
Predicting Seedling Emergence of Three Canarygrass (Phalaris) Species under Semi-Arid Conditions Using Parametric and Non-Parametric Models



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
2.3. Estimation of TT and HTT
2.4. Development of the Parametric Model
2.5. Development of the Non-Parametric Model
2.6. Model Accuracy
2.7. Data Validation
3. Results
3.1. Weather Conditions
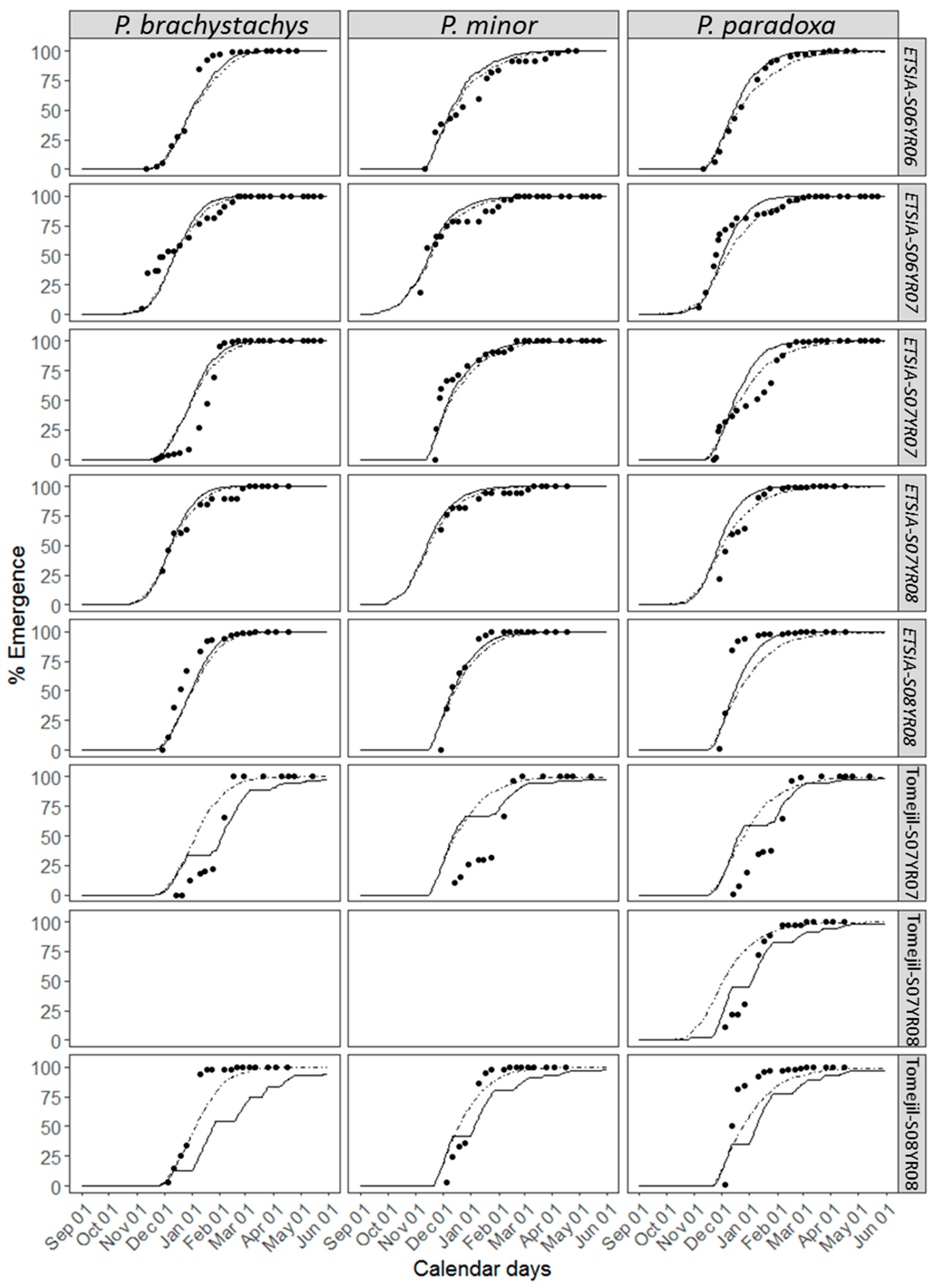
3.2. Description of the Emergence
3.3. Accuracy of the Parametric Model
3.4. Accuracy of the Non-Parametric Model
3.5. Validation of the Models with Independent Data Sets
4. Discussion
4.1. Emergence Pattern
4.2. Threshold Parameters (Tb, To, Tc and Ψb)
4.3. Parametric vs. Non-Parametric Models
4.4. Applicability of the Models
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- García Baudín, J.; Contreras de Vera, A.; Salto, T. Especies adventicias del género Phalaris, en los cereales de invierno de Andalucía Occidental. Boletín Sanid. Veg. Plagas 1982, 8, 185–191. [Google Scholar]
- Damanakis, M. Weed species in wheat fields of Greece-1982, 1983 survey. Zizaniologia 1983, 1, 85–90. [Google Scholar]
- Chhokar, R.S.; Sharma, R.K. Multiple herbicide resistance in littleseed canarygrass (Phalaris minor): A threat to wheat production in India. Weed Biol. Manag. 2008, 8, 112–123. [Google Scholar] [CrossRef]
- Bir, S.S.S.M. Observations on the weed flora of cultivable lands in Punjab- wheat fields in Patiala District. New Bot. 1979, 6, 79–89. [Google Scholar]
- Xu, G.; Shen, S.; Zhang, Y.; Clements, D.R.; Yang, S.; Li, J.; Dong, L.; Zhang, F.; Jin, G.; Gao, Y. Designing Cropping Systems to Improve the Management of the Invasive Weed Phalaris minor Retz. Agronomy 2019, 9, 809. [Google Scholar] [CrossRef] [Green Version]
- USDA PLANTS database. Choice Rev. Online 2006, 43. [CrossRef]
