
Effect of Exogenously Applied Methyl Jasmonate on Yield and Quality of Salt-Stressed Hydroponically Grown Sea Fennel (Crithmum maritimum L.)

 , , ,
, , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Cultivation and Experiment Design
2.2. Fresh, Dry Weights of Shoots and Root Growth Parameters
2.3. Mineral Analysis
2.4. Fatty Acids
2.5. Chlorophylls and Carotenoids
2.6. Total Flavonoids Content
2.7. Total Phenolic Content and Antioxidant Capacity
2.8. Statistics
3. Results
3.1. Biomass and Root Growth Parameters
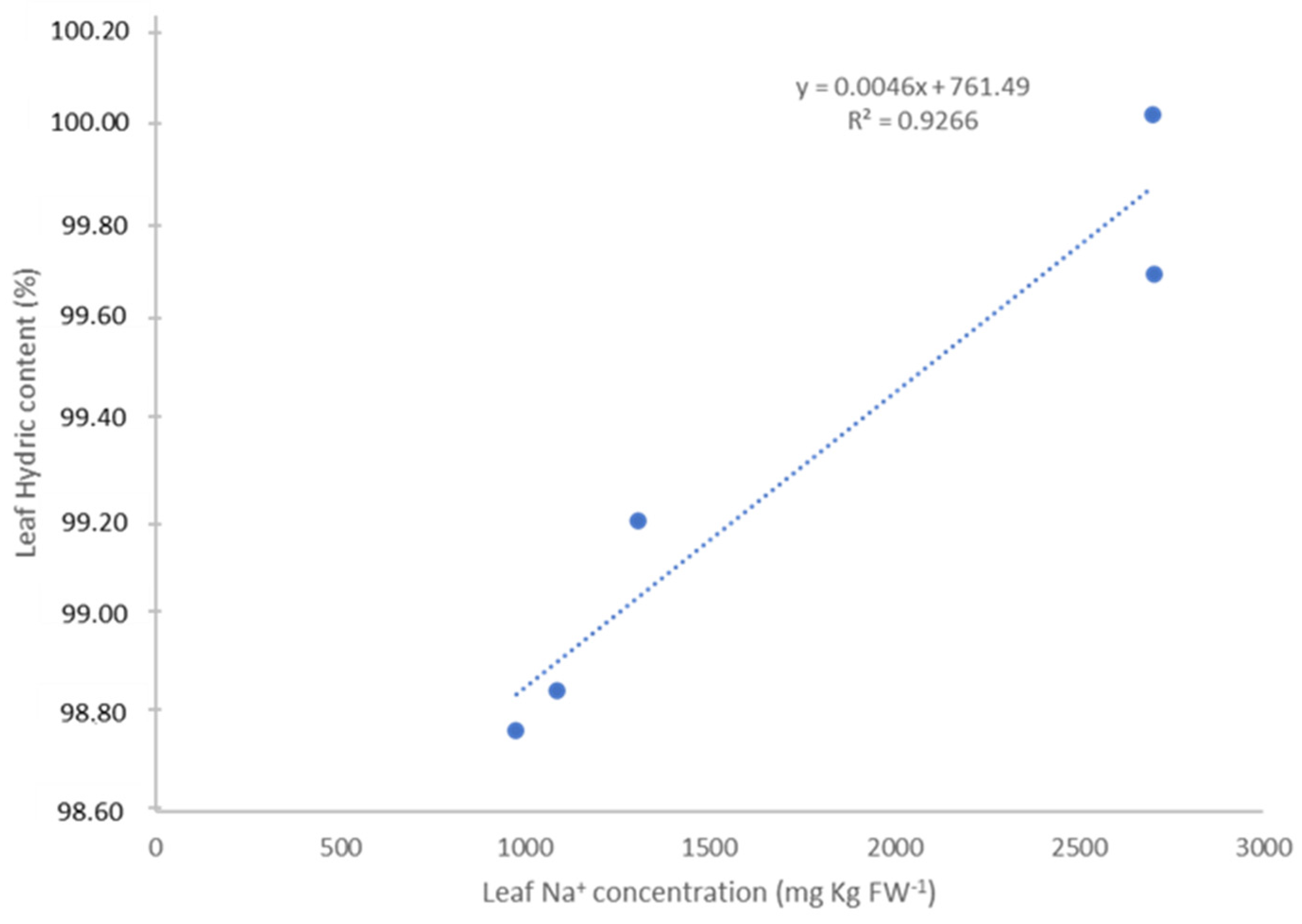
3.2. Ion Content
3.3. Fatty Acids
3.4. Total Phenolic Compounds and Total Flavonoids
3.5. Chlorophylls and Carotenoids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Verzeletti, C.; Maes, L.; Santinello, M.; Vereecken, C.A. Food-related family lifestyle associated with fruit and vegetable consumption among young adolescents in Belgium Flanders and the Veneto Region of Italy. Appetite 2010, 54, 394–397. [Google Scholar] [CrossRef]
- Grosso, G.; Marventano, S.; Nolfo, F.; Rametta, S.; Bandini, L.; Ferranti, R.; Bonomo, M.C.; Matalone, M.; Galvano, F.; Mistretta, A. Personal eating, lifestyle, and family-related behaviors correlate with fruit and vegetable consumption in adolescents living in Sicily, Southern Italy. Int. J. Vitam. Nutr. Res. 2013, 83, 355–366. [Google Scholar] [CrossRef]
- Ngozika, E.; Obeagu, E.A. Review on Fast Foods and Family Lifestyle. Int. J. Curr. Res. Biol. Med. 2018, 3, 26–30. [Google Scholar] [CrossRef]
- Da Silva Santos, J.M. Minimally Processed Baby Leaf Vegetables: Phytonutrient Characterization and Nutritional Stability. Ph.D. Thesis, Universidade do Porto, Porto, Portugal, 2014; p. 173. [Google Scholar]
- Saini, R.K.; Ko, E.Y.; Keum, Y.S. Minimally processed ready-to-eat baby-leaf vegetables: Production, processing, storage, microbial safety and nutritional potential. Food Rev. Int. 2017, 33, 644–663. [Google Scholar] [CrossRef]
- Gil, M.I.; Tudela, J.A.; Martínez-Sánchez, A.; Luna, M.C. Harvest maturity indicators of leafy vegetables. Stewart Postharvest Rev. 2012, 8, 1–9. [Google Scholar] [CrossRef]
- Aires, A.; Marques, E.; Carvalho, R.; Rosa, E.A.S.; Saavedra, M.J. Evaluation of biological value and appraisal of polyphenols and glucosinolates from organic baby-leaf salads as antioxidants and antimicrobials against ımportant human pathogenic bacteria. Molecules 2013, 18, 4651–4668. [Google Scholar] [CrossRef] [PubMed]
- Aires, A.; Carvalho, R.; Rosa, E.A.S.; Saavedra, M.J. Phytochemical characterization and antioxidant properties of baby-leaf watercress produced under organic production system. CyTA J. Food 2013, 11, 343–351. [Google Scholar] [CrossRef]
- Santos, J.; Oliva-Teles, M.T.; Delerue-Matos, C.; Oliveira, M.B.P.P. Multi-elemental analysis of ready-to-eat “baby leaf” vegetables using microwave digestion and high-resolution continuum source atomic absorption spectrometry. Food Chem. 2014, 151, 311–316. [Google Scholar] [CrossRef]
- Sundriyal, M.; Sundriyal, R.C.; Url, S. Wild edible plants of the Sikkim Himalaya: Nutritive values of selected species. Econ. Bot. 2001, 55, 377–390. [Google Scholar] [CrossRef]
