
FTIR Screening to Elucidate Compositional Differences in Maize Recombinant Inbred Lines with Contrasting Saccharification Efficiency Yields

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Fourier-Transform Infrared Spectroscopy
2.2. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vermerris, W.; Saballos, A.; Ejeta, G.; Mosier, N.S.; Ladisch, M.R.; Carpita, N.C. Molecular breeding to enhance ethanol production from corn and sorghum stover. Crop Sci. 2007, 47, S142. [Google Scholar] [CrossRef]
- Van der Weijde, T.; Alvim Kamei, C.L.; Torres, A.F.; Vermerris, W.; Dolstra, O.; Visser, R.G.F.; Trindade, L.M. The potential of C4 grasses for cellulosic biofuel production. Front. Plant Sci. 2013, 4, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Vermerris, W. Cell wall Biosynthetic Genes of Maize and their Potential for Bioenergy Production. In Handbook of Maize; Springer: New York, NY, USA, 2009. [Google Scholar]
- Courtial, A.; Soler, M.; Chateigner-Boutin, A.-L.; Reymond, M.; Mechin, V.; Wang, H.; Grima-Pettenati, J.; Barriere, Y. Breeding grasses for capacity to biofuel production or silage feeding value: An updated list of genes involved in maize secondary cell wall biosynthesis and assembly. Maydica 2013, 58, 67–102. [Google Scholar]
- Dhugga, K.S. Maize biomass yield and composition for biofuels. Crop Sci. 2007, 47, 2211–2227. [Google Scholar] [CrossRef]
- Pauly, M.; Keegstra, K. Cell-wall carbohydrates and their modification as a resource for biofuels. Plant J. 2008, 54, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, R.; Cesarino, I.; Rataj, K.; Xiao, Y.; Sundin, L.; Goeminne, G.; Kim, H.; Cross, J.; Morreel, K.; Araujo, P.; et al. Genotypic variation in phenolic components of cell-walls in relation to the digestibility of maize stalks. Plant Physiol. 2010, 11, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Lapierre, C. Application of New Methods for the Investigation of Lignin Structure. In Forage Cell Wall Structure and Digestibility; American Society of Agronomy, Inc.: Madison, WI, USA, 1993; pp. 133–166. [Google Scholar] [CrossRef]
- Mosier, N.; Wyman, C.; Dale, B.; Elander, R.; Lee, Y.Y.; Holtzapple, M.; Ladisch, M. Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol. 2005, 96, 673–686. [Google Scholar] [CrossRef] [PubMed]
- McCann, M.C.; Carpita, N.C. Biomass recalcitrance: A multi-scale, multi-factor, and conversion-specific property. J. Exp. Bot. 2015, 66, 4109–4118. [Google Scholar] [CrossRef] [Green Version]
- Barrière, Y.; Méchin, V.; Riboulet, C.; Guillaumie, S.; Thomas, J.; Bosio, M.; Fabre, F.; Goffner, D.; Pichon, M.; Lapierre, C.; et al. Genetic and genomic approaches for improving biofuel production from maize. Euphytica 2009, 170, 183–202. [Google Scholar] [CrossRef]
- Selig, M.J.; Viamajala, S.; Decker, S.R.; Tucker, M.P.; Himmel, M.E.; Vinzant, T.B. Deposition of lignin droplets produced during dilute acid pretreatment of maize stems retards enzymatic hydrolysis of cellulose. Biotechnol. Prog. 2007, 23, 1333–1339. [Google Scholar] [CrossRef]