- Gonzalez-Andujar, J.L.; Jimenez-Hidalgo, M.; Garcia-Torres, L.; Saavedra, M. Demography and population dynamic of the arable weed Phalaris brachystachys L. (short-spiked canary grass) in winter wheat. Crop Prot. 2005, 24, 581–584. [Google Scholar] [CrossRef]
- Alcantara, C.; Fuentes-García, M.; Carranza, R.; Sanchez-Gamonoso, M.; Saavedra, M. Aplicaciones de Herbicidas de Postemergencia Temprana y Tardía Contra Alpisteras (Phalaris spp.) en Trigo Duro; Consejería de Agricultura, Pesca y Desarrollo Rural. Instituto de Investigación y Formación Agraria y Pesquera: Cordoba, Spain, 2017. [Google Scholar]
- Ortiz, R.; Contreras, J.M.; Ruiz, A.; Sanz, M.A.; Romero, M.; Gordillo, M.; Taberner, A.; Urbano, J.M.; Urbano Fuentes-Guerra, J.M. Malas hierbas preocupantes en españa. Actas la Soc. Española Malherbologia 2015, 1, 497–503. [Google Scholar]
- Cudney, D.W.; Hill, J.E. The response of wheat grown with three population levels of canarygrass to various herbicide treatments. Proc. West. Soc. Weed Sci. 1979, 55–56. [Google Scholar]
- Jimenez-Hidalgo, M. Incidencia y Evolución de los Alpistes (Phalaris ssp.) En el Trigo (Triticum sp.); Universidad de Córdoba: Andalusia, Spain, 1993. [Google Scholar]
- Afentouli, C.G.; Eleftherohorinos, I.G. Competition between wheat and canarygrass biotypes and their response to herbicides. Weed Sci. 1999, 47, 55–61. [Google Scholar] [CrossRef]
- González-Diaz, L.; Bastida, F.; Gonzalez-Andujar, J.L. Short comunication. Modelling of the population dynamics of Phalaris brachystachys Link under various herbicide control scenarios in a Mediterranean climate. Spanish J. Agric. Res. 2009, 7, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Heap, I. International Survey of Herbicide Resistant Weeds. Available online: http://www.weedscience.org (accessed on 1 May 2020).