- Giménez, A.; Fernández, J.A.; Pascual, J.A.; Ros, M.; López-Serrano, M.; Egea-Gilabert, C. An Agroindustrial Compost as Alternative to Peat for Production of Baby Leaf Red Lettuce in a Floating System. Sci. Hortic. 2019, 246, 907–915. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Svecova, E.; Rea, E.; Lucini, L. Effects of saline stress on mineral composition, phenolic acids and flavonoids in leaves of artichoke and cardoon genotypes grown in floating system. J. Sci. Food Agric. 2013, 93, 1119–1127. [Google Scholar] [CrossRef]
- Borgognone, D.; Cardarelli, M.; Lucini, L.; Colla, G. Does CaCl2 play a role in improving biomass yield and quality of cardoon grown in a floating system under saline conditions? HortScience 2014, 49, 1523–1528. [Google Scholar] [CrossRef]
- Rufí-Salís, M.; Parada, F.; Arcas-Pliz, V.; Petit-Boix, A.; Villalba, G.; Gabarrell, X. Closed-loop crop cascade to optimizze nutrient flows and grow low-impact vegetables in cites. Front. Plant Sci. 2020, 11, 566550. [Google Scholar] [CrossRef] [PubMed]
- D’Imperio, M.; Renna, M.; Cardinali, A.; Buttaro, D.; Serio, F.; Santamaria, P. Calcium biofortification and Bioaccessibility in Soilless “Baby Leaf” Vegetable Production. Food Chem. 2016, 213, 149–156. [Google Scholar] [CrossRef]
- Renna, M. Reviewing the Prospects of Sea Fennel (Crithmum maritimum L.) as Emerging Vegetable Crop. Plants 2018, 7, 92. [Google Scholar] [CrossRef] [Green Version]
- Riddle, J. Folk Tradition and Folk Medicine: Recognition of Drugs in Classical Antiquity. Pharm. Hist. 2013, 55, 64–87. [Google Scholar] [PubMed]
- Mekinić, I.G.; Blažević, I.; Mudnić, I.; Burčul, F.; Grga, M.; Skroza, D.; Jerčić, I.; Ljubenkov, I.; Boban, M.; Miloš, M.; et al. Sea fennel (Crithmum maritimum L.): Phytochemical profile, antioxidative, cholinesterase inhibitory and vasodilatory activity. J. Food Sci. Technol. 2016, 53, 3104–3112. [Google Scholar] [CrossRef] [Green Version]
- Pavela, R.; Maggi, F.; Lupidi, G.; Cianfaglione, K.; Dauvergne, X.; Bruno, M.; Benelli, G. Efficacy of sea fennel (Crithmum maritimum L., Apiaceae) essential oils against Culex quinquefasciatus Say and Spodoptera littoralis (Boisd.). Ind. Crop. Prod. 2017, 109, 603–610. [Google Scholar] [CrossRef]
- Franke, W. Vitamin C in sea fennel (Crithmum maritimum), an edible wild plant. Econ. Bot. 1982, 36, 163–165. [Google Scholar] [CrossRef]
- Males, Z.; Zuntar, I.; Nigovic, B.; Plazibat, M.; Vundac, V.B. Quantitative analysis of the polyphenols of the aerial parts of rock samphire—Crithmum maritimum L. Acta Pharm. 2003, 53, 139–144. [Google Scholar] [PubMed]
- Meot-Duros, L.; Magné, C. Antioxidant activity and phenol content of Crithmum maritimum L. leaves. Plant Physiol. Biochem. 2009, 47, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Jallali, I.; Megdiche, W.; M’Hamdi, B.; Oueslati, S.; Smaoui, A.; Abdelly, C.; Ksouri, R. Changes in phenolic composition and antioxidant activities of the edible halophyte Crithmum maritimum L. with physiological stage and extraction method. Acta Physiol. Plant. 2012, 34, 1451–1459. [Google Scholar] [CrossRef]
- Bartnik, M.; Wierzchowska-Renke, K.; Głowniak, P.; Głowniak, K. Phenolic acids in Crithmum maritimum L. (Apiaceae) after Tytanit fertilization. Acta Soc. Bot. Pol. 2017, 86, 3560. [Google Scholar] [CrossRef] [Green Version]
