
Exploratory Assessment to Evaluate Seed Sprouting under Elevated CO2 Revealed Improved Biomass, Physiology, and Nutritional Value of Trachyspermum ammi

 ,
, 

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
2.2. Physiological Analyses
2.3. Determination of Primary Metabolises
2.4. Measurement of Secondary Metabolises
2.5. Evaluation of Biological Activities
2.6. Statistical Analysis
3. Results and Discussion
3.1. Effect of eCO2 on the Growth, Photosynthesis, and Respiration of T. ammi Sprouts
3.2. Sprouting and eCO2 Improved the Tissue Quality of T. ammi
3.2.1. Improved Photosynthesis Induced the Primary Metabolism
3.2.2. Improved Photosynthesis Induced the Secondary Metabolism
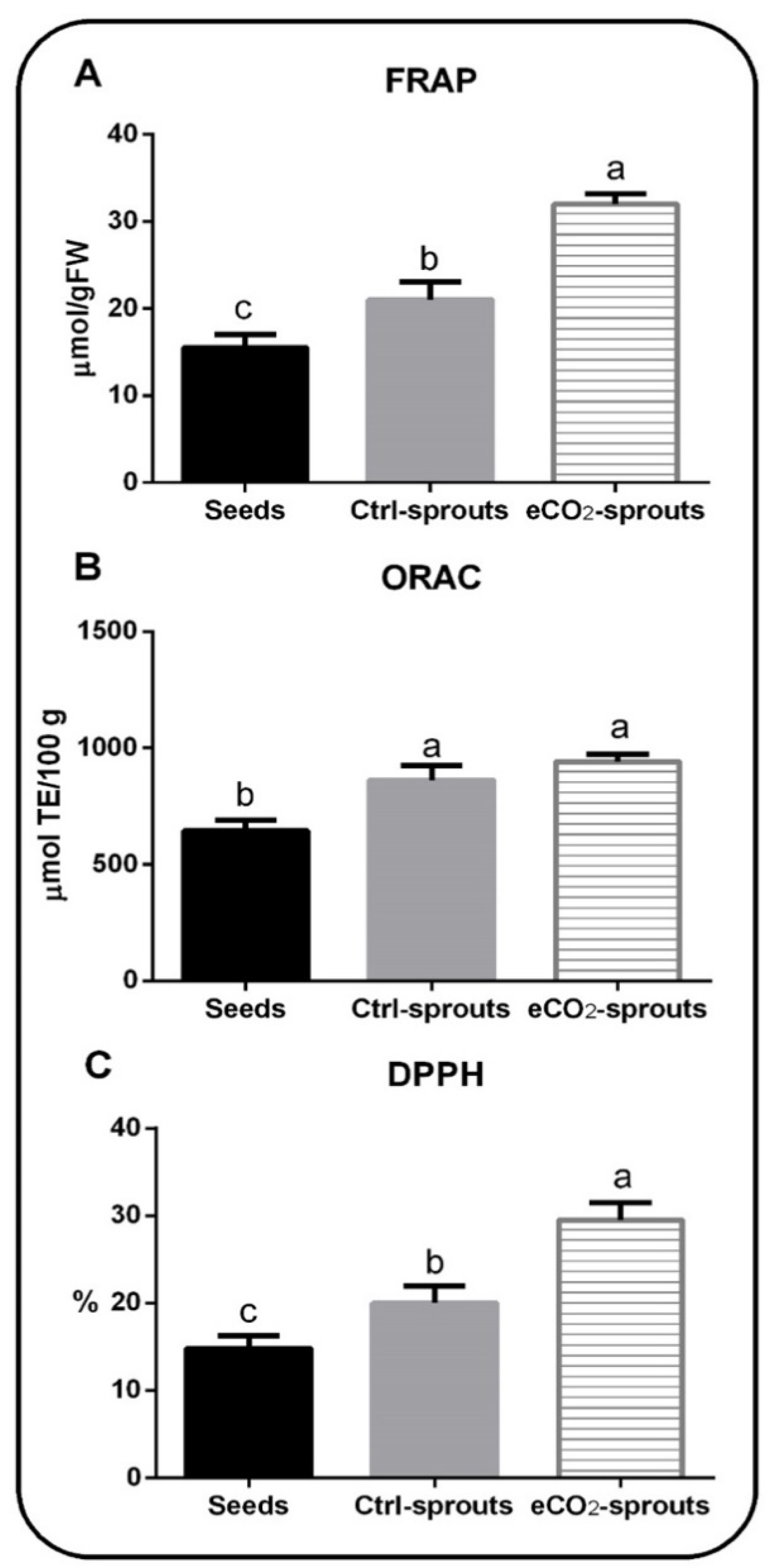
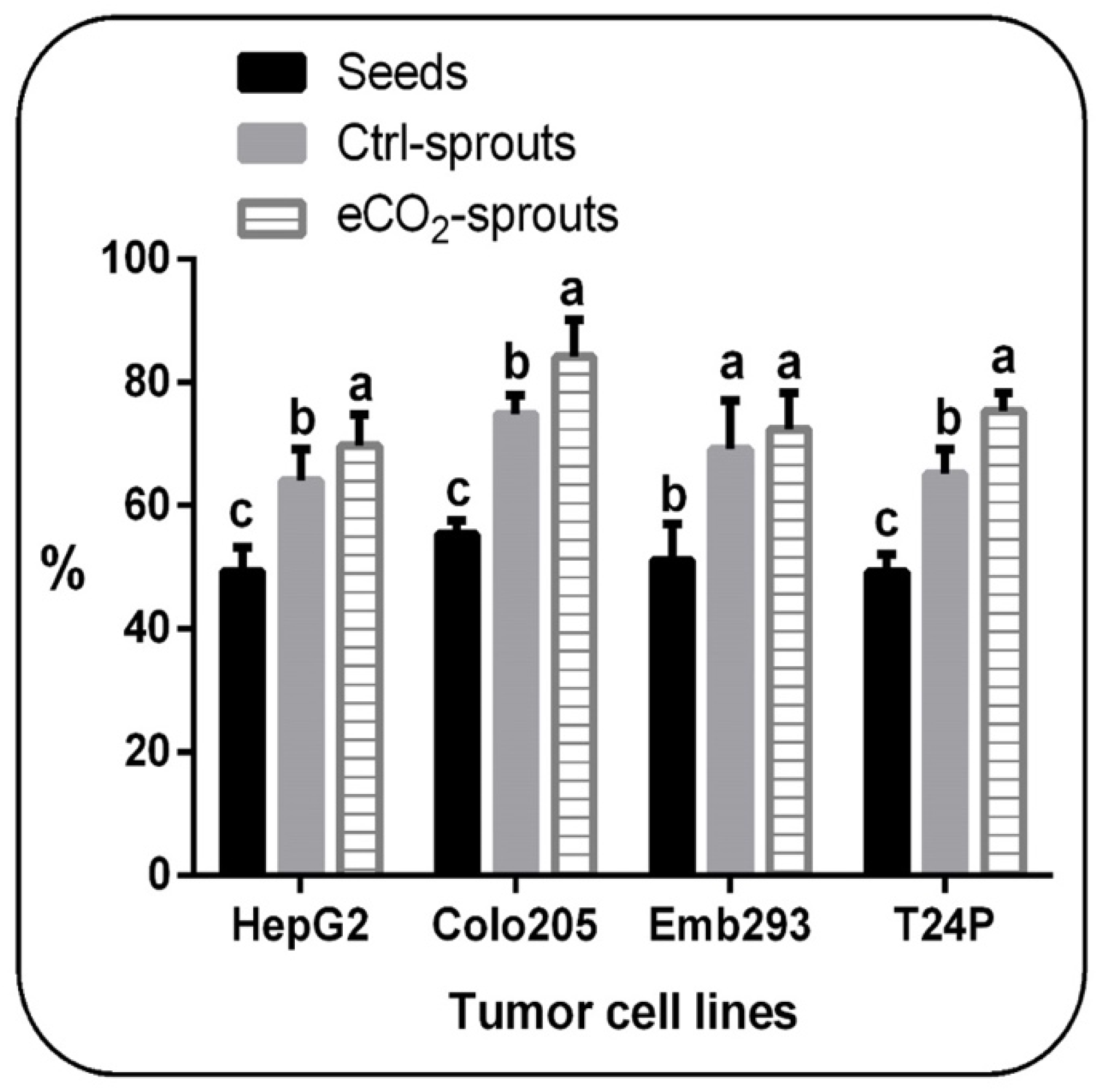
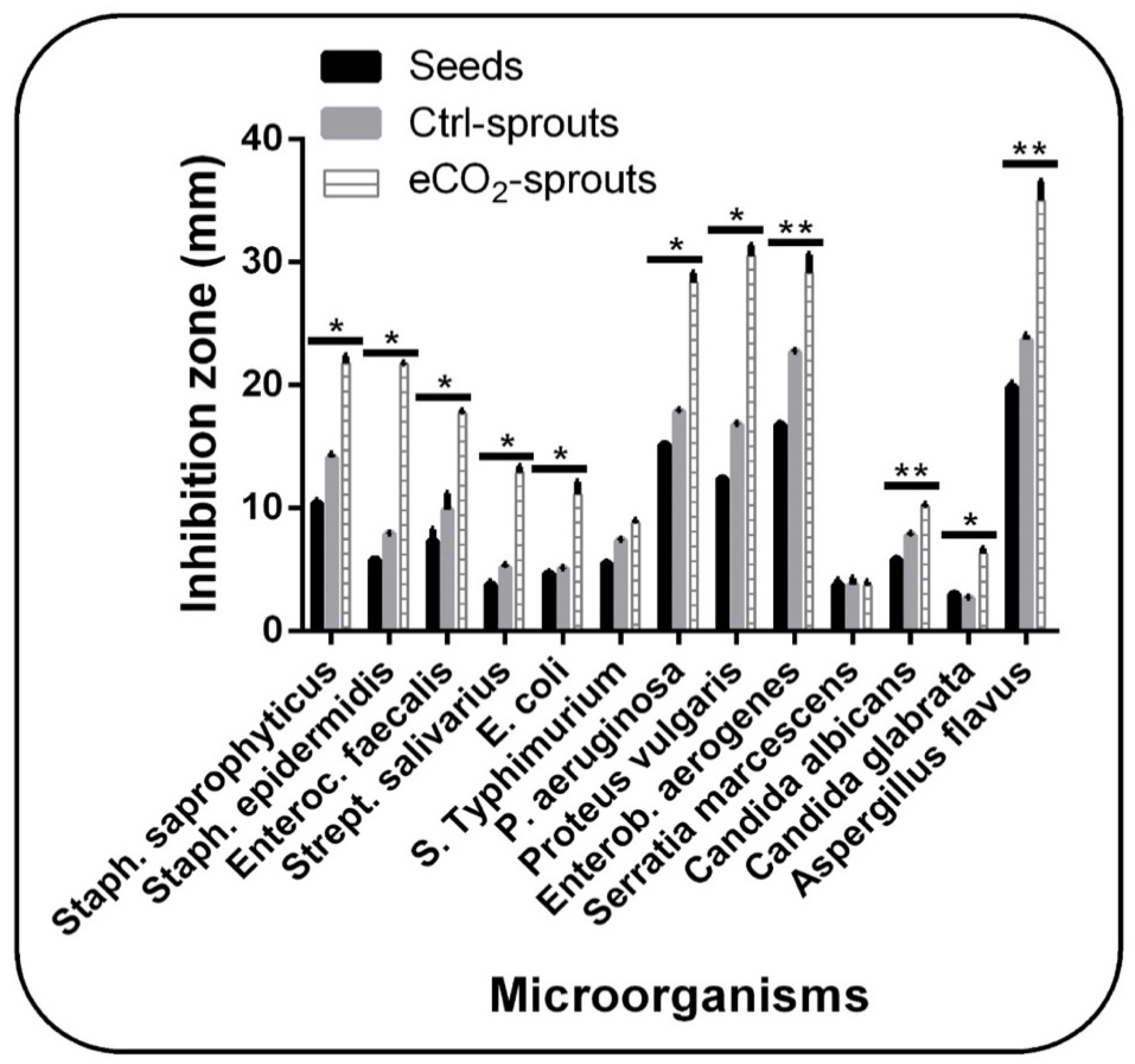
3.3. Sprouting and eCO2 Enhanced the Biological Activities of T. ammi
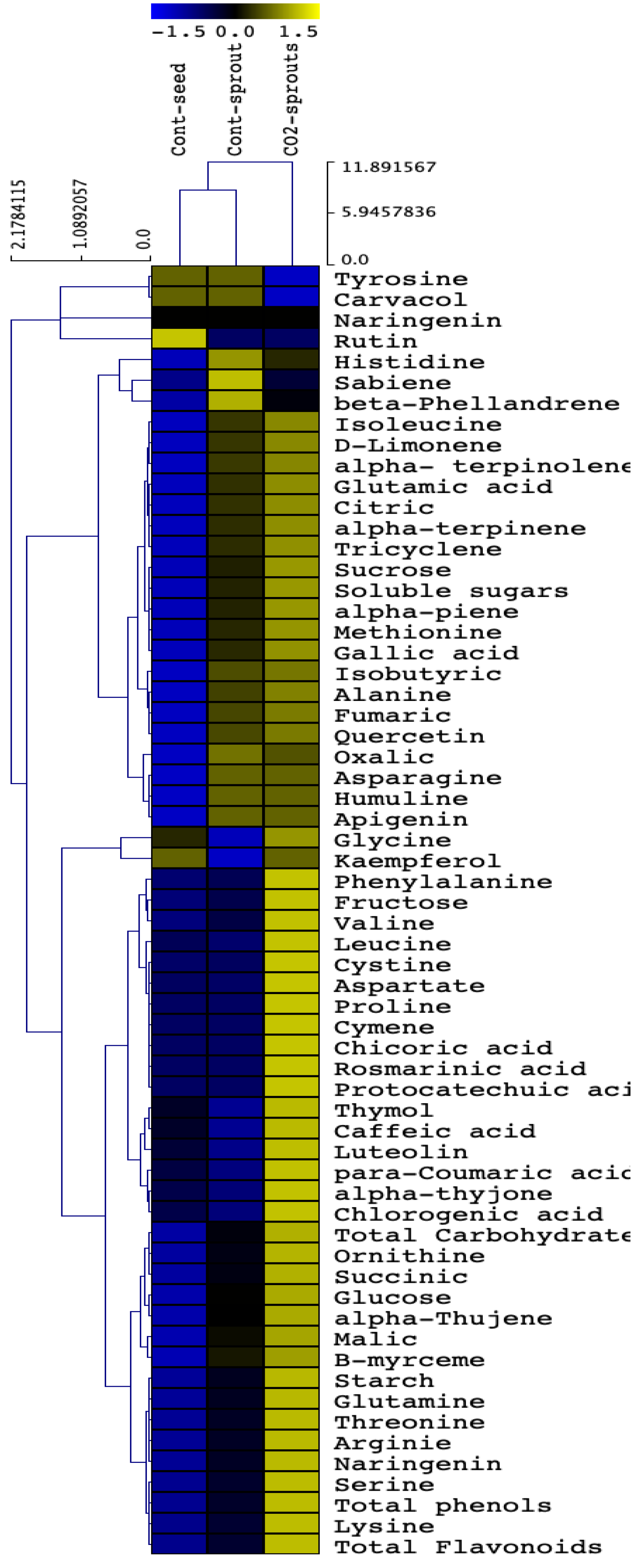
3.4. Hierarchical Analysis Findings
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marton, M.; Mandoki, Z.; Caspo, J.; Caspo-Kiss, Z. The role of sprouts in human nutrition. A review. Aliment. Hung. Univ. Transylvania 2010, 3, 81–117. [Google Scholar]
- Mattioli, S.; Dal Bosco, A.; Castellini, C.; Falcinelli, B.; Sileoni, V.; Marconi, O.; Mancinelli, A.C.; Cotozzolo, E.; Benincasa, P. Effect of heat- and freeze-drying treatments on phytochemical content and fatty acid profile of alfalfa and flax sprouts. J. Sci. Food Agric. 2019, 99, 4029–4035. [Google Scholar] [CrossRef]
- Dal Bosco, A.; Castellini, C.; Martino, M.; Mattioli, S.; Marconi, O.; Sileoni, V.; Benincasa, P. The effect of dietary alfalfa and flax sprouts on rabbit meat antioxidant content, lipid oxidation and fatty acid composition. Meat Sci. 2015, 106, 31–37. [Google Scholar] [CrossRef]
- Silva, L.R.; Pereira, M.J.; Azevedo, J.; Gonçalves, R.F.; Valentão, P.; de Pinho, P.G.; Andrade, P.B. Glycine max (L.) Merr., Vigna radiata L. and Medicago sativa L. sprouts: A natural source of bioactive compounds. Food Res. Int. 2013, 50, 167–175. [Google Scholar] [CrossRef]
- Dias, D.A.; Urban, S.; Roessner, U. A historical overview of natural products in drug discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [Green Version]