- Forcella, F.; Benech-Arnold, R.L.; Sánchez, R.; Ghersa, C.M. Modelling seedling emergence. Field Crops Res. 2000, 67, 123–139. [Google Scholar] [CrossRef]
- Royo-Esnal, A.; Torra, J.; Chantre, G.R. Weed Emergence Models. In Decision Support Systems for Weed Management; Springer: Berlin/Heidelberg, Germany, 2020; pp. 85–116. [Google Scholar]
- Bradford, K.J. Applications of hydrothermal time to quantifying and modeling seed germination and dormancy. Weed Sci. 2002, 50, 248–260. [Google Scholar] [CrossRef]
- Grundy, A.C. Predicting weed emergence: A review of approaches and future challenges. Weed Res. 2003, 43, 1–11. [Google Scholar] [CrossRef]
- Cao, R.; Francisco-Fernández, M.; Anand, A.; Bastida, F.; González-Andújar, J.L. Computing statistical indices for hydrothermal times using weed emergence data. J. Agric. Sci. 2011, 149, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Andujar, J.L.; Francisco-Fernandez, M.; Cao, R.; Reyes, M.; Urbano, J.M.; Forcella, F.; Bastida, F. A comparative study between nonlinear regression and nonparametric approaches for modelling Phalaris paradoxa seedling emergence. Weed Res. 2016, 56, 367–376. [Google Scholar] [CrossRef]
- Mesgaran, M.B.; Mashhadi, H.R.; Alizadeh, H.; Hunt, J.; Young, K.R.; Cousens, R.D. Importance of distribution function selection for hydrothermal time models of seed germination. Weed Res. 2013, 53, 89–101. [Google Scholar] [CrossRef]
- Derakhshan, A.; Gherekhloo, J.; Vidal, R.A.; De Prado, R. Quantitative Description of the Germination of Littleseed Canarygrass (Phalaris minor) in Response to Temperature. Weed Sci. 2014, 62, 250–257. [Google Scholar] [CrossRef]
- Singh, S.; Kirkwood, R.C.; Marshall, G. Biology and control of Phalaris minor Retz. (littleseed canarygrass) in wheat. Crop Prot. 1999, 18, 1–16. [Google Scholar] [CrossRef]
- Sousa-Ortega, C.; Chamber, E.; Urbano, J.M.; Izquierdo, J.; Loureiro, I.; Marí, A.I.; Cordero, F.; Vargas, M.; Saavedra, M.; Lezaun, J.A.; et al. Should emergence models for Lolium rigidum be changed throughout climatic conditions? The case of Spain. Crop Prot. 2020, 128, 105012. [Google Scholar] [CrossRef]
- Sousa-Ortega, C.; Royo-Esnal, A.; DiTommaso, A.; Izquierdo, J.; Loureiro, I.; Marí, A.I.; Cordero, F.; Vargas, M.; Saavedra, M.; Paramio, J.A.; et al. Modeling the Emergence of North African Knapweed (Centaurea diluta), an Increasingly Troublesome Weed in Spain. Weed Sci. 2020, 1–30. [Google Scholar] [CrossRef]
- Saxton, K.E.; Rawls, W.J. Soil water characteristic estimates by texture and organic matter for hydrologic solutions. Soil Sci. Soc. Am. J. 2006, 70, 1569–1578. [Google Scholar] [CrossRef] [Green Version]