- Zafeiropoulou, V.; Tomou, E.M.; Ioannidou, O.; Karioti, A.; Skaltsa, H. Sea fennel: Phytochemical analysis of Greek wild and cultivated Crithmum maritimum L. populations, based on HPLC-PDA-MS and NMR methods. J. Pharmacogn. Phytochem. 2020, 9, 998–1004. [Google Scholar] [CrossRef] [Green Version]
- Nabet, N.; Boudries, H.; Chougui, N.; Loupassaki, S.; Souagui, S.; Burló, F.; Hernández, F.; Carbonell-Barrachina, A.A.; Madani, K.; Larbat, R. Biological activities and secondary compound composition from Crithmum maritimum aerial parts. Int. J. Food Prop. 2017, 20, 1843–1855. [Google Scholar] [CrossRef] [Green Version]
- Marchioni-Ortu, A.; Bocchieri, E. A study of the germination responses of a Sardinian population of sea fennel (Crithmum maritimum). Can. J. Bot. 1984, 62, 1832–1835. [Google Scholar] [CrossRef]
- Atia, A.; Ben Hamed, K.; Debez, A.; Abdelly, C. Salt and seawater effects on the germination of Crithmum maritimum. In Biosalite Agriculture and Salinity Tolerance in Plant; Birkhäuser: Basel, Switzerland, 2006; pp. 29–33. [Google Scholar] [CrossRef]
- Atia, A.; Debez, A.; Barhoumi, Z.; Smaoui, A.; Abdelly, C. ABA, GA3, and nitrate may control seed germination of Crithmum maritimum (Apiaceae) under saline conditions. Comptes Rendus Biol. 2009, 332, 704–710. [Google Scholar] [CrossRef]
- Meot-Duros, L.; Magné, C. Effect of salinity and chemical factors on seed germination in the halophyte Crithmum maritimum L. Plant Soil 2008, 313, 83–87. [Google Scholar] [CrossRef]
- Nimac, A.; Lazarevic, B.; Petek, M.; Vidak, M.; Satovic, Z.; Carovic-Stanko, K. Effects of Salinity and Seed Priming on Germination of Sea Fennel (Crithmum maritimum L.). Agric. Conspec. Sci. 2018, 83, 181–185. [Google Scholar]
- Montesano, F.F.; Gattulo, C.E.; Parente, A.; Terzano, R.; Renna, M. Cultivation of Potted Sea Fennel, an Emerging Mediterranean Halophyte, Using a Renewable Seaweed-Based Material as a Peat Substitute. Agriculture 2018, 8, 96. [Google Scholar] [CrossRef] [Green Version]
- Grigoriadou, K.; Maloupa, E. Micropropagation and salt tolerance of in vitro grown Crithmum maritimum L. Plant Cell Tissue Organ Cult. 2008, 94, 209–217. [Google Scholar] [CrossRef]
- Ben Hamed, K.; Castagna, A.; Salem, E.; Ranieri, A.; Abdelly, C. Sea Fennel (Crithmum maritimum L.) under Salinity Conditions: A Comparison of Leaf and Root Antioxidant Responses. Plant Growth Regul. 2007, 53, 185–194. [Google Scholar] [CrossRef]
- Amor, N.B.; Hamed, K.B.; Debez, A.; Grignon, C.; Abdelly, C. Physiological and antioxidant responses of the perennial halophyte Crithmum maritimum to salinity. Plant Sci. 2005, 168, 889–899. [Google Scholar] [CrossRef]
- Zarrouk, M.; El Almi, H.; Ben Youssef, N.; Sleimi, N.; Smaoui, A.; Ben Miled, D.; Abdelly, C. Lipid composition of seeds of local halophytes: Cakile maritima, Zygophyllum album and Crithmum maritimum. In Cash Crop Halophytes: Recent Studies: Tasks for Vegetation Science; Lieth, H., Mochtchenko, M., Eds.; Springer: Dordrecht, The Netherlands, 2003; Volume 38. [Google Scholar] [CrossRef]
- Bahmani, K.; Darbandi, A.I.; Alfekaiki, D.F.; Sticklen, M. Phytochemical diversity of fennel landraces from various growth types and origins. Agron. Res. 2016, 14, 1530–1547. [Google Scholar]
- Wasternack, C.; Song, S. Jasmonates: Biosynthesis, metabolism, and signaling by proteins activating and repressing transcription. J. Exp. Bot. 2017, 68, 1303–1321. [Google Scholar] [CrossRef]
