomato’s hybrid varieties showing high yield potential and gaining popularity among growers [
71]. In many countries, including Greece, the hybrid is the predominant type of commercial cultivar. Tomato hybrids dominate in high-input agricultural systems (greenhouse cultivation/hydroponics) and may be extended to the lower-input systems, where economic benefit can be demonstrated [
72]. In our study, we have proven that hybrids which are structured from recombinant lines and developed under low input conditions are suitable for such cultivation conditions.
Earliness is one of the primary criteria in every breeding program [
73]. This study’s genetic materials did not differ significantly regarding fruit weight, as it was shown for the characteristic of fruit number per plant. As such, the latter seems to be the reason why, in total yield, there were statistically significant differences. The lines having high number of fruits per plant are characterized by high yielding potential. Recombinant inbred lines ‘For-2′ and ‘Elp-3′ are promising genetic materials for earliness. Furthermore, four reconstructed hybrids exceeded both original hybrids, ‘Formula’ and ‘Elpida’, in absolute values
The ‘Elpida’ F1 hybrid possessed a higher level of inbreeding depression (28%) than the ‘Formula’ F1 hybrid (16%), both of which presented tolerance to inbreeding and were not prohibitive in designing breeding programs for the production of inbred lines.
Patel et al. [
71] stated that the crosses exhibiting high heterobeltiosis with low inbreeding depression were characterized by the accumulation of favorable additive genes for the traits evaluated. The inbred line ‘Elp-4′ reached the productivity in total yield of the original hybrid ‘Elpida’ F
1, and surpassed the ‘Formula’ F
1 hybrid by 6%. This reinforces the view of it being possible to isolate inbred lines superior to hybrids [
52,
74]. Although tomato hybrids are the dominant type of variety, studies conducted by many researchers confirm the ’recovery’ of inbred lines through selection by hybrids, thus supporting inbred vigor. Experimental studies on the ‘isolation’ of inbred lines by breeding and selecting tomato hybrids reinforce the above position [
45,
49,
52,
75]. At the same time, it has been found that, through selection methodologies, the frequency of alleles with additive action increases, resulting in inbred lines that could yield, theoretically, as much as hybrids [
76]. According to Syukur et al. [
77], the characters which are controlled by additive genes will be easier to select, mainly for improving inbred line varieties. Sharma et al. [
78], Kumaravel et al. [
79], and Hosamani [
80] found that additive variance was consistently greater than non-additive variation, indicating that the major portion of the genetic variation for the characters number of fruits per plant and total fruits yield per plant was controlled by genes with additive effects [
81]. Analogous results are shown in the present study. New breeding objectives should consider the concept of agricultural sustainability by maintaining high yield and fruit quality under low input farming practices, especially under adverse climate conditions [
82]. Each of the recombinant inbred lines developed under low input farming systems in this study are characterized by earliness, high yield, and desirable fruit quality traits, which could be cultivars targeted to low input farming systems.
In the case of the isolation of recombinant inbred lines with a yielding potential equivalent to hybrids, a question arises about the behavior of hybrids resulting from these superior inbred lines. According to Guan Yi-Xin, et al. [
83], homozygous parental lines produce restructured F
1 hybrids with a high degree of heterosis in many characteristics. This was the primary task of this study. The recombinant lines produced with base material dynamic commercial hybrids under honeycomb pedigree methodology were proven to be equal to, or even better than, the original hybrids. This means that additive genes replaced deleterious recessives, and inbred vigor surpassed hybrid vigor. In such a situation, it is necessary to check whether the specific genetic materials can, by crossing them, create additional heterosis; therefore, if there is still a sufficiently high genetic load of recessive deleterious genes, they, in the state of heterozygosity, will cover their action behind the action of dominant genes.
