
Warm and Wet Autumns Favour Yield Losses of Oilseed Rape Caused by Phoma Stem Canker




Abstract
:1. Introduction
2. Materials and Methods
2.1. Location of Experimental Sites
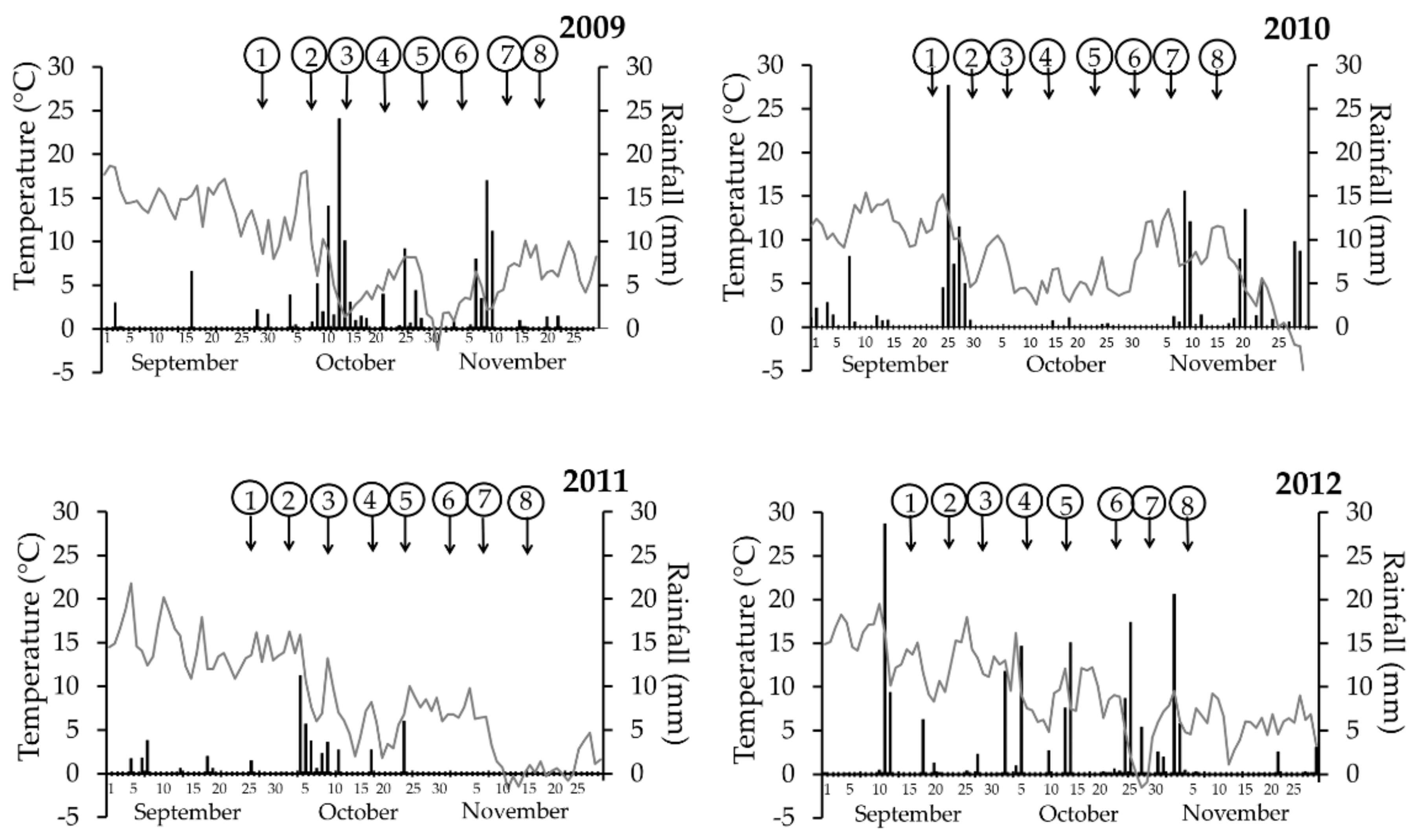