- Mamadalieva, N.Z.; Doniyor, R.; Sagdullaev, S.S. Medicinal Plants of the Apiaceae and Rutaceae Families from the Chimgan Mountains (Uzbekistan): Ethnopharmacology, Chemical Composition and Biological Activities. Curr. Tradit. Med. 2018, 4, 166–183. [Google Scholar] [CrossRef]
- Morsi, N.M. Antimicrobial effect of crude extracts of Nigella sativa on multiple antibiotics-resistant bacteria. Acta Biochim. Pol. 2000, 49, 63–74. [Google Scholar]
- Kowalczyk, A.; Przychodna, M.; Sopata, S.; Bodalska, A.; Fecka, I. Thymol and Thyme Essential Oil-New Insights into Selected Therapeutic Applications. Molecules 2020, 25, 4125. [Google Scholar] [CrossRef]
- Singh, G.; Maurya, S.; Catalan, C.; De Lampasona, M.P. Chemical constituents, antifungal and antioxidative effects of ajwain essential oil and its acetone extract. J. Agric. Food Chem. 2004, 52, 3292–3296. [Google Scholar] [CrossRef]
- Sivropoulou, A.; Papanikolaou, E.; Nilolaou, C.; Kokkini, S.; Lanaras, T.; Arsenakis, M. Antimicrobial and cytotoxic activities of origanum essential oils. J. Agric. Food Chem. 1996, 44, 1202–1205. [Google Scholar] [CrossRef]
- Srivastava, K.C. Extract of a spice Omum (Trachyspermum ammi) shows antiaggregatory effects and alters arachidonic acid metabolism in human platelets. Prostaglandins Leukot Essent. Fat. Acids 1988, 33, 1–6. [Google Scholar] [CrossRef]
- Björkman, M.; Klingen, I.; Birch, A.N.E.; Bones, A.M.; Bruce, T.J.A.; Johansen, T.J. Phytochemicals of Brassicaceae in plant protection and human health-Influences of climate, environment and agronomic practice. Phytochemistry 2011, 72, 538–556. [Google Scholar] [CrossRef] [PubMed]
- Baenas, N.; García-Viguera, C.; Moreno, D.A. Elicitation. A tool for enriching the bioactive composition of foods. Molecules 2014, 19, 13541–13563. [Google Scholar] [CrossRef] [Green Version]