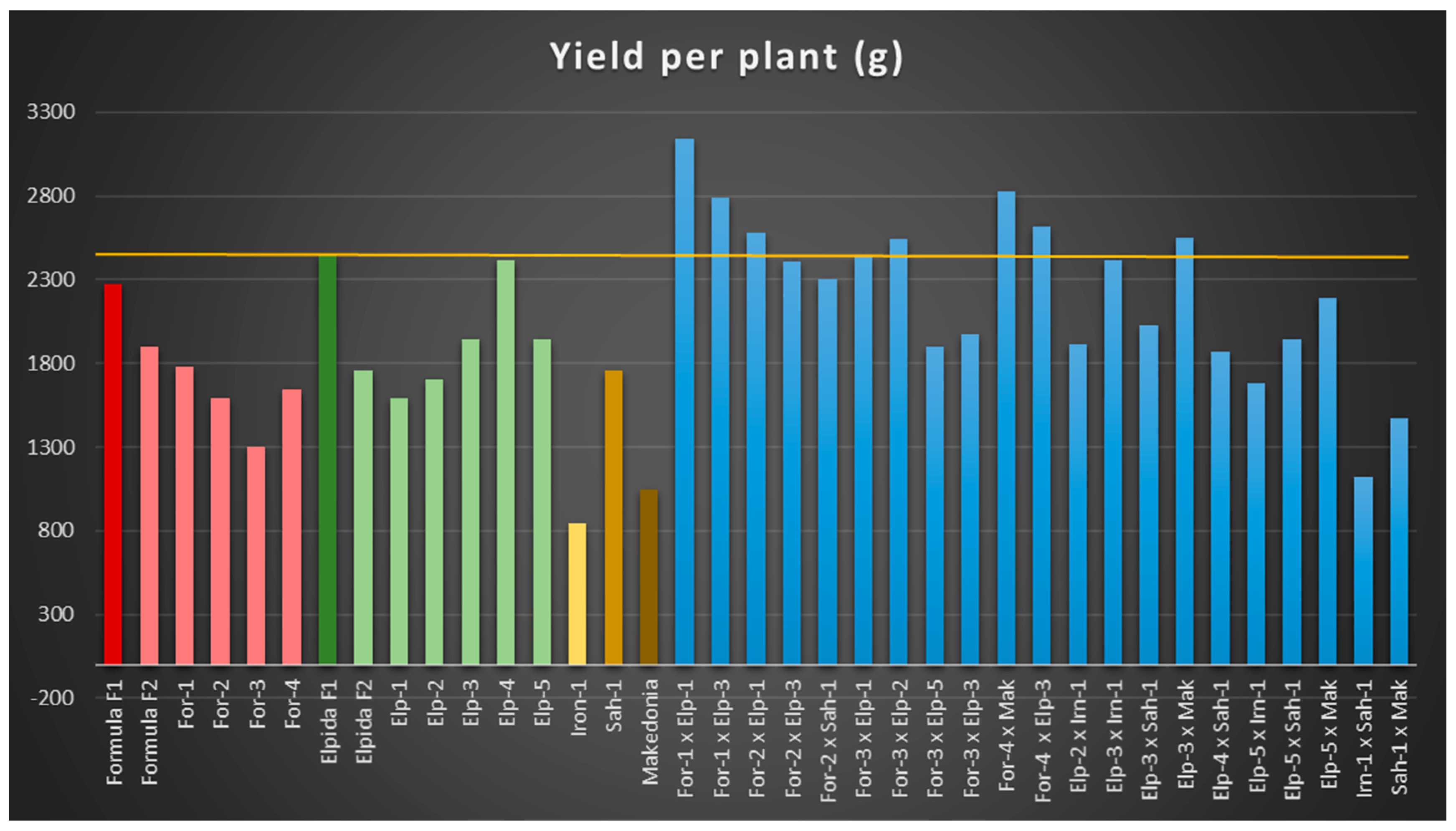
The study of the total yield of genetic materials revealed no significant superiority of seven reconstructed hybrids from the original F1 hybrids ‘Formula’ and ‘Elpida’, with ‘For1 × Elp1′ reaching 3140.9 g per plant. These reconstructed hybrids produced under low input farming systems could be an alternative type of cultivar suitable for such systems.
An increase in production is associated with an increase in the number of fruits per plant or/and in fruit weight [
75]. Avdikos et al. [
52] reported a high positive correlation between yield per plant and number of fruits per plant (r = 0.97), while a low correlation was observed between yield per plant and fruit weight per plant (r = 0.27). The results indicated the importance of the characteristic of the number per plant in a breeding program. Solieman et al. [
84] reported a positive correlation between the total yield and the number of fruits per plant, while the correlation was negative between the number of fruits per plant and fruit weight. This is confirmed by our results, as the reconstructed hybrids, (‘For-1 × Elp-1′) and (‘For-4 × Mak’), scored the highest yield, with the highest number of fruits per plant. Similarly, the recombinant ‘Elp-4′ line, provided a high yield and a high number of fruits per plant, whereas the ‘Irn-1′ recombinant line showed the lowest yield and the lowest number of fruits per plant. It is noteworthy that the inbreeding depression for fruit number per plant in F
2 generation of ‘Formula’ and ‘Elpida’ hybrids is similar to that recorded for total yield (‘Formula’ F
2: 16% total yield—19% fruit number, ‘Elpida’ F
2: 28% total yield—27% fruit number).