- Mansfield, S.D.; Mooney, C.; Saddler, J.N. Substrate and enzyme characteristics that limit cellulose hydrolysis. Biotechnol. Prog. 1999, 15, 804–816. [Google Scholar] [CrossRef]
- Berlin, A.; Balakshin, M.; Gilkes, N.; Kadla, J.; Maximenko, V.; Kubo, S.; Saddler, J. Inhibition of cellulase, xylanase and β-glucosidase activities by softwood lignin preparations. J. Biotechnol. 2006, 125, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Nakagame, S.; Chandra, R.P.; Saddler, J.N. The effect of isolated lignins, obtained from a range of pretreated lignocellulosic substrates, on enzymatic hydrolysis. Biotechnol. Bioeng. 2010, 105, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.F.; Visser, R.G.F.; Trindade, L.M. Bioethanol from maize cell walls: Genes, molecular tools, and breeding prospects. GCB Bioenergy 2015, 7, 591–607. [Google Scholar] [CrossRef]
- Ragauskas, A.J.; Beckham, G.T.; Biddy, M.J.; Chandra, R.; Chen, F.; Davis, M.F.; Davison, B.H.; Dixon, R.A.; Gilna, P.; Keller, M.; et al. Lignin valorization: Improving lignin processing in the biorefinery. Science 2014, 344, 6185. [Google Scholar] [CrossRef]
- López-Malvar, A.; Butron, A.; Malvar, R.A.; McQueen-Mason, S.J.; Faas, L.; Gómez, L.D.; Revilla, P.; Figueroa-Garrido, D.J.; Santiago, R. Association mapping for maize stover yield and saccharification efficiency using a multiparent advanced generation intercross (MAGIC) population. Sci. Rep. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Oliveira, D.M.; Mota, T.R.; Grandis, A.; Morais, G.R.D.; Lucas, R.C.D.; Polizeli, M.L.T.M.; Marchiosi, R.; Buckeridge, M.S.; Ferrarese-Filho, O.; Santos, W.D.D. Lignin plays a key role in determining biomass recalcitrance in forage grasses. Renew. Energy 2020, 147, 2206–2217. [Google Scholar] [CrossRef]
- Costa, R.M.F.D.; Barrett, W.; Carli, J.; Allison, G.G. Analysis of Plant Cell Walls by Attenuated Total Reflectance Fourier Transform Infrared Spectroscopy. In The Plant Cell Wall; Humana: New York, NY, USA, 2020; pp. 97–313. ISBN 9781071606193. [Google Scholar]
- Jiménez-Galindo, J.C.; Malvar, R.A.; Butrón, A.; Santiago, R.; Samayoa, L.F.; Caicedo, M.; Ordás, B. Mapping of resistance to corn borers in a MAGIC population of maize. BMC Plant Biol. 2019, 19, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Butrón, A.; Santiago, R.; Cao, A.; Samayoa, L.; Malvar, R. QTLs for Resistance to Fusarium Ear Rot in a Multiparent Advanced Generation Intercross (MAGIC) Maize Population. Plant Dis. 2019, 103, 897–904. [Google Scholar] [CrossRef]
- Gomez, L.D.; Whitehead, C.; Barakate, A.; Halpin, C.; McQueen-Mason, S.J. Automated saccharification assay for determination of digestibility in plant materials. Biotechnol. Biofuels 2010, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Faix, O. Classification of lignins from different botanical origins by FT-IR spectroscopy. Holzforsch. Int. J. Biol. Chem. Phys. Technol. Wood 1991, 43, 195–203. [Google Scholar] [CrossRef]