- Saleh, A.M.; Selim, S.; Al Jaouni, S.; AbdElgawad, H. CO2 enrichment can enhance the nutritional and health benefits of parsley (Petroselinum crispum L.) and dill (Anethum graveolens L.). Food Chem. 2018, 269, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Leakey, A.D.; Xu, F.; Gillespie, K.M.; McGrath, J.M.; Ainsworth, E.A.; Ort, D.R. Genomic basis for stimulated respiration by plants growing under elevated carbon dioxide. Proc. Natl. Acad. Sci. USA 2009, 9, 3597–3602. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, K.; Watanabe, C.K.; Terashima, I. Effects of elevated atmospheric CO2 on primary metabolite levels in Arabidopsis thaliana Col-0 leaves: An examination of metabolome data. Plant Cell Physiol. 2015, 56, 2069–2078. [Google Scholar]
- Almuhayawi, M.; AbdElgawad, H.; Al Jaouni, S.; Selim, S.; Hassan, A.H.; Khamis, G. Elevated CO2 improves glucosinolate metabolism and stimulates anticancer and anti-inflammatory properties of broccoli sprouts. Food Chem. 2020, 328, 127102. [Google Scholar] [CrossRef]
- Almuhayawi, M.S.; Hassan, A.H.; Al Jaouni, S.K.; Alkhalifah, D.H.M.; Hozzein, W.N.; Selim, S.; Khamis, G. Influence of elevated CO2 on nutritive value and health-promoting prospective of three genotypes of Alfalfa sprouts (Medicago sativa). Food Chem. 2020, 340, 128–147. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Peshev, D.; Zinta, G.; Van den Ende, W.; Janssens, I.A.; Asard, H. Climate extreme effects on the chemical composition of temperate grassland species under ambient and elevated CO2: A comparison of fructan and non-fructan accumulators. PLoS ONE 2014, 9, e92044. [Google Scholar] [CrossRef]
- Hamad, I.; AbdElgawad, H.; Al Jaouni, S.; Gaurav, Z.; Han, A.; Sherif, H.; Momtaz, H.; Nashwa, H.; Samy, S. Metabolic analysis of various date palm fruit (Phoenix dactylifera L.) cultivars from Saudi Arabia to assess their nutritional quality. Molecules 2015, 20, 13620–13641. [Google Scholar] [CrossRef]
- Hassanpour, H.; Khavari-Nejad, R.A.; Niknam, V.; Razavi, K.; Najafi, F. Effect of penconazole and drought stress on the essential oil composition and gene expression of Mentha pulegium L. (Lamiaceae) at flowering stage. Acta Physiol. Plant. 2014, 36, 1167–1175. [Google Scholar] [CrossRef]