- Faghih, S.; Ghobadi, C.; Zarei, A. Response of strawberry plant cv. ‘Camarosa’ to Salicylic Acid and Methyl Jasmonate application under salt stress condition. J. Plant Growth Regul. 2017, 36, 651–659. [Google Scholar] [CrossRef]
- Hassini, I.; Martinez-Ballesta, M.C.; Boughanmi, N.; Moreno, D.A.; Carvajal, M. Improvement of broccoli sprouts (Brassica oleracea L. var. italica) growth and quality by KCl seed priming and methyl jasmonate under salinity stress. Sci. Hortic. 2017, 226, 141–151. [Google Scholar] [CrossRef]
- Manan, A.; Ayyub, C.M.; Pervez, M.A.; Ahmad, R. Methyl Jasmonate brings about resistance against salinity stressed tomato plants by altering biochemical and physiological processes. Pak. J. Agric. Sci. 2016, 53, 35–41. [Google Scholar] [CrossRef]
- Pérez-Balibrea, S.; Moreno, D.A.; García-Viguera, C. Improving the Phytochemical Composition of Broccoli Sprouts by Elicitation. Food Chem. 2011, 129, 35–44. [Google Scholar] [CrossRef]
- Xie, Q.; Yan, F.; Hu, Z.; Wei, S.; Lai, J.; Chen, G. Accumulation of anthocyanin and its associated gene expression in purple tumorous stem mustard (Brassica juncea Var. tumida Tsen et Lee) sprouts when exposed to light, dark, sugar, and Methyl Jasmonate. J. Agric. Food Chem. 2019, 67, 856–866. [Google Scholar] [CrossRef]
- Lara, L.J.; Egea-Gilabert, C.; Niñirola, D.; Conesa, E.; Fernández, J.A. Effect of Aeration of the Nutrient Solution on the Growth and Quality of Purslane (Portulaca Oleracea). J. Hortic. Sci. Biotechnol. 2011, 86, 603–610. [Google Scholar] [CrossRef]
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A direct method for fatty acid methyl ester synthesis: Application to wet meat tissues, oils, and feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging avtivity. Food Chem. 2005, 571–577. [Google Scholar] [CrossRef]
- Everette, J.D.; Brayant, Q.M.; Green, A.M.; Abbey, Y.A.; Wangila, G.W.; Walker, R.B. Through study of reactivity of various compound classes toward the Folin-Ciocalteu reagent. J. Agric. Food Chem. 2010, 58, 8139–8144. [Google Scholar] [CrossRef] [Green Version]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Pérez-Tortosa, V.; López-Orenes, A.; Martínez-Pérez, A.; Ferrer, M.A.; Calderón, A.A. Antioxidant activity and rosmarinic acid changes in salicylic acid-treated Thymus membranaceus shoot. Food Chem. 2012, 130, 362–369. [Google Scholar] [CrossRef]
- Aslam, R.; Bostan, N.; Maria, M.; Safdar, W. A Critical Review on Halophytes: Salt Tolerant Plants. J. Med. Plants Res. 2011, 5, 7108–7118. [Google Scholar] [CrossRef]
- Ben Hamed, K.; Debez, A.; Chibani, F.; Abdelly, C. Salt Response of Crithmum maritimum, an Oleagineous Halophyte. Trop. Ecol. 2004, 45, 151–159. [Google Scholar]
- Hamdani, F.; Derridj, A.; Rogers, H.J. Diverse salinity responses in Crithmum maritimum tissues at different salinities over time. J. Soil Sci. Plant Nutr. 2017, 17, 716–734. [Google Scholar] [CrossRef] [Green Version]
- Attaran, F.; Mayor, I.T.; Cruz, J.A.; Rosa, B.A.; Koo, A.J.K.; Chen, J.; Kramer, D.M.; He, S.Y.; Howe, G.A. Temporal dynamics of growth and photosynthesis suppression in response to jasmonate signalling. Plant Physiol. 2014, 165, 1302–1314. [Google Scholar] [CrossRef] [Green Version]
