
Physiological Properties and Molecular Regulation in Different Edamame Cultivars under Drought Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Condition
2.2. Drought Treatment
2.3. Trait Investigation
2.4. Quantitative Reverse-Transcription Polymerase Chain Reaction
2.5. Statistical Analysis
3. Results
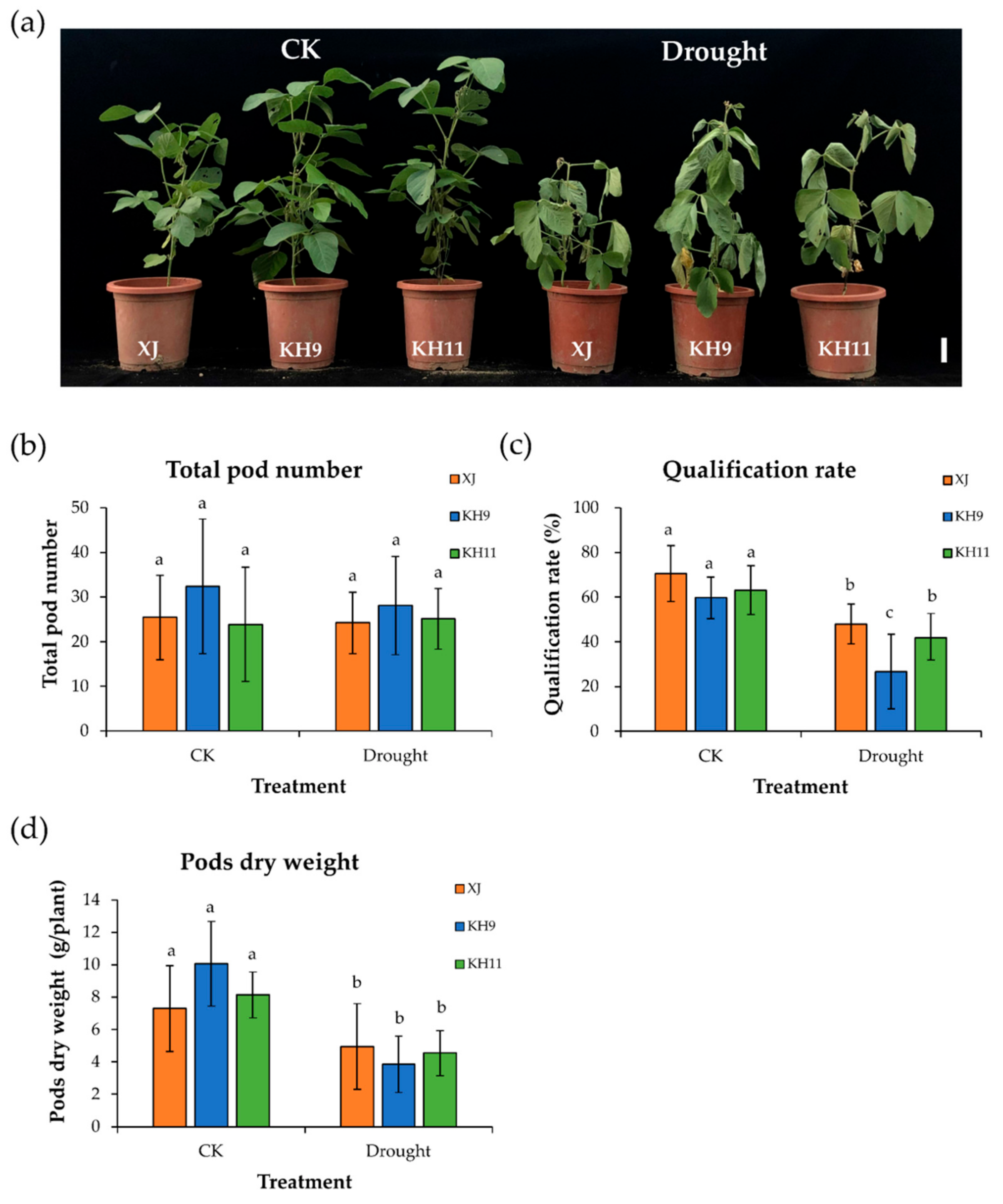
3.1. Phenotypes and Growth Rates of the Edamame Cultivars at Different Developmental Stages
3.2. Pod Seed Numbers and Yields Were Different in the Three Cultivars
3.3. Drought Stress Decreased the Yield in Edamame
3.4. Effect of Drought Stress on the Expression Level of Genes Involved in ABA Biosynthesis and the ABA Signaling Pathway
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Ogles, C.; Guertal, E.; Weaver, D. Edamame cultivar evaluation in central Alabama. J. Agron. 2016, 108, 2371–2378. [Google Scholar] [CrossRef]
- Zarkadas, C.G.; Gagnon, C.; Gleddie, S.; Khanizadeh, S.; Cober, E.R.; Guillemette, R.J. Assessment of the protein quality of fourteen soybean [Glycine max (L.) Merr.] cultivars using amino acid analysis and two-dimensional electrophoresis. Food Res. Int. 2007, 40, 129–146. [Google Scholar] [CrossRef]
- Shiu, M.S.; Shyu, Y.T.; Wu, S.J. Flooding stress and high-pressure treatment enhance the GABA content of the vegetable soybean (Glycine max Merr.). Agriculture 2020, 10, 175. [Google Scholar] [CrossRef]
- Stone, M.S.; Martyn, L.; Weaver, C.M. Potassium intake, bioavailability, hypertension, and glucose control. Nutrients 2016, 8, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirabayashi, Y.; Mahendran, R.; Koirala, S.; Konoshima, L.; Yamazaki, D.; Watanabe, S.; Kim, H.; Kanae, S. Global flood risk under climate change. Nat. Clim. Chang. 2013, 3, 816–821. [Google Scholar] [CrossRef]
- Noorka, I.R.; Tabasum, S. Dose-response behaviour of water scarcity towards genetical and morphological traits in spring wheat (tricticum aestivum l.). Pak. J. Bot. 2015, 47, 1225–1230. [Google Scholar]
- Burnett, S.E.; Pennisi, S.V.; Thomas, P.A.; van Iersel, M.W. Controlled drought affects morphology and anatomy of Salvia splendens. J. Am. Soc. Hortic. Sci. 2005, 130, 775–781. [Google Scholar] [CrossRef] [Green Version]
- Anjum, F.; Yaseen, M.; Rasul, E.; Wahid, A.; Anjum, S. Water stress in barley (Hordeum vulgare L.). II. Effect on chemical composition and chlorophyll contents. Pak. J. Agric. Sci. 2003, 40, 45–49. [Google Scholar]
- Luo, H.; Zhang, Y.; Zhang, W. Effects of water stress and rewatering on photosynthesis, root activity, and yield of cotton with drip irrigation under mulch. Photosynthetica 2016, 54, 65–73. [Google Scholar] [CrossRef]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Zhao, C.X. Water-deficit stress-induced anatomical changes in higher plants. C. R. Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef]