Almost all reconstructed hybrids possessed positive Relative heterosis regarding early yield, reaching 140% (‘For 4 × Mak’). A total of 13 out of 21 reconstructed hybrids showed positive heterobeltiosis, with ‘For 4 × Mak’ reaching 120%. All reconstructed hybrids presented high Standard Heterosis over ‘Makedonia’ in earliness. Over the mid-value of ‘Formula’ and ‘Elpida’ hybrids, we produced six reconstructed hybrids that had positive values reaching 36%. Thus, it was shown that, in second cycle there were new reconstructed hybrids produced which provided higher yields in earliness than the commercial hybrids.
Total fruit yield per plant is one of the most important traits, and it deserves the highest attention in any breeding program. Almost all hybrids showed positive Relative heterosis in total yield, reaching 86% (‘For1 × Elp1′). In heterobeltiosis, 15 reconstructed hybrids showed positive heterosis, from 1% to 77%. All second cycle reconstructed hybrids exhibited standard heterosis over domestic cultivar ‘Makedonia’, which varied from 7% to 200%. When compared with commercial hybrids, the reconstructed hybrids possessed Standard heterosis from 2% to 33%. The reconstructed hybrids ‘For1 × Elp1′, ‘For4 × Mak’ and ‘For1 × Elp3′ recorded the highest values in Standard heterosis, and appear as promising material for cultivation, at least in low input farming systems. Rehana et al. [
73] found six crosses exhibiting the highest positive heterosis for yield per plant, having values ranging from 26.46% to 88.46% in heterobeltiosis and from 16.09% to 48.44% in Relative heterosis. Rasheed et al. [
85] reported that the maximum better parent heterosis was 120.6%. Khan and Jindal [
67], with relevance to total yield, found that the crosses revealed significant and positive heterosis over better parent, ranging from 19.6% to 103.1%, and heterosis over standard check, ranging from 26.4% to 130.7%. Solieman et al. [
84] reported heterosis ranging from 22.29% to 64.33% for total fruit yield per plant. Heterosis for fruit yield was also reported by Thakur et al. [
86], Premalakshme et al. [
87], Harer et al. [
88], and Sharma and Thakur [
89].
The number of fruits per plant is a major yield contributing character [
16]. Fageria et al. [
90], Kurian et al. [
14], and Singh et al. [
91] observed a significant positive heterosis for a higher number of fruits per plant in tomato, suggesting good scope for yield improvement through its components. Regarding fruit number per plant, both in earliness and total production, almost all genetic materials showed positive Relative heterosis (reaching 101%). A similar situation was noticed for heterobeltiosis, with the best hybrid in earliness, ‘For 4 × Mak’, possessing 80%, and the best hybrid in total production, ‘For1 × Elp1′, possessing 74%. The Standard heterosis over ‘Makedonia’ was extremely high in both earliness and total production (reaching 198%). Eighteen and fourteen reconstructed hybrids out of twenty-one showed positive Standard heterosis over commercial hybrids in early and total production, respectively. Ahmad et al. [
15] found for the trait of fruit number that the range of positive heterosis was 3.8% to 83.9% over the better parent. Rehana et al. [
73] recorded the trait of fruit number per plant, heterobeltiosis, and Relative heterosis from 17.26% to 49.25%, respectively. Kumari and Sharma [
16] found 66.08% maximum heterosis over the better parent. Similar findings were also reported by Legon et al. [
92], Jamwal et al. [
93], Mirshamssi et al. [
94], Rani et al. [
95], and Ahmad et al. [
15], for higher fruit number per plant.
In terms of fruit weight, the results were not the same as in yield and number of fruits per plant. Most reconstructed hybrids showed negative values in Relative heterosis and heterobeltiosis, revealing that the higher yields shown for yield are only due to the characteristic number of fruits per plant. Some hybrids showed negative values for Standard heterosis over ‘Makedonia’ and all commercial hybrids. Heterotic analysis by Rasheed et al. [
85] depicted positive and significant heterosis, as well as heterobeltiosis for individual fruit weight in cross combinations. Maximum heterosis and heterobeltiosis were 88.5% and 54.3%, respectively. Ghadage et al. [
96] found that heterosis over better parent for average fruit weight ranged from −46.1% to 34.7%, whereas heterosis over standard check ranged from −35.4% to 32.9%. Rehana et al. [
73] recorded fruit weight maximum Relative heterosis and heterobeltiosis at 63.5% and 48.3%, respectively. Significant positive heterosis for this character has been reported by Mondal et al. [
97], Kumari and Sharma [
16], Savale et al. [
98], Tamata and Singh [
99], Gautam et al. [
100], Kattegoudar et al. [
101], and Mohammad l. Al-Daej, [
102].