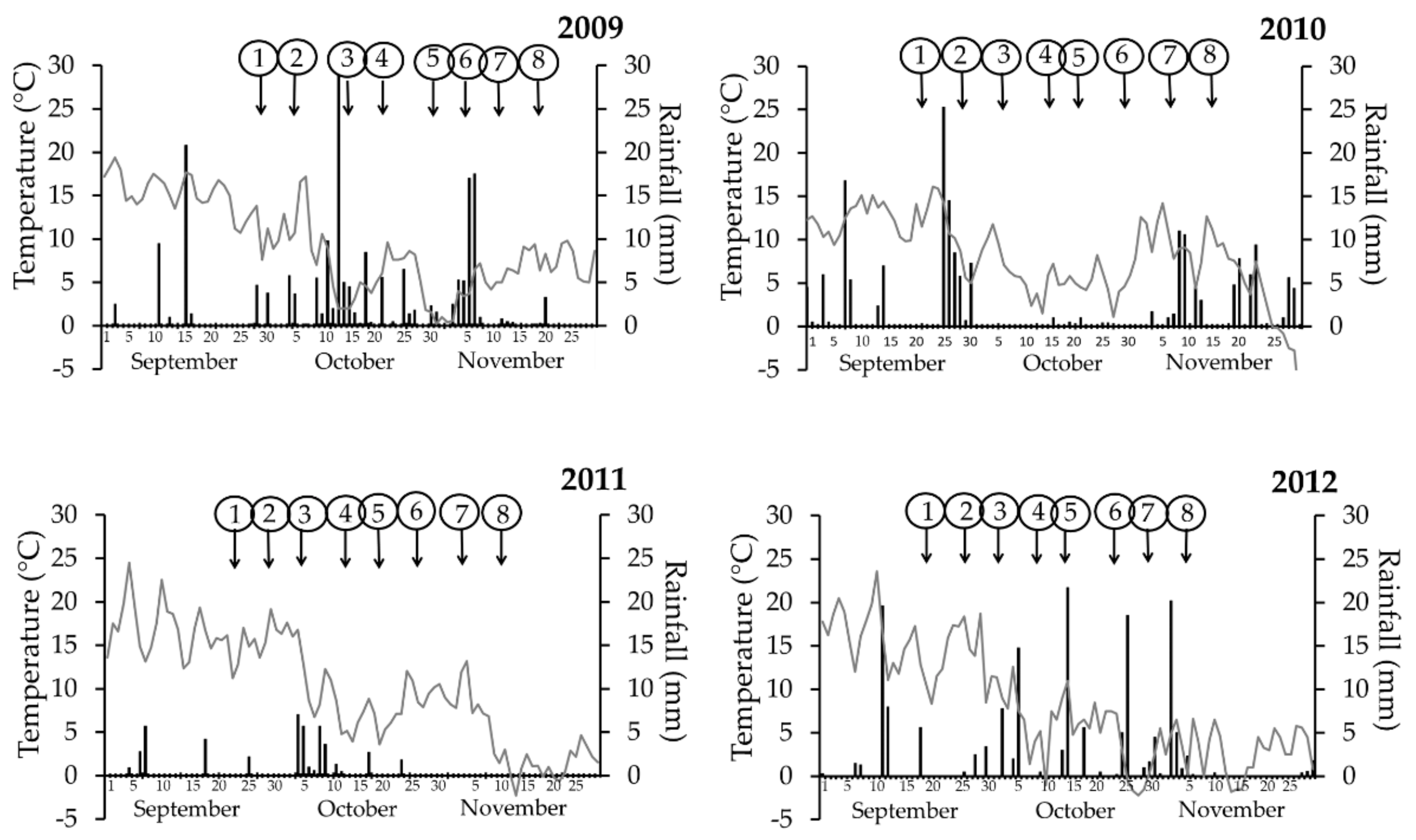
2.2. Meteorological Data
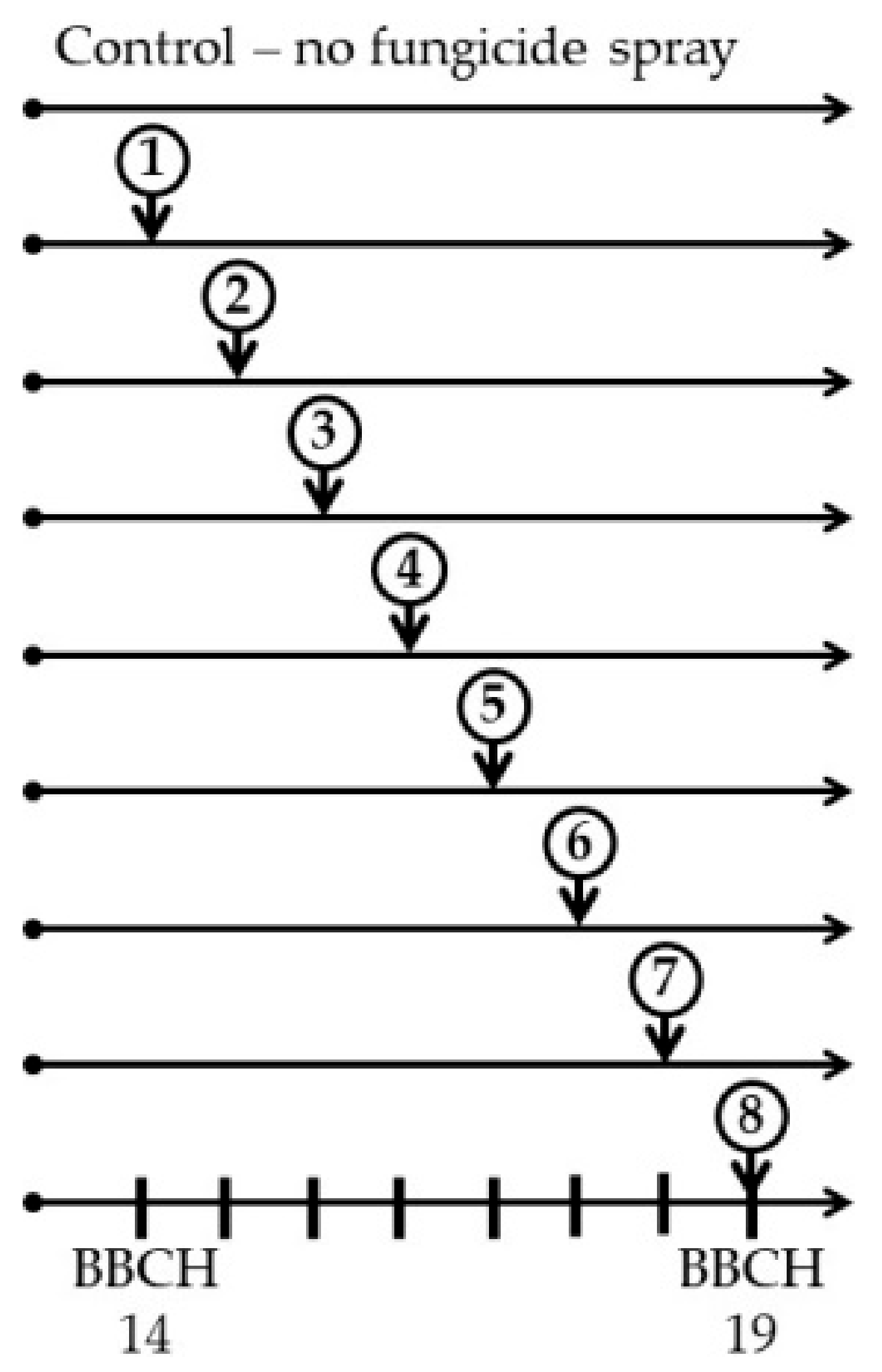
2.3. Field Experiments