- Osmolovskaya, N.; Shumilina, J.; Kim, A.; Didio, A.; Grishina, T.; Bilova, T.; Keltsieva, O.A.; Zhukov, V.; Tikhonovich, I.; Tarakhovskaya, E.; et al. Methodology of drought stress research: Experimental setup and physiological characterization. Int. J. Mol. Sci. 2018, 19, 4089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raman, A.; Verulkar, S.; Mandal, N.; Variar, M.; Shukla, V.; Dwivedi, J.; Singh, B.; Singh, O.; Swain, P.; Mall, A.; et al. Drought yield index to select high yielding rice lines under different drought stress severities. Rice 2012, 5, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Prasad, S.; Yadav, V.; Kumar, A.; Jaiswal, B.; Kumar, A.; Khan, N.A.; Dwivedi, D.K. Effect of drought stress on yield and yield components of rice (Oryza sativa L.) genotypes. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2752–2759. [Google Scholar]
- Ebrahimian, E.; Seyyedi, S.M.; Bybordi, A.; Damalas, C.A. Seed yield and oil quality of sunflower, safflower, and sesame under different levels of irrigation water availability. Agric. Water Manag. 2019, 218, 149–157. [Google Scholar] [CrossRef]
- Jumrani, K.; Bhatia, V.S. Impact of combined stress of high temperature and water deficit on growth and seed yield of soybean. Physiol. Mol. Biol. Plants 2018, 24, 37–50. [Google Scholar] [CrossRef]
- Kadam, N.N.; Struik, P.C.; Rebolledo, M.C.; Yin, X.; Jagadish, S.K. Genome-wide association reveals novel genomic loci controlling rice grain yield and its component traits under water-deficit stress during the reproductive stage. J. Exp. Bot. 2018, 69, 4017–4032. [Google Scholar] [CrossRef] [PubMed]
- Sourour, A.; Afef, O.; Mounir, R.; Mongi, B.Y. A review: Morphological, physiological, biochemical and molecular plant responses to water deficit stress. Int. J. Eng. Sci. 2017, 6, 1–4. [Google Scholar] [CrossRef]
- Krishnan, S.; Merewitz, E.B. Drought stress and Trinexapac-ethyl modify phytohormone content within Kentucky bluegrass leaves. J. Plant Growth Regul. 2015, 34, 1–12. [Google Scholar] [CrossRef]
- Spollen, W.G.; LeNoble, M.E.; Samuels, T.D.; Bernstein, N.; Sharp, R.E. Abscisic acid accumulation maintains maize primary root elongation at low water potentials by restricting ethylene production. Plant Physiol. 2000, 122, 967–976. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Jensen, C.R.; Andersen, M.N. Hydraulic and chemical signals in the control of leaf expansion and stomatal conductance in soybean exposed to drought stress. Funct. Plant Biol. 2003, 30, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Setter, T.L.; Flannigan, B.A. Water deficit inhibits cell division and expression of transcripts involved in cell proliferation and endoreduplication in maize endosperm. J. Exp. Bot. 2001, 52, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Trouverie, J.; Thévenot, C.; Rocher, J.P.; Sotta, B.; Prioul, J.L. The role of abscisic acid in the response of a specific vacuolar invertase to water stress in the adult maize leaf. J. Exp. Bot. 2003, 54, 2177–2186. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.I.; Hong, J.H.; Ha, J.O.; Kang, J.Y.; Kim, S.Y. ABFs, a family of ABA-responsive element binding factors. J. Biol. Chem. 2000, 275, 1723–1730. [Google Scholar] [CrossRef] [Green Version]