- Shyu, C.; Brutnell, T.P. Growth deference balance in grass biomass production: The role of jasmonates. J. Exp. Bot. 2015, 66, 4165–4176. [Google Scholar] [CrossRef] [Green Version]
- Mahmud, S.; Sharmin, S.; Chowdhury, B.L.D.; Hossain, M.A.; Bhuiyan, M.J.H. Mitigation of Salt Stress in Rice Plant at Germination Stage by Using Methyl Jasmonate. Asian J. Med. Biol. Res. 2016, 2, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Comparot, S.M.; Graham, C.M.; Reid, D.M. Methyl Jasmonate Elicits a Differential Antioxidant Response in Light-and Dark-Grown Canola (Brassica napus) Roots and Shoots. Plant Growth Regul. 2002, 38, 21–30. [Google Scholar] [CrossRef]
- Yuan, F.; Liang, X.; Li, Y.; Yin, S.; Wang, B. Methyl Jasmonate Improves Tolerance to High Salt Stress in the Recretohalophyte Limonium bicolor. Funct. Plant Biol. 2019, 46, 82–92. [Google Scholar] [CrossRef]
- Li, C.; Wang, P.; Menzles, N.; Lombi, E. Effects of methyljasmonate on plant growth and leaf properties. J. Plant Nutr. Soil Sci. 2018, 181, 409–418. [Google Scholar] [CrossRef]
- Hamed, K.B.; Messedi, D.; Ranieri, A.; Abdelly, C. Diversity in the response of two potential halophytes (Batis maritima and Crithmum maritimum) to salt stress. In Biosaline Agriculture and High Salinity Tolerance; Birkhäuser: Basel, Switzland, 2008; pp. 71–80. [Google Scholar] [CrossRef]
- Evans, N.H. Modulation of guard cell plasma membrane potassium currents by methyl jasmonate. Plant Physiol. 2003, 131, 8–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armengaud, P.; Breitling, R.; Amtmann, A. The Potassium-Dependent Transcriptome of Arabidopsis Reveals a Prominent Role of Jasmonic Acid in Nutrient Signaling. Plant Physiol. 2004, 136, 2556–2576. [Google Scholar] [CrossRef] [Green Version]
- Battal, P.; Erez, M.E.; Turker, M.; Berber, I. Molecular and Physiological Changes in Maize (Zea Mays) Induced by Exogenous NAA, ABA and MeJa during Cold Stress. Ann. Bot. Fenn. 2008, 45, 173–185. [Google Scholar] [CrossRef]
- Sánchez-Romera, B.; Ruiz-Lozano, J.M.; Li, G.; Luu, D.T.; Martínez-Ballesta, M.D.C.; Carvajal, M.; Zamarreño, A.M.; García-Mina, J.M.; Maurel, C.; Aroca, R. Enhancement of Root Hydraulic Conductivity by Methyl Jasmonate and the Role of Calcium and Abscisic Acid in This Process. Plant Cell Environ. 2014, 37, 995–1008. [Google Scholar] [CrossRef] [PubMed]
- Djanaguiraman, M.; Pasad, P.V.V. Effects of salinity on ion transport, water relations and oxidative damage. In Ecophysiology and Response of Plants under Salt Stress; Springer: New York, NY, USA, 2012; pp. 89–114. [Google Scholar] [CrossRef]
- Smoleń, S.; Lukasiewicz, A.; Klimek-Chodacka, M.; Baranski, R. Effect of Soil Salinity and Foliar Application of Jasmonic Acid on Mineral Balance of Carrot Plants Tolerant and Sensitive to Salt Stress. Agronomy 2020, 10, 659. [Google Scholar] [CrossRef]
- Manaa, A.; Ben Ahmed, H.; Valot, B.; Bouchet, J.P.; Aschi-Smiti, S.; Causse, M.; Faurobert, M. Salt and Genotype Impact on Plant Physiology and Root Proteome Variations in Tomato. J. Exp. Bot. 2011, 62, 2797–2813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benevenuto, R.F.; Seldal, T.; Hegland, S.J.; Rodriguez-Saona, C.; Kawash, J.; Polashock, J. Transcriptional Profiling of Methyl Jasmonate-Induced Defense Responses in Bilberry (Vaccinium myrtillus L.). BMC Plant Biol. 2019, 19, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brkić, D.; Bošnir, J.; Bevardi, M.; Bošković, A.G.; Miloš, S.; Lasić, D.; Krivohlavek, A.; Racz, A.; Ćuić, A.M.; Trstenjak, N.U. Nitrate in leafy green vegetables and estimated intake. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 31–41. [Google Scholar] [CrossRef]