- Kacuráková, M.; Capek, P.; Sasinková, V.; Wellner, N.; Ebringerová, A. FT-IR study of plant cell wall model compounds: Pectic polysaccharides and hemicelluloses. Carbohydr. Polym. 2000, 43, 195–203. [Google Scholar] [CrossRef]
- Sills, D.L.; Gossett, J.M. Using FTIR to predict saccharification from enzymatic hydrolysis of alkali-pretreated biomasses. Biotechnol. Bioeng. 2012, 109, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xiuwen, W.; Hu, J.; Liu, Q.; Shen, D.; Xiao, R. Thermal degradation of softwood lignin and hardwood lignin by TG-FTIR and Py-GC/MS. Polym. Degrad. Stab. 2014, 108, 133–138. [Google Scholar] [CrossRef]
- Traoré, M.; Kaal, J.; Cortizas, A.M. Application of FTIR spectroscopy to the characterization of archeological wood. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2016, 153, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Cuello, C.; Marchand, P.; Laurans, F.; Grand-Perret, C.; Lainé-Prade, V.; Pilate, G.; Déjardin, A. ATR-FTIR microspectroscopy brings a novel insight into the study of cell wall chemistry at the cellular level. Front. Plant Sci. 2020, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Carpita, N.C.; Defernez, M.; Findlay, K.; Wells, B.; Shoue, D.A.; Catchpole, G.; Wilson, R.H.; McCann, M.C. Cell wall architecture of the elongating maize coleoptile. Plant Physiol. 2001, 127, 551–565. [Google Scholar] [CrossRef]
- Kubo, S.; Kadla, J.F. Hydrogen bonding in lignin: A fourier transform infrared model compound study. Biomacromolecules 2005, 6, 2815–2821. [Google Scholar] [CrossRef]
- McCann, M.C.; Defernez, M.; Urbanowicz, B.R.; Tewari, J.C.; Langewisch, T.; Olek, A.; Wells, B.; Wilson, R.H.; Carpita, N.C. Neural network analyses of infrared spectra for classifying cell wall architectures. Plant Physiol. 2007, 143, 1314–1326. [Google Scholar] [CrossRef] [Green Version]
- Szymanska-Chargot, M.; Zdunek, A. Use of FT-IR Spectra and PCA to the bulk characterization of cell wall residues of fruits and vegetables along a fraction process. Food Biophys. 2013, 8, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Abidi, N.; Cabrales, L.; Haigler, C.H. Changes in the cell wall and cellulose content of developing cotton fibers investigated by FTIR spectroscopy. Carbohydr. Polym. 2014, 100, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Bekiaris, G.; Lindedam, J.; Peltre, C.; Decker, S.R.; Turner, G.B.; Magid, J.; Bruun, S. Rapid estimation of sugar release from winter wheat straw during bioethanol production using FTIR-photoacoustic spectroscopy. Biotechnol. Biofuels 2015, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupoi, J.S.; Singh, S.; Parthasarathi, R.; Simmons, B.A.; Henry, R.J. Recent innovations in analytical methods for the qualitative and quantitative assessment of lignin. Renew. Sustain. Energy Rev. 2015, 49, 871–906. [Google Scholar] [CrossRef] [Green Version]
- Christophe, F.; Séné, B.; Mccann, M.C.; Wilson, R.H.; Crinter, R. Fourier-transform Raman and Fourier-transform Lnfrared spectroscopy. Plant Physiol. 1994, 106, 1623–1631. [Google Scholar]