A morphological feature of the tomato fruit is pericarp thickness [
103]. Large pericarp thickness expresses a fruit’s firmness and is considered a desirable characteristic for those fruits which are intended to be transported, as the duration of post-harvest life is extended. However, consumers favor fruits with a small pericarp thickness. The largest pericarp thickness was found in the original ‘Elpida’ F
1 hybrid, followed by the reconstructed hybrid (‘Elp-2 × Irn-1′). Most recombinant lines, from both the Formula and the ‘Elpida’ family, produced fruits with a large pericarp size, with no statistically significant differences from the original Elpida F
1 hybrid. It is worth noting that the reconstructed hybrid (‘For-2 × Elp-1′), recorded the lowest pericarp thickness (4.7 mm), although the parental lines For-2 and Elp-1 exhibited, as individuals, a large pericarp thickness (6 and 6.8 mm, respectively). Almost all reconstructed hybrids recorded negative values for Relative heterosis, heterobeltiosis, and Standard heterosis over commercial hybrids regarding pericarp thickness. Ghadage et al. [
96] found a range of heterosis over better parent for pericarp, from −22.9% to 14.7%, whereas heterosis over standard check ranged from −10.9% to 25.3%. Kumari and Sharma [
16] reported a maximum heterotic effect of pericarp thickness over better parent by 19.37%. Significant positive heterosis for this character has been reported by Joshi et al. [
104], Rattan [
105], Garg et al. [
106], and Sureshkumara et al. [
107].
From a quality point of view, reducing locule number is desirable and a negative estimate of heterosis is valuable [
108]. Thus, to reduce the number of locules per fruit, heterosis breeding can be exploited efficiently. The locule number of the fruit is an indicator of fruit firmness. A smaller number of locules per fruit increases fruit firmness and vice versa. Wild tomato species produce fruit with 2 to 4 locules, while cultivated varieties can develop more than 15 locules. This affects fruit shape dramatically and leads to a 50% increase in fruit size [
109]. Consumers usually prefer fruits with a small number of locules. Among the genetic materials studied, the largest locule number was noted by the recombinant line ‘Sah-1′, followed by the domestic cultivar ‘Makedonia’, with 8.9 and 7.5 locules, respectively. Primarily, statistically significant differences were observed in both recombinant lines and reconstructed hybrids for the fruit’s locule number, not consistent with the materials’ fruit weight, where no statistically significant differences were observed. As for this trait, the majority of hybrids had negative estimates for all types of heterosis. Ahmad et al. [
15] found that about 50% of combinations showed positive heterosis over the better parent, ranging from 5.4% to 27.5%. Rasheed et al. [
85] recorded that nine hybrids exhibited positive heterosis, while six hybrids revealed negative heterosis regarding the number of locules per fruit, with a maximum heterosis of 32.4% and a heterobeltiosis of 22.7%. Maximum negative heterosis and heterobeltiosis were −39.39% and −46.42%, respectively.
The dominance model (masking of deleterious recessive alleles) presents heterosis as a simple reversal of inbreeding depression (unmasking of deleterious recessive alleles). The dominance model suffers from certain limitations, which suggest that it is only a partial explanation for the phenomenon of heterosis. An essential criticism of this model is that, if complementation of deleterious alleles is causal for heterosis, then the potential to generate heterosis by crossing commercially available inbred lines should decrease over time [
110]. Elite germplasm has been exploited in breeding programs for many years and, during this period, the majority of slightly deleterious alleles would have been expected to have been purged [
111]. Models of heterosis relying entirely on the concept of dominance would predict that the potential for heterosis should also have decreased over the same period [
112]. Tomato is an autogamous species which has a small load of deleterious recessive genes. Besides, the continuous selfing and selection of the desired genotypes in the breeding process resulted in inbred recombinant lines, with inbred vigor equal to or even higher than hybrid vigor [
52]. This means that a large portion of the small pool of recessive deleterious alleles has been replaced by a desirable additive. It would be expected that, according to the dominance model, the heterosis recorded after crossing the recombinant lines would be minimal. However, as the results of this study showed, the elite genotypes of recombinant inbred lines became the parents of elite restructured hybrids, with increased levels of re-heterosis for all characters measured. This may prove that dominance is not the only case in explaining heterosis in tomato for yield components and fruit quality characteristics.



 ,
, 






{kind=link}
{kind=link}
{kind=link}