- Fuentes-Yagüe, J.; García-Legaspi, G. Técnicas de Riego. Ministerio de Agricultura, Pesca y Alimentación; Ministerio de Agricultrua, pesca y alimentación, Ed.; Mundi-Prensa México: Ciudad Cuauhtémoc, México, 1999; ISBN 968-7462-17-5. [Google Scholar]
- Duong, T. Ks: Kernel density estimation and kernel discriminant analysis for multivariate data in R. J. Stat. Softw. 2007, 21, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Royo-Esnal, A.; Torra, J.; Conesa, J.A.; Recasens, J. Characterisation of emergence of autumn and spring cohorts of Galium spp. in winter cereals. Weed Res. 2010, 50, 572–585. [Google Scholar] [CrossRef] [Green Version]
- Taylor, I.N.; Walker, S.R.; Adkins, S.W. Burial depth and cultivation influence emergence and persistence of Phalaris paradoxa seed in an Australian sub-tropical environment. Weed Res. 2005, 45, 33–40. [Google Scholar] [CrossRef]
- Sousa-Ortega, C.; Royo-Esnal, A.; Loureiro, I.; Marí, A.I.; Lezáun, J.A.; Cordero, F.; Saavedra, M.; Paramio, J.A.; Fernández, J.L.; Torra, J.; et al. Modelling Emergence of Sterile Oat (Avena sterilis spp ludoviciana) Under Semi-arid Conditions. Weed Sci. 2021, 1–29. [Google Scholar] [CrossRef]
- Mesgaran, M.B.; Onofri, A.; Mashhadi, H.R.; Cousens, R.D. Water availability shifts the optimal temperatures for seed germination: A modelling approach. Ecol. Modell. 2017, 351, 87–95. [Google Scholar] [CrossRef]
- Ahmadi, A.; Hosseini, M.; Zeidali, E. Study of ecological characteristics of canary grass (Phalaris minor). Tech. J. Eng. Appl. Sci. 2013, 3, 1835–1840. [Google Scholar]
- Alcantara, C.; Jimenez-Hidalgo, M.J.; Saavedra, M. Responses of Phalaris minor Rezt. and Phalaris brachystachys Link to different levels of soil water availability. Spanish J. Agric. Res. 2010, 8, 1074. [Google Scholar] [CrossRef] [Green Version]
- Taylor, I.N.; Peters, N.C.B.; Adkins, S.W.; Walker, S.R. Germination response of Phalaris paradoxa L. seed to different light quality. Weed Res. 2004, 44, 254–264. [Google Scholar] [CrossRef]
- Ohadi, S.; Rahimian Mashhadi, H.; Tavakkol-Afshari, R.; Beheshtian Mesgaran, M. Modelling the effect of light intensity and duration of exposure on seed germination of Phalaris minor and Poa annua. Weed. Res. 2010, 50, 209–217. [Google Scholar] [CrossRef]
- Izquierdo, J.; González-Andújar, J.L.; Bastida, F.; Lezaún, J.A.; del Arco, M.J.S. A Thermal Time Model to Predict Corn Poppy (Papaver rhoeas) Emergence in Cereal Fields. Weed Sci. 2009, 57, 660–664. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Variation in Seed Dormancy and Germination within and between Individuals and Populations of a Species. In Seeds (Second Edition): Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: New York, NY, USA, 2014; pp. 277–373. ISBN 9780124166776. [Google Scholar]