- Pandey, K.K. A study of chemical structure of soft and hardwood and wood polymers by FTIR spectroscopy. J. Appl. Polym. Sci. 1999, 71, 1969–1975. [Google Scholar] [CrossRef]
- Åkerholm, M.; Salmén, L. Interactions between wood polymers studied by dynamic FT-IR spectroscopy. Polymer 2001, 42, 963–969. [Google Scholar] [CrossRef]
- McCann, M.C.; Bush, M.; Milioni, D.; Sado, P.; Stacey, N.J.; Catchpole, G.; Defernez, M.; Carpita, N.C.; Hofte, H.; Ulvskov, P.; et al. Approaches to understanding the functional architecture of the plant cell wall. Phytochemistry 2001, 57, 811–821. [Google Scholar] [CrossRef]
- Schulz, H.; Baranska, M. Identification and quantification of valuable plant substances by IR and Raman spectroscopy. Vib. Spectrosc. 2007, 43, 13–25. [Google Scholar] [CrossRef]
- Zhang, M.; Lapierre, C.; Nouxman, N.L.; Nieuwoudt, M.K.; Smith, B.G.; Chavan, R.R.; McArdle, B.H.; Harris, P.J. Location and characterization of lignin in tracheid cell walls of radiata pine (Pinus radiata D. Don) compression woods. Plant Physiol. Biochem. 2017, 118, 187–198. [Google Scholar] [CrossRef]
- Marchessault, R.H. To cellulose and wood polysaccharides. Pure Appl. Chem. 1962, 5, 107–130. [Google Scholar] [CrossRef]
- Harrington, K.J.; Higgins, H.G.; Michell, A.J. Infrared spectra of Eucalyptus regnans F. Muell. and Pinus radiata D. Don. Holzforschung Int. J. Biol. Chem. Phys. Technol. Wood 1964, 18, 108–113. [Google Scholar]
- Blackwell, J. Infrared and Raman Spectroscopy of Cellulose. In Cellulose Chemistry and Techology; ACS: Washington, DC, USA, 1977; pp. 206–218. [Google Scholar]
- Schwanninger, M.; Rodrigues, J.C.; Pereira, H.; Hinterstoisser, B. Effects of short-time vibratory ball milling on the shape of FT-IR spectra of wood and cellulose. Vib. Spectrosc. 2004, 36, 23–40. [Google Scholar] [CrossRef]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, S.Y.; Liu, Y.S.; Zeng, Y.; Himmel, M.E.; Baker, J.O.; Bayer, E.A. How does plant cell wall nanoscale architecture correlate with enzymatic digestibility? Science 2012, 338, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.; Li, X.; Bonawitz, N.D.; Chapple, C. Emerging strategies of lignin engineering and degradation for cellulosic biofuel production. Curr. Opin. Biotechnol. 2008, 19, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Weng, J.K.; Chapple, C. Improvement of biomass through lignin modification. Plant J. 2008, 54, 569–581. [Google Scholar] [CrossRef]
- Xiong, W.; Wu, Z.; Liu, Y.; Li, Y.; Su, K.; Bai, Z.; Guo, S.; Hu, Z.; Zhang, Z.; Bao, Y.; et al. Mutation of 4-coumarate: Coenzyme A ligase 1 gene affects lignin biosynthesis and increases the cell wall digestibility in maize brown midrib5 mutants. Biotechnol. Biofuels 2019, 12, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.; Su, R.; Qi, W.; He, Z. Bioconversion of lignocellulose into bioethanol: Process intensification and mechanism research. Bioenergy Res. 2011, 4, 225–245. [Google Scholar] [CrossRef]