- de Ollas, C.; Dodd, I.C. Physiological impacts of ABA-JA interactions under water-limitation. Plant. Mol. Biol. 2016, 91, 641–650. [Google Scholar] [CrossRef]
- Zhao, W.; Cheng, Y.H.; Zhang, C.; Shen, X.J.; You, Q.B.; Guo, W.; Li, X.; Song, X.J.; Zhou, X.A.; Jiao, Y.Q. Genome-wide identification and characterization of the GmSnRK2 family in soybean. Int. J. Mol. Sci. 2017, 18, 1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, T.; Park, Y.; Hwang, I. Abscisic acid: Biosynthesis, inactivation, homoeostasis and signalling. Essays Biochem. 2015, 58, 29–48. [Google Scholar] [PubMed]
- Hoeft, R.G.; Aldrich, S.R.; Nafziger, E.D.; Johnson, R.R. Modern Corn and Soybean Production; MCSP Publications: Savoy, IL, USA, 2000. [Google Scholar]
- Manavalan, L.P.; Guttikonda, S.K.; Phan Tran, L.S.; Nguyen, H.T. Physiological and molecular approaches to improve drought resistance in soybean. Plant Cell Physiol. 2009, 50, 1260–1276. [Google Scholar] [CrossRef] [Green Version]
- Todaka, D.; Takahashi, F.; Yamaguchi-Shinozaki, K.; Shinozaki, K. ABA-responsive gene expression in response to drought stress: Cellular regulation and long-distance signaling. Adv. Bot. Res. 2019, 92, 83–113. [Google Scholar]
- Tuteja, N. Abscisic acid and abiotic stress signaling. Plant Signal. Behav. 2007, 2, 135–138. [Google Scholar] [PubMed] [Green Version]
- Huang, G.T.; Ma, S.L.; Bai, L.P.; Zhang, L.; Ma, H.; Jia, P.; Liu, J.; Zhong, M.; Guo, Z.F. Signal transduction during cold, salt, and drought stresses in plants. Mol. Biol. Rep. 2011, 39, 969–987. [Google Scholar]
- Singh, D.; Laxmi, A. Transcriptional regulation of drought response: A tortuous network of transcriptional factors. Front. Plant Sci. 2015, 6, 895. [Google Scholar] [CrossRef] [Green Version]
- Palada, M.; Ma, C. Performance of vegetable soybean cultivars under organic crop management system. In SEAVEG 2012 High. Value Vegetables in Southeast. Asia: Production, Supply and Demand; Holmer, R., Linwattana, G., Nath, P., Keatinge, J.D.H., Eds.; AVRDC–The World Vegetable Center: Tainan, Taiwan, 2013; p. 181. [Google Scholar]
- Routray, W.; Rayaguru, K. 2-Acetyl-1-pyrroline: A key aroma component of aromatic rice and other food products. Food Rev. Int. 2018, 34, 539–565. [Google Scholar]
- Chen, G.; Wiatrak, P. Soybean development and yield are influenced by planting date and environmental conditions in the southeastern coastal plain, United States. J. Agron. 2010, 102, 1731–1737. [Google Scholar] [CrossRef]
- Wei, Y.; Jin, J.; Jiang, S.; Ning, S.; Liu, L. Quantitative response of soybean development and yield to drought stress during different growth stages in the Huaibei Plain, China. Agronomy 2018, 8, 97. [Google Scholar] [CrossRef] [Green Version]