- García, A.L.; Recasens, J.; Forcella, F.; Torra, J.; Royo-Esnal, A. Hydrothermal Emergence Model for Ripgut Brome (Bromus diandrus). Weed Sci. 2013, 61, 146–153. [Google Scholar] [CrossRef]
- Masin, R.; Loddo, D.; Gasparini, V.; Otto, S.; Zanin, G. Evaluation of Weed Emergence Model AlertInf for Maize in Soybean. Weed Sci. 2014, 62, 360–369. [Google Scholar] [CrossRef]
- Masin, R.; Loddo, D.; Benvenuti, S.; Otto, S.; Zanin, G. Modeling Weed Emergence in Italian Maize Fields. Weed Sci. 2012, 60, 254–259. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Code 1 | Sowing Date | First Relevant Rain | Total Emergence | ||
|---|---|---|---|---|---|---|
| PHABR | PHAMI | PHAPA | ||||
| ETSIA | S06YR06 | 11 November 2005 | 14 November 2005 | 1794 | 44 | 1519 |
| ETSIA | S06YR07 | 13 September 2006 | 43 | 32 | 174 | |
| ETSIA | S07YR07 | 12 November 2006 | 16 November 2006 | 1391 | 62 | 914 |
| ETSIA | S07YR08 | 21 September 2007 | 46 | 38 | 178 | |
| ETSIA | S08YR08 | 15 November 2007 | 20 November 2007 | 95 | 66 | 895 |
| Tomejil | S07YR07 | 15 November 2006 | 16 November 2006 | 55 | 90 | 473 |
| Tomejil | S07YR08 | 21 September 2007 | - | - | 36 | |
| Tomejil | S08YR08 | 20 November 2007 | 22 November 2007 | 190 | 58 | 679 |
| Location | Year | Number of Emerged Seedling | ||
|---|---|---|---|---|
| P. brachystachys | P. minor | P. paradoxa | ||
| Sevilla-Garden | 2016/17 | 12 | 29 | - |
| 2017/18 | - | 48 | 16 | |
| 2018/19 | 17 | 103 | - | |
| ETSIA | 2018/19 | 26 | 24 | 18 |
| Location | Latitude | Longitude | Sowing Date(2019) | Sand (%) | Silt (%) | Clay (%) | Emerged Seedling |
|---|---|---|---|---|---|---|---|
| Burgos | 42.4402 N | 3.7209 W | Sept 18 | 22 | 46 | 32 | 369 |
| ETSIA | 37.3524 N | 5.9392 W | Sept 20 | 38 | 30 | 32 | 242 |
| Guadalcazar | 37.7558 N | 4.9354 W | Oct 17 | 36 | 33 | 30 | 231 |
| Huesca | 42.1277 N | 0.3987 W | Oct 17 | 25 | 40 | 35 | 514 |
| Tomejil | 37.4027 N | 5.5878 W | Sept 23 | 5 | 32 | 62 | 158 |
| Valladolid | 41.7789 N | 4.8752 W | Oct 10 | 62 | 22 | 16 | 507 |
| Location | Month | Temperature (°C) | Precipitation (mm) | ||||
|---|---|---|---|---|---|---|---|
| 2005/06 | 2006/07 | 2007/08 | 2005/06 | 2006/07 | 2007/08 | ||
| ETSIA | September | 22.4 | 23.7 | 23.0 | 0.0 | 37.0 | 42.8 |
| ETSIA | October | 17.7 | 19.6 | 18.8 | 119.2 | 197.8 | 22.6 |
| ETSIA | November | 11.5 | 14.3 | 13.1 | 25.4 | 120.6 | 91.4 |
| ETSIA | December | 9.7 | 8.9 | 9.6 | 29.0 | 43.4 | 15.0 |