- Fu, C.; Xiao, X.; Xi, Y.; Ge, Y.; Chen, F.; Bouton, J.; Dixon, R.A.; Wang, Z.Y. Downregulation of cinnamyl alcohol dehydrogenase (CAD) leads to improved saccharification efficiency in switchgrass. Bioenergy Res. 2011, 4, 153–164. [Google Scholar] [CrossRef]
- Fornalé, S.; Capellades, M.; Encina, A.; Wang, K.; Irar, S.; Lapierre, C.; Ruel, K.; Joseleau, J.P.; Berenguer, J.; Puigdomènech, P.; et al. Altered lignin biosynthesis improves cellulosic bioethanol production in transgenic maize plants down-regulated for cinnamyl alcohol dehydrogenase. Mol. Plant 2012, 5, 817–830. [Google Scholar] [CrossRef] [Green Version]
- Ralph, J.; Brunow, G.; Boerjan, W. Lignins. Encycl. Life Sci. 2007, 1–10. [Google Scholar] [CrossRef]
- Wilkerson, C.G.; Mansfield, S.D.; Lu, F.; Withers, S.; Park, J.Y.; Karlen, S.D.; Gonzales-Vigil, E.; Padmakshan, D.; Unda, F.; Rencoret, J.; et al. Monolignol ferulate transferase introduces chemically labile linkages into the lignin backbone. Science 2014, 344, 90–93. [Google Scholar] [CrossRef] [Green Version]
- Costa, R.M.F.D.; Pattathil, S.; Avci, U.; Winters, A.; Hahn, M.G.; Bosch, M. Desirable plant cell wall traits for higher-quality miscanthus lignocellulosic biomass. Biotechnol. Biofuels 2019, 12, 1–18. [Google Scholar] [CrossRef]
- Kumar, M.; Turner, S. Plant cellulose synthesis: CESA proteins crossing kingdoms. Phytochemistry 2015, 112, 91–99. [Google Scholar] [CrossRef]
- Hall, M.; Bansal, P.; Lee, J.H.; Realff, M.J.; Bommarius, A.S. Cellulose crystallinity—A key predictor of the enzymatic hydrolysis rate. FEBS J. 2010, 277, 1571–1582. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, Y.; Sugiyama, J.; Chanzy, H.; Langan, P. Crystal structure and hydrogen bonding system in cellulose Iα from synchrotron X-ray and neutron fiber diffraction. J. Am. Chem. Soc. 2003, 125, 14300–14306. [Google Scholar] [CrossRef]
- Grantham, N.J.; Wurman-Rodrich, J.; Terrett, O.M.; Lyczakowski, J.J.; Stott, K.; Iuga, D.; Simmons, T.J.; Durand-Tardif, M.; Brown, S.P.; Dupree, R.; et al. An even pattern of xylan substitution is critical for interaction with cellulose in plant cell walls. Nat. Plants 2017, 3, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Busse-Wicher, M.; Gomes, T.C.F.; Tryfona, T.; Nikolovski, N.; Stott, K.; Grantham, N.J.; Bolam, D.N.; Skaf, M.S.; Dupree, P. The pattern of xylan acetylation suggests xylan may interact with cellulose microfibrils as a twofold helical screw in the secondary plant cell wall of Arabidopsis thaliana. Plant J. 2014, 79, 492–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawar, P.M.A.; Derba-Maceluch, M.; Chong, S.L.; Gandla, M.L.; Bashar, S.S.; Sparrman, T.; Ahvenainen, P.; Hedenström, M.; Özparpucu, M.; Rüggeberg, M.; et al. In muro deacetylation of xylan affects lignin properties and improves saccharification of aspen wood. Biotechnol. Biofuels 2017, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| 2016 | 2017 | ||
|---|---|---|---|
| RILs | Saccharification (nmol mg−1 material−1 h−1) | RILs | Saccharification (nmol mg−1 material−1 h−1) |
| Low | |||