- Al Jaouni, S.; Saleh, A.M.; Wadaan, M.A.M.; Hozzein, W.N.; Selim, S.; AbdElgawad, H. Elevated CO2 induces a global metabolic change in basil (Ocimum basilicum L.) and peppermint (Mentha piperita L.) and improves their biological activity. J. Plant Physiol. 2018, 224, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Erice, G.; Irigoyen, J.J.; Pérez, P.; Martínez-Carrasco, R.; Sánchez-Díaz, M. Effect of elevated CO2, temperature and drought on photosynthesis of nodulated alfalfa during a cutting regrowth cycle. Physiol. Plant. 2006, 126, 458–468. [Google Scholar] [CrossRef]
- Watanabe, C.K.; Sato, S.; Yanagisawa, S.; Uesono, Y.; Terashima, I.; Noguchi, K. Effects of elevated CO2 on levels of primary metabolites and transcripts of genes encoding respiratory enzymes and their diurnal patterns in Arabidopsis thaliana: Possible relationships with respiratory rates. Plant Cell Physiol. 2014, 55, 341–357. [Google Scholar] [CrossRef] [Green Version]
- Hozzein, W.N.; Saleh, A.M.; Habeeb, T.H.; Wadaan, M.A.M.; AbdElgawad, H. CO2 treatment improves the hypocholesterolemic and antioxidant properties of fenugreek seeds. Food Chem. 2020, 308, 1256–1261. [Google Scholar] [CrossRef]
- Cruzat, V.; Rogero, M.M.; Keane, K.N.; Curi, R.; Newsholme, P. Glutamine: Metabolism and immune function, supplementation and clinical translation. Nutrients 2018, 10, 1564. [Google Scholar] [CrossRef] [Green Version]
- Kanunnikova, N.P. Role of brain glutamic acid metabolism changes in neurodegenerative pathologies. J. Biol. Earth Sci. 2012, 2, 1–10. [Google Scholar]
- Lomelino, C.L.; Andring, J.T.; McKenna, R.; Kilberg, M.S. Asparagine synthetase: Function, structure, and role in disease. J. Biol. Chem. 2017, 19952–19958. [Google Scholar] [CrossRef] [Green Version]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef]
- Li, P.; Li, H.; Zong, Y.; Li, F.Y.; Han, Y.; Hao, X. Photosynthesis and metabolite responses of Isatisindigotica Fortune to elevated [CO2]. Crop J. 2017, 5, 345–353. [Google Scholar] [CrossRef]
- Krumbein, A.; Kläring, H.P.; Schonhof, I.; Schreiner, M. Atmospheric carbon dioxide changes photochemical activity, soluble sugars and volatile levels in broccoli (Brassica oleracea var. italica). J. Agric. food Chem. 2010, 58, 3747–3752. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Ashkani, S.; Baghdadi, A.; Pazoki, A.; Jaafar, H.Z.; Rahmat, A. Improvement in flavonoids and phenolic acids production and pharmaceutical quality of sweet basil (Ocimum basilicum L.) by Ultraviolet-B irradiation. Molecules 2016, 21, 1203. [Google Scholar] [CrossRef] [Green Version]