| ETSIA | January | 7.1 | 8.2 | 10.8 | 38.0 | 30.4 | 44.8 |
| ETSIA | February | 9.1 | 11.8 | 13.4 | 52.8 | 59.6 | 68.4 |
| ETSIA | March | 13.4 | 13.4 | 14.1 | 63.6 | 12.4 | 20.0 |
| ETSIA | April | 17.0 | 15.3 | 16.8 | 35.2 | 38.0 | 165.4 |
| Average | 13.5 | 14.4 | 14.9 | 363.2 | 539.2 | 470.4 | |
| Tomejil | September | 24.2 | 23.4 | 65.8 | 30.0 | ||
| Tomejil | October | 20.4 | 18.9 | 93.2 | 44.2 | ||
| Tomejil | November | 15.2 | 13.3 | 78.8 | 118.2 | ||
| Tomejil | December | 9.4 | 10.1 | 37.8 | 12.8 | ||
| Tomejil | January | 8.8 | 11.1 | 32.8 | 57.4 | ||
| Tomejil | February | 12.0 | 13.6 | 58.0 | 65.0 | ||
| Tomejil | March | 12.0 | 13.0 | 20.6 | 25.8 | ||
| Tomejil | April | 14.3 | 16.0 | 43.2 | 177.6 | ||
| Average | 14.5 | 14.9 | 430.2 | 531.0 | |||
| Location. | Sowing 1 | P. brachystachys | P. paradoxa | P. minor | |||
|---|---|---|---|---|---|---|---|
| 50% 2 | Emergence 3 Period | 50% 2 | Emergence 3 Period | 50% 2 | Emergence 3 Period | ||
| ETSIA | S06YR06 | Dec. 28 | 44.1 | Dec. 20 | 59.1 | Dec. 20 | 90.2 |
| ETSIA | S06YR07 | Nov. 30 | 89.9 | Nov. 23 | 86.8 | Nov. 11 | 88.8 |
| ETSIA | S07YR07 | Jan. 17 | 31.6 | Jan. 07 | 74.9 | Nov. 26 | 61.6 |
| ETSIA | S07YR08 | Dec. 07 | 88.6 | Dec. 07 | 45.7 | Mar. 18 | 46.7 |
| ETSIA | S08YR08 | Dec. 18 | 42.0 | Dec. 07 | 16.4 | Dec. 10 | 37.8 |
| Tomejil | S07YR07 | Feb. 01 | 48.0 | Jan. 29 | 54.0 | Jan. 30 | 61.8 |
| Tomejil | S07YR08 | - | - | Jan. 02 | 52.9 | - | 36.9 |
| Tomejil | S08YR08 | Dec. 30 | 30.7 | Dec. 11 | 30.4 | Dec. 30 | 36.9 |
| Model | P. brachystachys | P. paradoxa | P. minor | |||
|---|---|---|---|---|---|---|
| K | b | k | b | k | b | |
| TT | 8.264187 | 2.134985 | 5.161581 | 1.401254 | 4.589567 | 1.3303 |
| HTT | 9.231423 | 2.410334 | 6.093273 | 1.747469 | 4.711248 | 1.402724 |
| Location | Sowing | P. brachystachys | P. paradoxa | P. minor | |||
|---|---|---|---|---|---|---|---|
| TT | HTT | TT | HTT | TT | HTT | ||
| ETSIA | S06YR06 | 10.2 | 8.2 | 5.6 | 5.3 | 7.3 | 9.5 |
| ETSIA | S06YR07 | 10.0 | 11.1 | 10.7 | 8.8 | 5.4 | 6.4 |
| ETSIA | S07YR07 | 13.0 | 13.5 | 8.2 | 12.8 | 9.5 | 8.5 |
| ETSIA | S07YR08 | 5.5 | 7.0 | 7.5 | 12.3 | 3.4 | 5.3 |
| ETSIA | S08YR08 | 11.1 | 10.0 | 19.9 | 14.7 | 8.5 | 7.6 |
| Tomejil | S07YR07 | 24.7 | 15.9 | 24.4 | 22.2 | 30.4 | 26.4 |
| Tomejil | S07YR08 | - | - | 22.2 | 13.0 | - | - |
| Tomejil | S08YR08 | 13.7 | 34.8 | 17.3 | 24.5 | 11.9 | 16.5 |
| Average | 12.6 | 14.4 | 14.5 | 14.2 | 10.9 | 11.5 | |
| Location | Sowing | P. brachystachys | P. paradoxa | P. minor | |||
|---|---|---|---|---|---|---|---|
| TT | HTT | TT | HTT | TT | HTT | ||
| ETSIA | S06YR06 | 7.9 | 8.0 | 12.3 | 3.6 | 7.3 | 9.7 |