- Mishra, V.; Cherkauer, K.A. Retrospective droughts in the crop growing season: Implications to corn and soybean yield in the Midwestern United States. Agric. Forest Meteorol. 2010, 150, 1030–1045. [Google Scholar] [CrossRef]
- Sadeghipour, O.; Abbasi, S. Soybean response to drought and seed inoculation. World Appl. Sci. J. 2012, 17, 55–60. [Google Scholar]
- Thao, N.P.; Tran, L.-S.P. Potentials toward genetic engineering of drought-tolerant soybean. Crit. Rev. Biotechnol. 2012, 32, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Andersen, M.N.; Jensen, C.R. Loss of pod set caused by drought stress is associated with water status and ABA content of reproductive structures in soybean. Funct. Plant Biol. 2003, 30, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Maleki, A.; Naderi, A.; Naseri, R.; Fathi, A.; Bahamin, S.; Maleki, R. Physiological performance of soybean cultivars under drought stress. Bull. Env. Pharmacol. Life Sci. 2013, 2, 38–44. [Google Scholar]
- Liu, F.; Jensen, C.R.; Andersen, M.N. Pod set related to photosynthetic rate and endogenous ABA in soybeans subjected to different water regimes and exogenous ABA and BA at early reproductive stages. Ann. Bot. 2004, 94, 405–411. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Total Number of Pods | Qualified Rate (%) | Shelling Percentage (%) | Total Pods Dry Weight (g/plant) | Qualified Pod Dry Weight (g/plant) | |
|---|---|---|---|---|---|---|
| Year | ||||||
| 2017 | XJ | 26.20 ± 3.97 b | 59.69 ± 9.61 a | 56.15 ± 8.28 a | 16.70 ± 4.95 b | 12.20 ± 1.86 b |
| KH9 | 26.06 ± 4.97 b | 53.63 ± 12.64 a | 54.37 ± 5.41 a | 25.16 ± 3.71 a | 14.66 ± 4.67 a | |
| KH11 | 32.06 ± 5.04 a | 50.14 ± 10.08 a | 58.87 ± 4.79 a | 22.62 ± 5.53 a | 16.14 ± 3.85 a | |
| 2018 | XJ | 31.06 ± 6.26 a | 49.87 ± 8.27 b | 49.42 ± 7.34 a | 18.21 ± 2.07 b | 10.32 ± 2.87 b |
| KH9 | 25.88 ± 6.82 b | 60.10 ± 11.57 a | 48.73 ± 12.29 a | 24.76 ± 4.88 a | 14.77 ± 4.43 a | |
| KH11 | 32.13 ± 5.37 a | 58.61 ± 8.50 a | 50.19 ± 11.59 a | 23.94 ± 4.07 a | 15.18 ± 3.44 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.-T.; Lu, C.-T.; Tzen, J.T.C.; Yang, C.-Y. Physiological Properties and Molecular Regulation in Different Edamame Cultivars under Drought Stress. Agronomy 2021, 11, 939. https://doi.org/10.3390/agronomy11050939
Chen C-T, Lu C-T, Tzen JTC, Yang C-Y. Physiological Properties and Molecular Regulation in Different Edamame Cultivars under Drought Stress. Agronomy. 2021; 11(5):939. https://doi.org/10.3390/agronomy11050939
Chicago/Turabian StyleChen, Chung-Tse, Chun-Tang Lu, Jason T. C. Tzen, and Chin-Ying Yang. 2021. "Physiological Properties and Molecular Regulation in Different Edamame Cultivars under Drought Stress" Agronomy 11, no. 5: 939. https://doi.org/10.3390/agronomy11050939
APA StyleChen, C. -T., Lu, C. -T., Tzen, J. T. C., & Yang, C. -Y. (2021). Physiological Properties and Molecular Regulation in Different Edamame Cultivars under Drought Stress. Agronomy, 11(5), 939. https://doi.org/10.3390/agronomy11050939