- Fedor, M.; Socha, K.; Urban, B.; Soroczyńska, J.; Matyskiela, M.; Borawska, M.H.; Bakunowicz-Łazarczyk, A. Serum concentration of zinc, copper, selenium, manganese, and Cu/Zn ratio in children and adolescents with myopia. Biol. Trace Elem. Res. 2017, 176, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Asensi-Fabado, M.A.; Munné-Bosch, S. Vitamins in plants: Occurrence, biosynthesis and antioxidant function. Trends Plant Sci. 2010, 15, 582–592. [Google Scholar] [CrossRef]
- Asif, M.; Yilmaz, O.; Ozturk, L. Elevated carbon dioxide ameliorates the effect of Zn deficiency and terminal drought on wheat grain yield but compromises nutritional quality. Plant Soil. 2017, 411, 57–67. [Google Scholar] [CrossRef]
- Goswami, N.; Chatterjee, S. Assessment of free radical scavenging potential and oxidative DNA damage preventive activity of Trachyspermum ammi L. (carom) and Foeniculum vulgare Mill. (fennel) seed extracts. BioMed Res. Int. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.Y.; Bunce, J.A.; Maas, J.L. Elevated carbon dioxide increases contents of antioxidant compounds in field-grown strawberries. J. Agric. Food Chem. 2003, 51, 4315–4320. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.Z.; Rahmat, A. Elevated carbon dioxide increases contents of flavonoids and phenolic compounds, and antioxidant activities in Malaysian young ginger (Zingiber officinale Roscoe.) varieties. Molecules 2010, 15, 7907–7922. [Google Scholar] [CrossRef] [Green Version]
- Jaafar, H.Z.; Ibrahim, M.H.; Karimi, E. Phenolics and flavonoids compounds, phenylanine ammonia lyase and antioxidant activity responses to elevated CO2 in Labisia pumila (Myrisinaceae). Molecules 2012, 17, 6331–6347. [Google Scholar] [CrossRef] [Green Version]