- Iammarino, M.; Di Taranto, A.; Cristino, M. Monitoring of Nitrites and Nitrates Levels in Leafy Vegetables (Spinach and Lettuce): A Contribution to Risk Assessment. J. Sci. Food Agric. 2014, 94, 773–778. [Google Scholar] [CrossRef]
- Chalbi, N.; Martínez-Ballesta, M.C.; Youssef, N.B.; Carvajal, M. Intrinsic stability of Brassicaceae plasma membrane in relation to changes in proteins and lipids as a response to salinity. J. Plant Physiol. 2015, 175, 148–156. [Google Scholar] [CrossRef]
- Ben Hamed, K.; Ben Youssef, N.; Ranieri, A.; Zarrouk, M.; Abdelly, C. Changes in content and fatty acid profiles of total lipids and sulfolipids in the halophyte Crithmum maritimum under salt stress. J. Plant Physiol. 2005, 162, 599–602. [Google Scholar] [CrossRef] [PubMed]
- Moller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [Green Version]
- Guil-Guerrero, J.L.; Rodrñiguez-García, I. Lipids classes, fatty acids and carotenes of the leaves of six edible wild plants. Eur. Food Res. Technol. 1999, 209, 313–316. [Google Scholar] [CrossRef]
- Deng, G.F.; Lin, X.; Xu, X.R.; Gao, L.L.; Xie, J.F.; Li, H.B. Antioxidant Capacities and Total Phenolic Contents of 56 Vegetables. J. Funct. Foods 2013, 5, 260–266. [Google Scholar] [CrossRef]
- Kim, H.J.; Chen, F.; Wang, X.; Rajapakse, N.C. Effect of Methyl Jasmonate on Secondary Metabolites of Sweet Basil (Ocimum basilicum L.). J. Agric. Food Chem. 2006, 54, 2327–2332. [Google Scholar] [CrossRef] [PubMed]
- Asadi Karam, E.; Asrar, Z.; Keramat, B. Effects of Methyl jasmonate pretreatment on phenolic compounds and PAL activity in Lepidium sativum L. subjected to copper toxicity. J. Plant Proc. Funct. 2014, 2, 89–96. [Google Scholar]
- Del Amor, F.; Cuadra-Crespo, P. Alleviation of salinity stress in broccoli using foliar urea or methyl-jasmonate: Analysis of growth, gas exchange, and isotope composition. Plant Growth Regul. 2011, 63, 55–62. [Google Scholar] [CrossRef]
- Sheyhakinia, S.; Bamary, Z.; Einali, A.; Valizadeh, J. The induction of salt stress tolerance by jasmonic acid treatment in roselle (Hibiscus sabdariffa L.) seedlings through enhancing antioxidant enzymes activity and metabolic changes. Biologia 2020, 75, 681–692. [Google Scholar] [CrossRef]
- Abdelgawad, Z.A.; Khalafaallah, A.A.; Abdallah, M.M. Impact of Methyl Jasmonate on Antioxidant Activity and Some Biochemical Aspects of Maize Plant Grown under Water Stress Condition. Agric. Sci. 2014, 5, 1077–1088. [Google Scholar] [CrossRef] [Green Version]
- Sirhindi, G.; Mushtaq, R.; Singh Gill, S.; Sharma, P.; Abd-Allah, E.F.; Ahmad, P. Jasmonic acid and methyl jasmonate modulate growth, photosynthetic activity and expression of photosystem II subunit genes in Brassica oleracea L. Sci. Rep. 2020, 10, 9322. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatments | Total Length (cm) | Surf Area (cm2) | Root Dia. (mm) | Root Volume (cm3) |
|---|---|---|---|---|
| Control1 | 222.48 ± 38.19b | 35.46 ±5.66b | 0.54 ± 0.04a | 0.49 ± 0.08b |