| ETSIA | S06YR07 | 9.9 | 11.1 | 16.6 | 7.1 | 5.4 | 5.5 |
| ETSIA | S07YR07 | 13.4 | 12.7 | 7.5 | 12.2 | 9.5 | 11.3 |
| ETSIA | S07YR08 | 6.5 | 6.5 | 4.3 | 12.4 | 3.4 | 3.6 |
| ETSIA | S08YR08 | 11.1 | 12.1 | 29.8 | 16.5 | 8.5 | 6.6 |
| Tomejil | S07YR07 | 26.4 | 13.9 | 13.7 | 21.0 | 30.4 | 22.3 |
| Tomejil | S07YR08 | - | - | 15.2 | 13.6 | - | - |
| Tomejil | S08YR08 | 11.4 | 36.5 | 27.1 | 28.3 | 11.9 | 17.8 |
| Average | 12.4 | 14.4 | 15.8 | 14.4 | 10.9 | 11.0 | |
| Weed | Experiment | Parametric Model | Non-Parametric Model | ||
|---|---|---|---|---|---|
| TT | HTT | TT | HTT | ||
| P. brachystachys | ETSIA-2018/19 | 15.3 | 16.3 | 15.7 | 17.1 |
| Sevilla Garden-2016/17 | 28.7 | 28.5 | 27.7 | 29.5 | |
| Sevilla Garden-2018/19 | 12.5 | 12.2 | 11.3 | 12.2 | |
| Average | 18.9 | 19.0 | 18.2 | 19.6 | |
| P. minor | ETSIA-2018/19 | 9.2 | 8.8 | 9.6 | 7.6 |
| Sevilla Garden-2016/17 | 31.9 | 30.5 | 32.7 | 31.0 | |
| Sevilla Garden-2017/18 | 12.7 | 11.2 | 14.3 | 12.3 | |
| Sevilla Garden-2018/19 | 7.5 | 5.8 | 7.6 | 5.9 | |
| Average | 15.4 | 14.1 | 16.1 | 14.2 | |
| P. paradoxa | Burgos-2019/20 | 10.6 | 6.8 | 16.9 | 4.2 |
| ETSIA-2018/19 | 12.3 | 11.6 | 19.7 | 11.2 | |
| ETSIA-2019/20 | 9.9 | 12.4 | 8.5 | 12.1 | |
| Guadalcazar-2019/20 | 14.8 | 17.7 | 11.7 | 15.8 | |
| Huesca-2019/20 | 11.1 | 9.3 | 17.0 | 8.5 | |
| Sevilla Garden-2017/18 | 18.0 | 12.9 | 25.7 | 13.6 | |
| Tomejil-2019/20 | 16.4 | 9.5 | 9.2 | 12.9 | |
| Valladolid-2019/20 | 14.6 | 8.6 | 20.2 | 6.3 | |
| Average | 13.5 | 11.1 | 16.1 | 10.6 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa-Ortega, C.; Royo-Esnal, A.; Urbano, J.M. Predicting Seedling Emergence of Three Canarygrass (Phalaris) Species under Semi-Arid Conditions Using Parametric and Non-Parametric Models. Agronomy 2021, 11, 893. https://doi.org/10.3390/agronomy11050893
Sousa-Ortega C, Royo-Esnal A, Urbano JM. Predicting Seedling Emergence of Three Canarygrass (Phalaris) Species under Semi-Arid Conditions Using Parametric and Non-Parametric Models. Agronomy. 2021; 11(5):893. https://doi.org/10.3390/agronomy11050893
Chicago/Turabian StyleSousa-Ortega, Carlos, Aritz Royo-Esnal, and José María Urbano. 2021. "Predicting Seedling Emergence of Three Canarygrass (Phalaris) Species under Semi-Arid Conditions Using Parametric and Non-Parametric Models" Agronomy 11, no. 5: 893. https://doi.org/10.3390/agronomy11050893
APA StyleSousa-Ortega, C., Royo-Esnal, A., & Urbano, J. M. (2021). Predicting Seedling Emergence of Three Canarygrass (Phalaris) Species under Semi-Arid Conditions Using Parametric and Non-Parametric Models. Agronomy, 11(5), 893. https://doi.org/10.3390/agronomy11050893