- Moein, M.R.; Zomorodian, K.; Pakshir, K.; Yavari, F.; Motamedi, M.; Zarshenas, M.M. Trachyspermum ammi (L.) sprague: Chemical composition of essential oil and antimicrobial activities of respective fractions. J. Evid. Based Complement. Altern. Med. 2015, 20, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Ghasemzadeh, A.; Jaafar, H.Z. Antioxidant potential and anticancer activity of young ginger (Zingiber officinale Roscoe) grown under different CO2 concentration. J. Med. Plants Res. 2011, 5, 3247–3255. [Google Scholar]
- Kumar, M.Y.; Tirpude, R.J.; Maheshwari, D.T.; Bansal, A.; Misra, K. Antioxidant and antimicrobial properties of phenolic rich fraction of Seabuckthorn (Hippophae rhamnoides L.) leaves in vitro. Food Chem. 2013, 141, 3443–3450. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Control Sprouts | eCO2 Sprouts |
|---|---|---|
| Fresh weight (g) | 8.6 ± 0.8 a | 9.7 ± 1.3 a |
| Dry weight (g) | 1.03 ± 0.2 a | 1.96 ± 0.2 b |
| Photosynthesis (µmol CO2 m−2 S−1) | 17.2 ± 0.4 a | 21 ± 1.2 b |
| Respiration (µmol CO2 m−2 S−1) | 0.08 ± 0.01 a | 0.1 ± 0.01 a |
| Parameters | Control Seeds | Control Sprouts | eCO2 Sprouts |
|---|---|---|---|
| Sugars (mg/g FW) | |||
| Glucose | 0.34 ± 0.01 a | 0.49 ± 0 a | 0.64 ± 0.3 ab |
| Fructose | 0.15 ± 0.01 a | 0.17 ± 0 a | 0.3 ± 0.1 a |
| Sucrose | 0.41 ± 0.0 a | 0.85 ± 0.1 b | 1.1 ± 0.03 b |
| Soluble sugars | 2.63 ± 0.07 a | 4.23 ± 0.1 b | 5.1 ± 0.1 bc |
| Starch | 14.05 ± 0.23 a | 20.05 ± 0.4 b | 30.8 ± 1.1 b |
| Total carbohydrates | 28.9 ± 0.9 a | 38.9 ± 1.7 b | 51.4 ± 1.2 b |
| Amino acids (mg/g FW) | |||
| Lysine | 2.1 ± 0.01 a | 2.4 ± 0.06 b | 3.2 ± 0.05 b |
| Histidine | 1.2 ± 0.01 a | 1.5 ± 0.05 a | 1.4 ± 0.04 a |
| Alanine | 5.8 ± 0.9 a | 11.9 ± 0.39 b | 13.4 ± 0.3 b |
| Arginine | 1.0 ± 0.01 a | 1.6 ± 0.08 ab | 2.8 ± 0.06 c |
| Isoleucine | 0.02 ± 0.01 a | 0.11 ± 0.01 b | 0.14 ± 0.01 b |
| Asparagine | 0.6 ± 0.01 a | 0.9 ± 0.03 b | 0.9 ± 0.02 b |
| Ornithine | 0.03 ± 0.01 a | 0.1 ± 0.01 b | 0.2 ± 0.01 b |
| Glycine | 1.2 ± 0.03 a | 1.0 ± 0.05 a | 1.3 ± 0.04 a |
| Phenylalanine | 0.38 ± 0.08 a | 0.4 ± 0.02 a | 0.6 ± 0.02 b |
| Serine | 0.3 ± 0.2 a | 0.6 ± 0.2 b | 1.3 ± 0.1 b |
| Proline | 1.3 ± 0.1 a | 1.3 ± 0.03 a | 1.4 ± 0.3 a |
| Valine | 0.28 ± 0.01 a | 0.37 ± 0.02 b | 0.8 ± 0.02 c |
| Aspartate | 0.04 ± 0 a | 0.03 ± 0 b | 0.7 ± 0 c |
| Cystine | 0.04 ± 0 a | 0.06 ± 0 b | 0.8 ± 0 c |