| Control2 | 207.84 ± 29.75b | 38.99 ± 5.98b | 0.60 ± 0.04a | 0.62 ± 0.12ab |
| NaCl | 347.57 ± 49.33a | 65.05 ± 8.59a | 0.63 ± 0.05a | 1.01 ± 0.13a |
| MeJa | 207.90 ± 25.61b | 33.14 ± 4.64b | 0.50 ± 0.03a | 0.46 ± 0.086b |
| NaCl + MeJa | 219.14 ± 37.8b | 44.25 ± 6.69ab | 0.65 ± 0.05a | 0.75 ± 0.12ab |
| Treatment | Mg2+ | Ca2+ | K+ | NH4+ | Na+ | K/Na |
|---|---|---|---|---|---|---|
| Control1 | 309.00 ± 6.09ab | 997.72 ± 10.036a | 2411.01 ± 48.78ab | 277.59 ± 23.13a | 977.27 ± 70.43b | 2.47 ± 0.2a |
| Control2 | 299.25 ± 20.21ab | 901.05 ± 53.26a | 2295.72 ± 125.96ab | 251.19 ± 14.84a | 1086.10 ± 67.04b | 2.11 ± 0.2a |
| NaCl | 255.03 ± 53.10b | 692.83 ± 36.78b | 1628.10 ± 113.72c | 384.90 ± 15.00ab | 2698.07 ± 16.54a | 0.60 ± 0.1b |
| MeJa | 283.91 ± 12.15ab | 926.53 ± 28.21a | 2901.40 ± 121.81a | 449.18 ± 104.60b | 1308.48 ± 51.43b | 2.21 ± 0.2a |
| NaCl + MeJa | 342.78 ± 10.98a | 1044.16 ± 88.79a | 2006.77 ± 134.36b | 447.61 ± 48.50b | 2703.18 ± 44.96a | 0.74 ± 0.1b |
| Treatment | F− | Cl− | NO2− | Br− | NO3− | PO43− | C2O42− |
|---|---|---|---|---|---|---|---|
| Control1 | 20.88 ± 0.97a | 2285.63 ± 162.74b | 963.25 ± 8.94a | 109.62 ± 0.90b | 296.95 ± 23.32a | 944.38 ± 25.93c | 116.46 ± 4.42b |
| Control2 | 25.83 ± 4.56a | 2515.93 ± 104.79b | 845.95 ± 36.81a | 105.87 ± 0.86b | 229.12 ± 8.13a | 911.26 ± 25.64c | 108.28 ± 1.11b |
| NaCl | 21.71 ± 0.87a | 4385.71 ± 31.93a | 453.87 ± 90.22b | 109.38 ± 0.67b | 255.84 ± 9.10a | 940.02 ± 14.36c | 134.28 ± 1.89a |
| MeJa | 25.83 ± 1.25a | 2840.38 ± 4.00b | 970.44 ± 12.22a | 120.20 ± 1.40a | 276.49 ± 1.66a | 1082.62 ± 23.03b | 128.90 ± 7.94ab |
| NaCl + MeJa | 24.81 ± 0.75a | 4986.10 ± 39.67a | 954.57 ± 15.37a | 132.47 ± 0.81a | 292.59 ± 0.74a | 1278.91 ± 20.64a | 126.25 ± 6.58ab |
| Treatment | ||||||
|---|---|---|---|---|---|---|
| Fatty Acid (mg gr−1 DW) | Control1 | Control2 | MeJa | NaCl | MeJa + NaCl | |
| Saturated fatty acid (SFA) | ||||||
| Lauric acid | C12:0 | - | - | - | 0.09 ± 0.00b | - |
| Myristate acid | C14:0 | 0.07 ± 0.00a | 0.12 ± 0.01a | 0.11 ± 0.00a | 0.17 ± 0.03b | 0.11 ± 0.00a |
| Palmitate acid | C16:0 | 1.50 ± 0.02a | 1.65 ± 0.01a | 1.48 ± 0.02a | 2.12 ± 0.31b | 1.38 ± 0.00a |
| Stearate acid | C18:0 | 0.49 ± 0.04ab | 0.43 ± 0.02a | 0.47 ± 0.01ab | 0.58 ± 0.04b | 0.41 ± 0.03a |
| Arachidate acid | C20:0 | 0.12 ± 0.01a | 0.19 ± 0.02bc | 0.15 ± 0.00ab | 0.33 ± 0.03d | 0.21 ± 0.02c |
| Behenate acid | C22:0 | - | 0.29 ± 0.00b | 0.17 ± 0.00a | 0.30 ± 0.02b | 0.23 ± 0.06ab |
| Lignocerate acid | C24:0 | - | 0.29 ± 0.00b | 0.23 ± 0.00a | 0.26 ± 0.00b | - |
| Monounsaturated fatty acid (MUFA) | ||||||
| Oleic acid | C18:1 n9 | 0.41 ± 0.01a | 0.39 ± 0.00a | 0.31 ± 0.00a | 0.69 ± 0.13b | 0.42 ± 0.03a |
| Polyunsaturated fatty acid (PUFA) | ||||||
| Omega 6 (w6) | ||||||
| Linoleic acid. | C18:2 n6 | 8.76 ± 0.12a | 10.54 ± 0.40ab | 8.90 ± 0.17a | 12.44 ± 1.37b | 8.70 ± 0.20a |
| Omega 3 (w3) | ||||||
| Linolenic acid | C18:3 n3 | 7.14 ± 0.32a | 7.93 ± 0.04ab | 6.65 ± 0.35a | 10.37 ± 0.223b | 6.13 ± 0.00a |
| DBI (unsaturated fatty acid x number of double bonds) | 39.35 ± 2.12b | 44.2 ± 3.04ab | 56.68 ± 3.56a | 38.06 ± 2.16b | 36.21 ± 1.98b | |
| w6/w3 | 1.23 ± 0.07 n.s. | 1.33 ± 0.00 n.s. | 1.34 ± 0.04 n.s. | 1.26 ± 0.15 n.s. | 1.42 ± 0.04 n.s. | |