| EPS21LR-415 | 70.354 | EPS21LR-267 | 138.409 |
| EPS21LR-494 | 73.340 | EPS21LR-280 | 140.569 |
| EPS21LR-629 | 77.574 | EPS21LR-560 | 142.657 |
| EPS21LR-711 | 78.112 | EPS21LR-347 | 142.719 |
| EPS21LR-289 | 78.183 | EPS21LR-283 | 142.979 |
| EPS21LR-748 | 78.336 | EPS21LR-409 | 143.652 |
| EPS21LR-472 | 79.001 | EPS21LR-597 | 143.789 |
| EPS21LR-643 | 79.213 | EPS21LR-703 | 143.864 |
| EPS21LR-414 | 79.443 | EPS21LR-260 | 144.628 |
| EPS21LR-526 | 79.588 | EPS21LR-698 | 144.732 |
| EPS21LR-316 | 80.571 | EPS21LR-522 | 148.238 |
| EPS21LR-353 | 80.747 | EPS21LR-670 | 149.194 |
| EPS21LR-547 | 81.101 | EPS21LR-614 | 149.238 |
| EPS21LR-317 | 81.409 | EPS21LR-348 | 150.578 |
| EPS21LR-698 | 81.641 | EPS21LR-398 | 150.623 |
| EPS21LR-753 | 82.205 | EPS21LR-284 | 151.153 |
| EPS21LR-675 | 82.983 | EPS21LR-578 | 152.176 |
| EPS21LR-416 | 82.984 | EPS21LR-619 | 152.182 |
| EPS21LR-524 | 83.346 | EPS21LR-514 | 152.870 |
| EPS21LR-285 | 83.747 | EPS21LR-617 | 152.978 |
| EPS21LR-750 | 83.753 | EPS21LR-427 | 153.046 |
| EPS21LR-522 | 83.909 | EPS21LR-414 | 153.132 |
| EPS21LR-426 | 83.936 | EPS21LR-462 | 153.548 |
| EPS21LR-284 | 83.998 | EPS21LR-669 | 153.581 |
| EPS21LR-759 | 84.292 | EPS21LR-253 | 154.624 |
| EPS21LR-251 | 84.670 | EPS21LR-740 | 155.013 |
| EPS21LR-378 | 84.768 | EPS21LR-361 | 155.073 |
| EPS21LR-677 | 85.155 | EPS21LR-682 | 155.156 |
| EPS21LR-257 | 85.341 | EPS21LR-411 | 155.316 |
| EPS21LR-478 | 85.468 | EPS21LR-276 | 155.693 |
| Means | 81.306 | Means | 149.380 |
| Range | 70.354–85.468 | Range | 138.409–155.693 |
| High | |||
| EPS21LR-749 | 114.589 | EPS21LR-539 | 196.892 |
| EPS21LR-623 | 114.671 | EPS21LR-304 | 196.911 |
| EPS21LR-663 | 114.899 | EPS21LR-325 | 196.961 |
| EPS21LR-395 | 115.104 | EPS21LR-483 | 197.079 |
| EPS21LR-259 | 115.128 | EPS21LR-753 | 197.152 |
| EPS21LR-261 | 115.189 | EPS21LR-646 | 197.893 |
| EPS21LR-243 | 115.207 | EPS21LR-598 | 198.361 |
| EPS21LR-489 | 115.817 | EPS21LR-451 | 199.220 |
| EPS21LR-337 | 116.051 | EPS21LR-396 | 200.177 |
| EPS21LR-473 | 116.829 | EPS21LR-442 | 200.615 |
| EPS21LR-657 | 117.058 | EPS21LR-655 | 201.290 |
| EPS21LR-709 | 117.321 | EPS21LR-381 | 201.830 |
| EPS21LR-325 | 117.476 | EPS21LR-515 | 202.263 |
| EPS21LR-653 | 117.506 | EPS21LR-672 | 203.171 |
| EPS21LR-720 | 117.547 | EPS21LR-482 | 203.753 |
| EPS21LR-503 | 117.935 | EPS21LR-259 | 203.789 |
| EPS21LR-405 | 118.170 | EPS21LR-487 | 204.326 |
| EPS21LR-695 | 118.372 | EPS21LR-412 | 204.728 |
| EPS21LR-512 | 118.901 | EPS21LR-343 | 204.963 |
| EPS21LR-726 | 119.333 | EPS21LR-517 | 205.161 |
| EPS21LR-560 | 119.530 | EPS21LR-741 | 205.377 |
| EPS21LR-514 | 120.591 | EPS21LR-547 | 205.750 |
| EPS21LR-741 | 120.596 | EPS21LR-395 | 205.809 |
| EPS21LR-733 | 120.959 | EPS21LR-508 | 206.084 |
| EPS21LR-668 | 121.101 | EPS21LR-248 | 208.187 |
| EPS21LR-364 | 121.287 | EPS21LR-723 | 209.809 |
| EPS21LR-246 | 121.350 | EPS21LR-694 | 211.761 |
| EPS21LR-584 | 123.927 | EPS21LR-594 | 212.177 |
| EPS21LR-511 | 125.722 | EPS21LR-679 | 218.327 |