| Leucine | 0.024 ± 0 a | 0.02 ± 0 b | 0.08 ± 0 c |
| Methionine | 0.005 ± 0 a | 0.015 ± 0 b | 0.02 ± 0 b |
| Threonine | 0.13 ± 0.02 a | 0.15 ± 0 a | 0.19 ± 0.01 a |
| Tyrosine | 0.8 ± 0.04 a | 0.8 ± 0.04 a | 0.7 ± 0.03 a |
| Glutamine | 69.2 ± 2.6 a | 76.2 ± 1.4 b | 88.9 ± 1.2 c |
| Glutamic acid | 24.1 ± 1.2 a | 39.1 ± 1.2 b | 45.1 ± 1 c |
| Organic acids (mg/gFW) | |||
| Oxalic | 2.1 ± 0.23 a | 2.2 ± 0.53 b | 2.19 ± 0.06 b |
| Malic | 1.5 ± 0.4 a | 2.19 ± 0.4 b | 2.76 ± 0.2 bc |
| Isobutyric | 0.7 ± 0.04 a | 1.7 ± 0.04 b | 1.85 ± 0.2 c |
| Fumaric | 0.31 ± 0.2 a | 0.36 ± 0.16 a | 0.37 ± 0.04 b |
| Succinic | 0.71 ± 0.45 a | 1.57 ± 0.4 b | 2.74 ± 0.07 c |
| Citric | 0.17 ± 0.0 a | 1.27 ± 0.1 b | 1.7 ± 0.1 c |
| Essential Oils (mg/g FW) | Control Seeds | Control Sprouts | eCO2 Sprouts |
|---|---|---|---|
| α-Pinene | 0.21 ± 0.01 a | 0.4 ± 0.07 b | 0.5 ± 0.02 b |
| α-Thujene | 2.2 ± 0.1 a | 4.2 ± 0.4 b | 6.2 ± 0.18 c |
| Sabiene | 4.9 ± 0.7 a | 5.3 ± 0.73 a | 5.0 ± 0.18 a |
| β-Myrcene | 3.9 ± 0.8 a | 8 ± 0.8 b | 10.8 ± 0.1 c |
| D-Limonene | 1.6 ± 0.1 a | 1.9 ± 0.2 a | 2.0 ± 0.09 a |
| α-Terpinene | 15.5 ± 1.7 a | 26.5 ± 1.2 b | 31.1 ± 0.4 c |
| Cymene | 24.4 ± 2.11 a | 24.4 ± 2.13 a | 31.1 ± 0.4 b |
| α-Terpinolene | 4.2 ± 0.43 a | 6.8 ± 0.6 b | 7.6 ± 0.1 bc |
| Thymol | 0.76 ± 0.04 ab | 0.67 ± 0.04 a | 0.95 ± 0.01 b |
| Carvacrol | 0.71 ± 0.04 a | 0.71 ± 0.04 a | 0.58 ± 0.02 b |
| α-Thujone | 0.17 ± 0.1 a | 0.15 ± 0.01 a | 0.29 ± 0 b |
| Tricyclene | 0.01 ± 0.0 a | 0.035 ± 0.0 b | 0.046 ± 0.1 b |
| β-Phellandrene | 0.01 ± 0.0 a | 0.03 ± 0.0 a | 0.019 ± 0 b |
| Humuline | 0.0083 ± 0 a | 0.013 ± 0 b | 0.013 ± 0 b |
| Phenolics and Flavonoids (mg/g FW) | Control-Seeds | Control-Sprouts | eCO2-Sprouts |
|---|---|---|---|
| Gallic acid | 0.13 ± 0 a | 0.36 ± 0 b | 0.47 ± 0.01 c |
| Caffeic acid | 0.58 ± 0.01 a | 0.52 ± 0 a | 0.71 ± 0.01 b |
| p-Coumaric acid | 1.72 ± 0.4 a | 1.52 ± 0.32 a | 2.59 ± 0.34 b |
| Chicoric acid | 0.94 ± 0.15 a | 0.94 ± 0.15 a | 1.46 ± 0.16 b |
| Rosmarinic acid | 1.08 ± 0.19 a | 1.08 ± 0.19 a | 1.74 ± 0.21 b |
| Protocatechuic acid | 3.62 ± 0.2 a | 3.62 ± 0.26 a | 4.24 ± 0.3 b |
| Quercetin | 0.039 ± 0 a | 0.09 ± 0 b | 0.1 ± 0 b |