| Total | 18.53 | 21.83 | 18.48 | 27.29 | 17.62 | |
| Treatment | ||||||
|---|---|---|---|---|---|---|
| Fatty Acid (%) | Control1 | Control2 | MeJa | NaCl | MeJa + NaCl | |
| Saturated fatty acid (SFA) | ||||||
| Lauric acid | C12:0 | - | - | - | 0.003 | - |
| Myristate acid | C14:0 | 0.38 | 0.55 | 0.59 | 0.62 | 0.62 |
| Palmitate acid | C16:0 | 8.11 | 7.55 | 8.01 | 7.77 | 7.84 |
| Stearate acid | C18:0 | 2.65 | 1.96 | 2.54 | 2.12 | 2.33 |
| Arachidate acid | C20:0 | 0.65 | 0.87 | 0.81 | 1.21 | 1.19 |
| Behenate acid | C22:0 | - | 1.32 | 0.92 | 1.10 | 1.30 |
| Lignocerate acid | C24:0 | - | 1.32 | 1.24 | 0.95 | - |
| Monounsaturated fatty acid (MUFA) | ||||||
| Oleic acid | C18:1 n9 | 2.21 | 1.78 | 1.68 | 2.53 | 2.38 |
| Polyunsaturated fatty acid (PUFA) | ||||||
| Omega 6 (w6) | ||||||
| Linoleic acid | C18:2 n6 | 47.37 | 48.28 | 48.18 | 45.04 | 49.46 |
| Omega 3 (w3) | ||||||
| Linolenic acid | C18:3 n3 | 38.61 | 36.32 | 36.00 | 38.04 | 34.85 |
| Treatments | Total Phenolic compounds (mg GA kg−1 FW) | Total Flavonoids (mg Rutin kg−1 FW) | Antioxidant Capacity (mg DPPHreduced kg−1 FW) |
|---|---|---|---|
| Control1 | 891.79 ± 15.79a | 1965.41 ± 47.17b | 113.64 ± 7.17a |
| Control2 | 883.09 ± 8.11a | 1968.39 ± 6.30b | 110.85 ± 5.47a |
| NaCl | 833.53 ± 9.42b | 2167.24 ± 22.09a | 109.92 ± 5.84a |
| MeJa | 901.50 ± 9.71a | 2186.94 ± 597a | 108.81 ± 1.77a |
| MeJa + NaCl | 844.40 ± 7.08b | 2273.53 ± 0.60a | 117.78 ± 1.09a |
| Treatments | Ch a (mg kg−1 DW) | Ch b (mg kg−1 DW) | C (mg kg−1 DW) | Ch a/Ch b |
|---|---|---|---|---|
| Control1 | 12.68 ± 0.05b | 7.51 ± 0.40a | 2.43 ± 0.09b | 1.69 ± 0.00a |
| Control2 | 15.41 ± 0.23b | 7.92 ± 0.03a | 2.62 ± 0.03b | 1.94 ± 0.00c |
| NaCl | 16.13 ± 0.45b | 8.85 ± 0.29a | 2.86 ± 0.21b | 1.82 ± 0.00b |
| MeJa | 17.77 ± 0.05a | 4.98 ± 0.03b | 4.25 ± 0.00a | 3.56 ± 0.00e |
| NaCl + MeJa | 14.22 ± 0.01b | 4.24 ± 0.08b | 3.40 ± 0.01ab | 3.35 ± 0.00d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labiad, M.H.; Giménez, A.; Varol, H.; Tüzel, Y.; Egea-Gilabert, C.; Fernández, J.A.; Martínez-Ballesta, M.d.C. Effect of Exogenously Applied Methyl Jasmonate on Yield and Quality of Salt-Stressed Hydroponically Grown Sea Fennel (Crithmum maritimum L.). Agronomy 2021, 11, 1083. https://doi.org/10.3390/agronomy11061083
Labiad MH, Giménez A, Varol H, Tüzel Y, Egea-Gilabert C, Fernández JA, Martínez-Ballesta MdC. Effect of Exogenously Applied Methyl Jasmonate on Yield and Quality of Salt-Stressed Hydroponically Grown Sea Fennel (Crithmum maritimum L.). Agronomy. 2021; 11(6):1083. https://doi.org/10.3390/agronomy11061083
Chicago/Turabian StyleLabiad, M. Hatim, Almudena Giménez, Hafise Varol, Yüksel Tüzel, Catalina Egea-Gilabert, Juan A. Fernández, and María del Carmen Martínez-Ballesta. 2021. "Effect of Exogenously Applied Methyl Jasmonate on Yield and Quality of Salt-Stressed Hydroponically Grown Sea Fennel (Crithmum maritimum L.)" Agronomy 11, no. 6: 1083. https://doi.org/10.3390/agronomy11061083
APA StyleLabiad, M. H., Giménez, A., Varol, H., Tüzel, Y., Egea-Gilabert, C., Fernández, J. A., & Martínez-Ballesta, M. d. C. (2021). Effect of Exogenously Applied Methyl Jasmonate on Yield and Quality of Salt-Stressed Hydroponically Grown Sea Fennel (Crithmum maritimum L.). Agronomy, 11(6), 1083. https://doi.org/10.3390/agronomy11061083