| EPS21LR-743 | 127.652 | EPS21LR-502 | 218.694 |
| Means | 118.527 | Means | 203.950 |
| Range | 114.589–127-652 | Range | 198.892–218.694 |
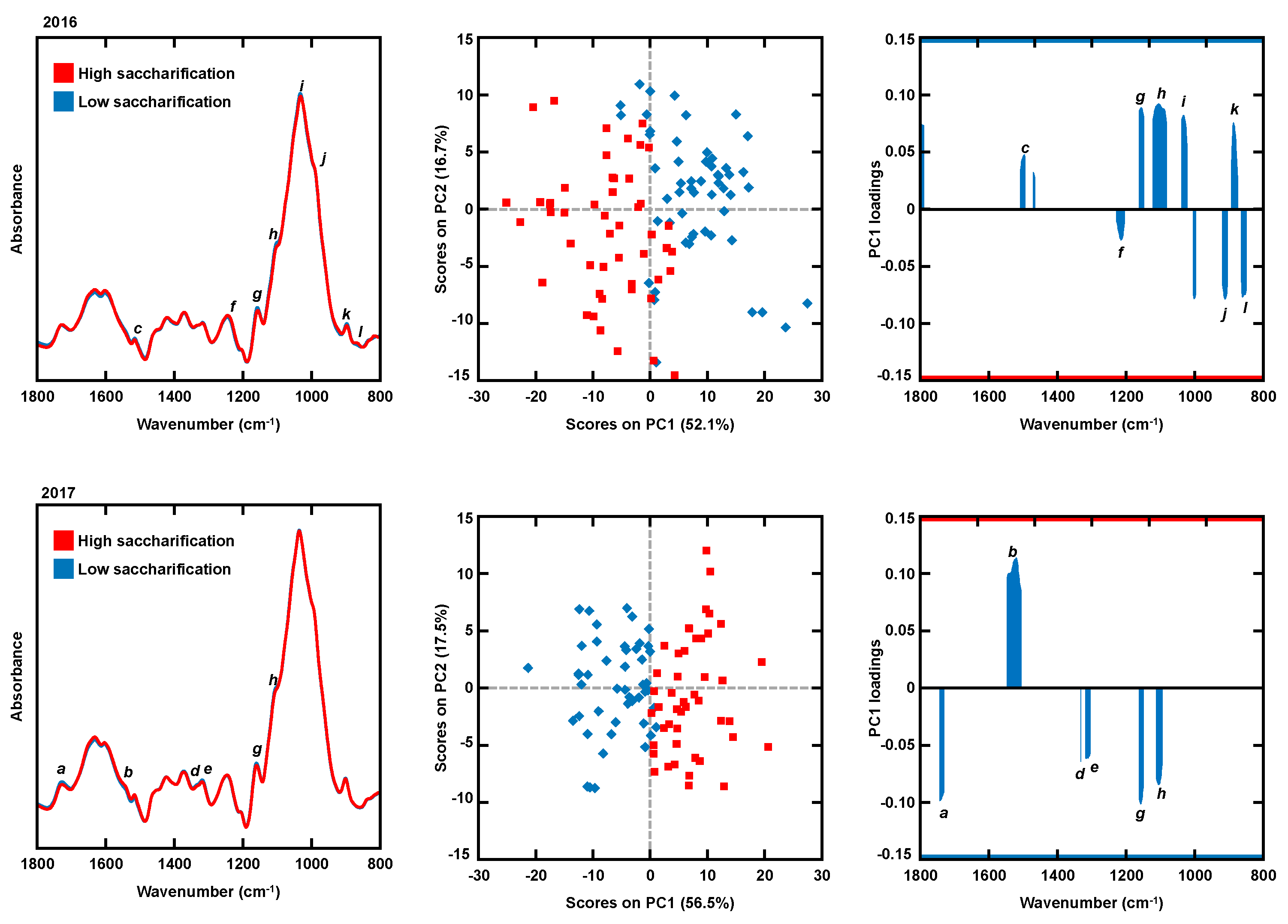
| PC1 Loading Spectral Region | Wavenumber (cm−1) | Reference | Assignment | Biomass Constituent |
|---|---|---|---|---|
| a | 1740 1735 1734 | [37,40,43,44] | C=O stretching | Acetyl and methyl esters |
| b | 1550 | [29,30,37] | Amide II (N-H deformation and stretching contribution from C-N stretching) | Proteins |
| c | 1504 1500 | [26,28] | Aromatic skeletal vibrations in guaiacyl rings | Lignin |
| d | 1336 | [38,41] | C-H in plane deformation | Cellulose |
| e | 1328 1311 1318 1311 | [38,39,43,44,45,46] | OH in-plane bending CH2 wagging CH in plane scissoring | Cellulose |
| f | 1224 1219 1220 | [27,28] | C–O stretch in syringyl rings | Lignin |
| g | 1160 1161 1163 1157 | [30,31,32,33,34,35] | C–O–C asymmetric stretching | Crystalline cellulose; associated to modifications cellulose-I > cellulose-II; linked to celluloses crystallinity features |
| h | 1116 | [31,36] | Aromatic C–H deformation | Lignin |
| i | 1035 1041 1040 | [24,29,47] | Aromatic C–H in plane deformation, G > S; plus C–O deformation in primary alcohols; plus C=O stretch (unconjugated) | Lignin |
| j | 915–925 | [24,29] | C-H out-of-plane; aromatic compounds | Aromatic compounds |
| k | 898 | [35] | C–O–C stretching | Cellulose |
| l | 872 875 | [25] | Glycosidic linkage in hemicelluloses | Hemicellulose |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Malvar, A.; Santiago, R.; Malvar, R.A.; Martín, D.; Pereira dos Santos, I.; Batista de Carvalho, L.A.E.; Faas, L.; Gómez, L.D.; da Costa, R.M.F. FTIR Screening to Elucidate Compositional Differences in Maize Recombinant Inbred Lines with Contrasting Saccharification Efficiency Yields. Agronomy 2021, 11, 1130. https://doi.org/10.3390/agronomy11061130
López-Malvar A, Santiago R, Malvar RA, Martín D, Pereira dos Santos I, Batista de Carvalho LAE, Faas L, Gómez LD, da Costa RMF. FTIR Screening to Elucidate Compositional Differences in Maize Recombinant Inbred Lines with Contrasting Saccharification Efficiency Yields. Agronomy. 2021; 11(6):1130. https://doi.org/10.3390/agronomy11061130
Chicago/Turabian StyleLópez-Malvar, Ana, Rogelio Santiago, Rosa Ana Malvar, Daniel Martín, Inês Pereira dos Santos, Luís A. E. Batista de Carvalho, Laura Faas, Leonardo D. Gómez, and Ricardo M. F. da Costa. 2021. "FTIR Screening to Elucidate Compositional Differences in Maize Recombinant Inbred Lines with Contrasting Saccharification Efficiency Yields" Agronomy 11, no. 6: 1130. https://doi.org/10.3390/agronomy11061130
APA StyleLópez-Malvar, A., Santiago, R., Malvar, R. A., Martín, D., Pereira dos Santos, I., Batista de Carvalho, L. A. E., Faas, L., Gómez, L. D., & da Costa, R. M. F. (2021). FTIR Screening to Elucidate Compositional Differences in Maize Recombinant Inbred Lines with Contrasting Saccharification Efficiency Yields. Agronomy, 11(6), 1130. https://doi.org/10.3390/agronomy11061130