| Naringenin | 0.005 ± 0.01 a | 0.02 ± 0.01 b | 0.05 ± 0.0 c |
| Kaempferol | 0.07 ± 0.01 a | 0.06 ± 0 a | 0.07 ± 0.01 a |
| Luteolin | 0.06 ± 0 a | 0.05 ± 0 a | 0.09 ± 0.01 b |
| Apigenin | 0.03 ± 0 a | 0.11 ± 0 b | 0.11 ± 0.02 b |
| Naringenin | 0.02 ± 0.01 a | 0.02 ± 0 a | 0.02 ± 0 a |
| Rutin | 0.00 ± 0 a | 0.014 ± 0 b | 0.017 ± 0 b |
| Chlorogenic acid | 0.013 ± 0 a | 0.01 ± 0 a | 0.03 ± 0 b |
| Total phenols | 4.2 ± 0.2 a | 8.2 ± 0.9 b | 17.2 ± 0.4 c |
| Total flavonoids | 0.58 ± 0.01 a | 1.15 ± 0 b | 2.6 ± 0.1 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, A.H.A.; Okla, M.K.; Al-amri, S.A.; El-Tayeb, M.A.; Moussa, I.M.; B. Elbadawi, Y.; Adel-Maksoud, M.A.; Ali, F.H.M.; S. Almaary, K.; Selim, S.; et al. Exploratory Assessment to Evaluate Seed Sprouting under Elevated CO2 Revealed Improved Biomass, Physiology, and Nutritional Value of Trachyspermum ammi. Agronomy 2021, 11, 830. https://doi.org/10.3390/agronomy11050830
Hassan AHA, Okla MK, Al-amri SA, El-Tayeb MA, Moussa IM, B. Elbadawi Y, Adel-Maksoud MA, Ali FHM, S. Almaary K, Selim S, et al. Exploratory Assessment to Evaluate Seed Sprouting under Elevated CO2 Revealed Improved Biomass, Physiology, and Nutritional Value of Trachyspermum ammi. Agronomy. 2021; 11(5):830. https://doi.org/10.3390/agronomy11050830
Chicago/Turabian StyleHassan, Abdelrahim H. A., Mohammad K. Okla, Saud A. Al-amri, Mohamed A. El-Tayeb, Ihab M. Moussa, Yahaya B. Elbadawi, Mostafa A. Adel-Maksoud, Fatma H. M. Ali, Khalid S. Almaary, Samy Selim, and et al. 2021. "Exploratory Assessment to Evaluate Seed Sprouting under Elevated CO2 Revealed Improved Biomass, Physiology, and Nutritional Value of Trachyspermum ammi" Agronomy 11, no. 5: 830. https://doi.org/10.3390/agronomy11050830
APA StyleHassan, A. H. A., Okla, M. K., Al-amri, S. A., El-Tayeb, M. A., Moussa, I. M., B. Elbadawi, Y., Adel-Maksoud, M. A., Ali, F. H. M., S. Almaary, K., Selim, S., & AbdElgawad, H. (2021). Exploratory Assessment to Evaluate Seed Sprouting under Elevated CO2 Revealed Improved Biomass, Physiology, and Nutritional Value of Trachyspermum ammi. Agronomy, 11(5), 830. https://doi.org/10.3390/agronomy11050830