2.4. Study of Seed Quality
2.5. Sampling, Isolation, and Identification of Leptosphaeria Isolates from Plants
2.6. Statistical Analyses
3. Results
3.1. Meteorological Data
3.2. Composition of the Pathogen Population
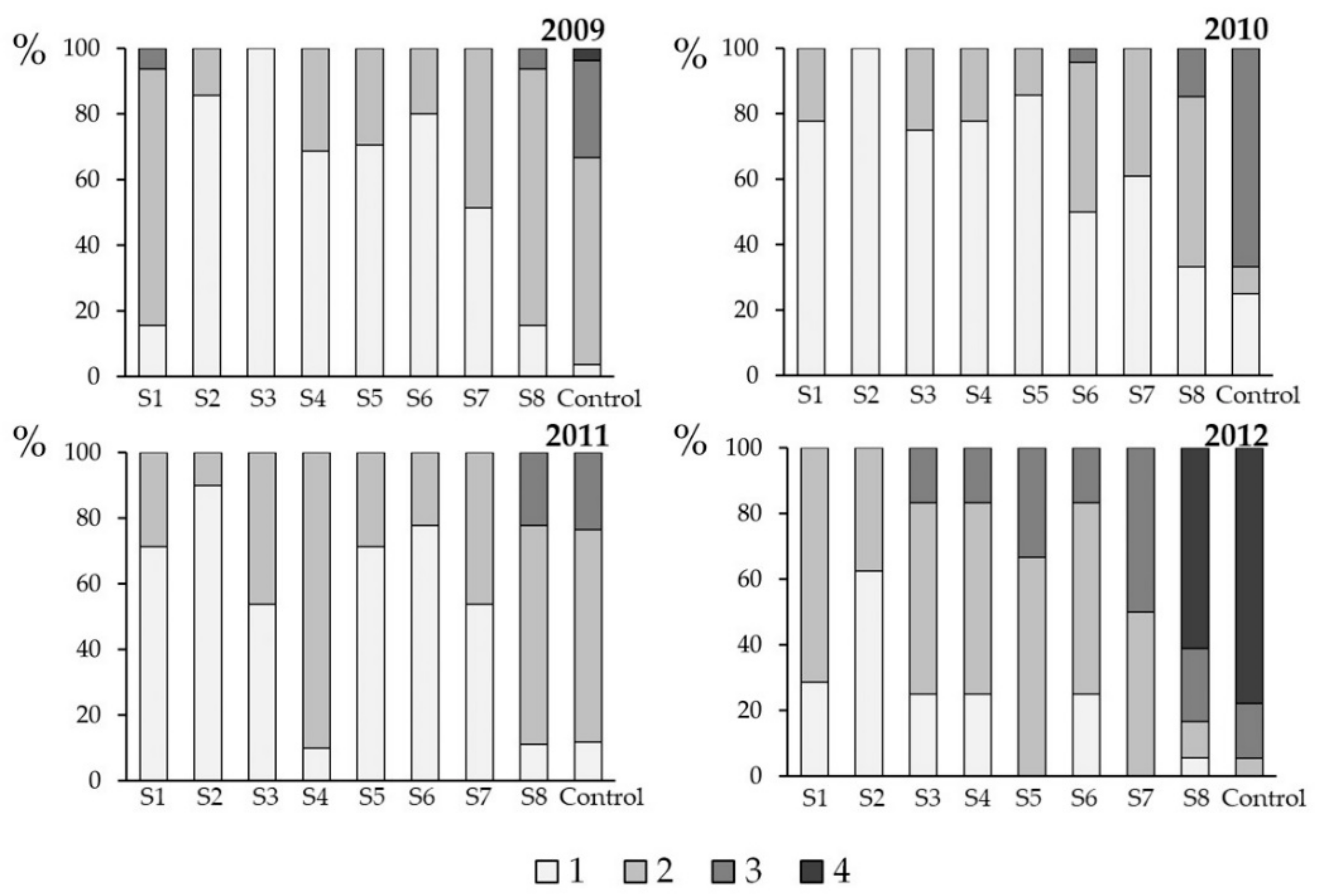
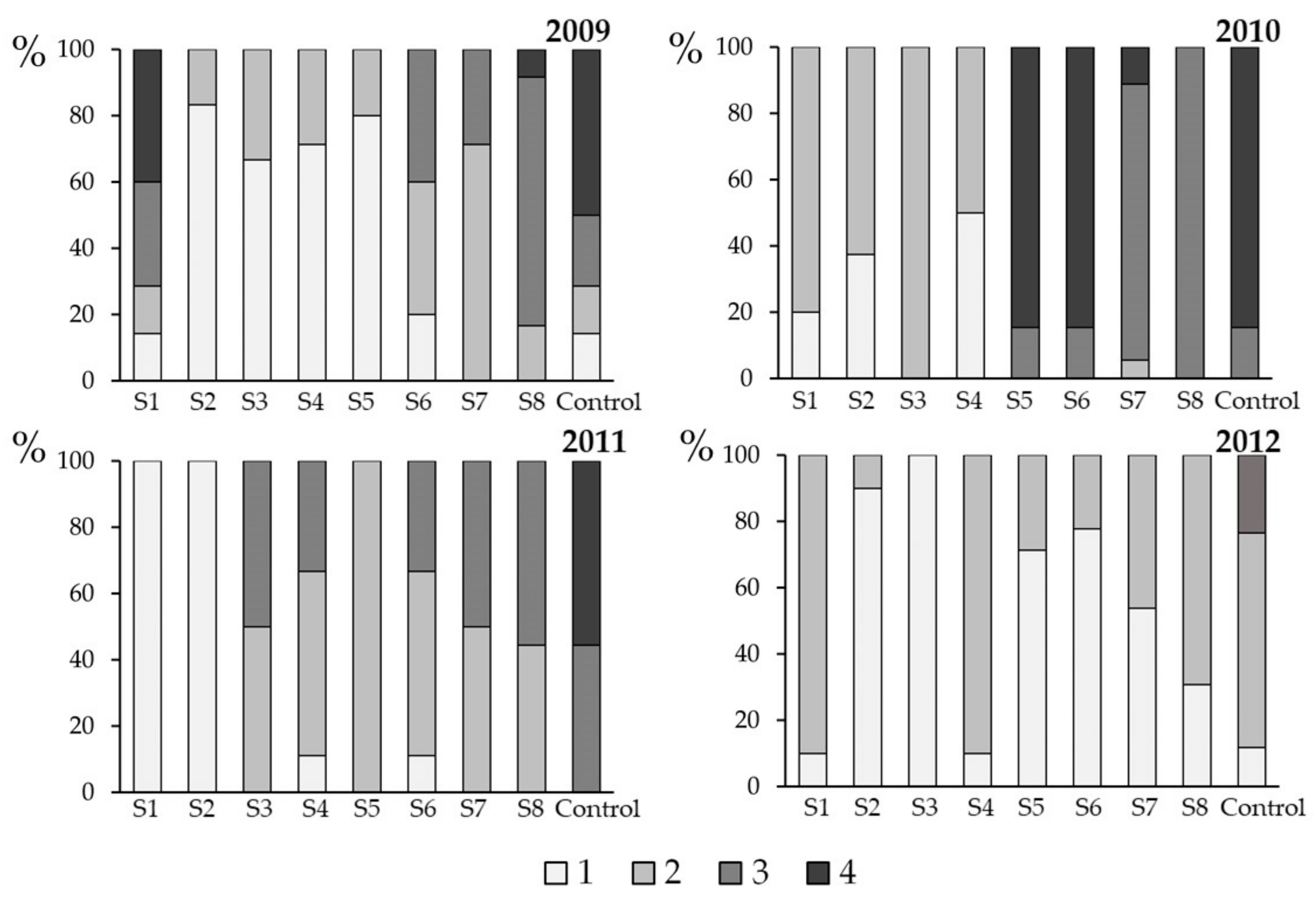
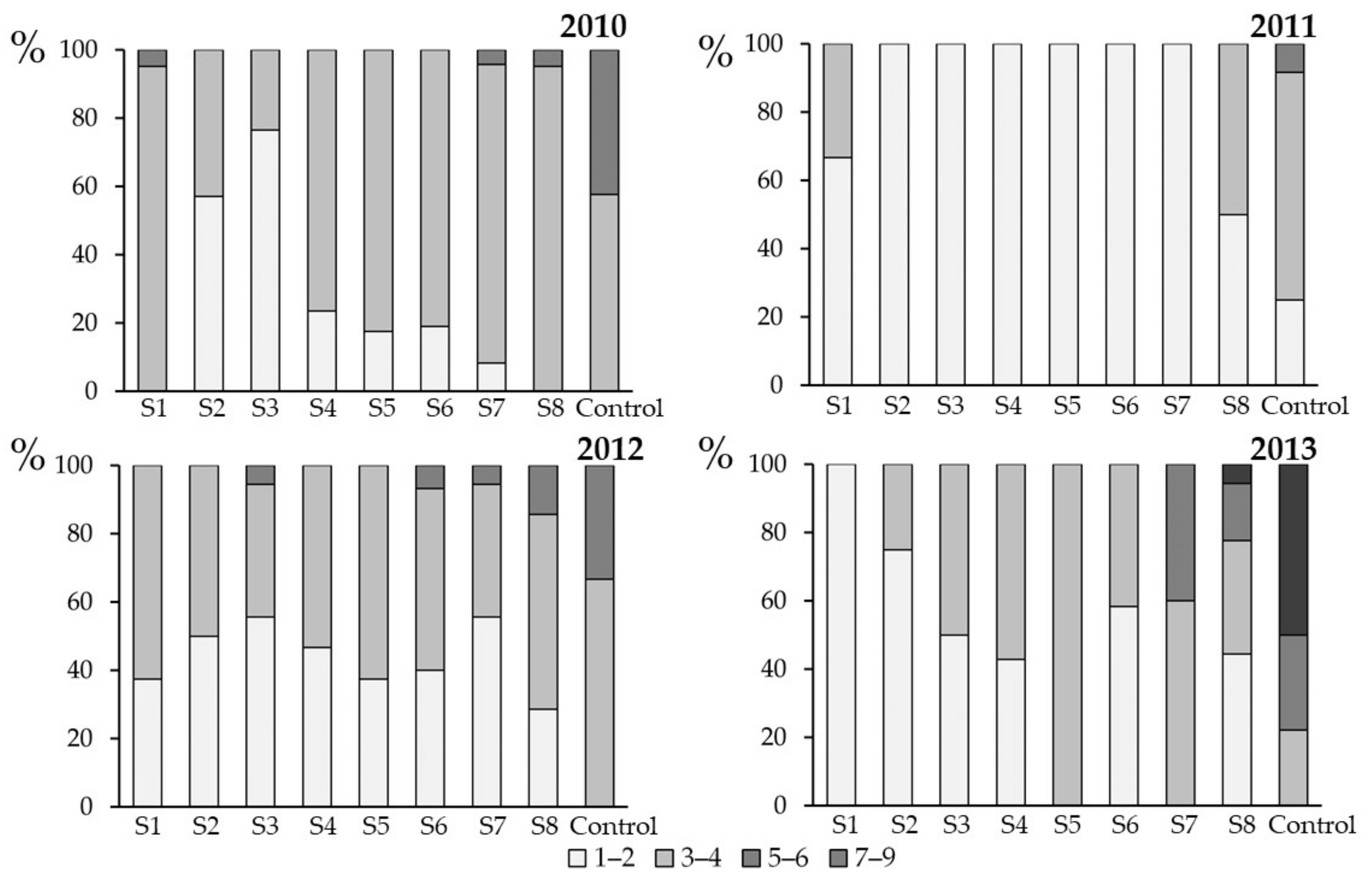
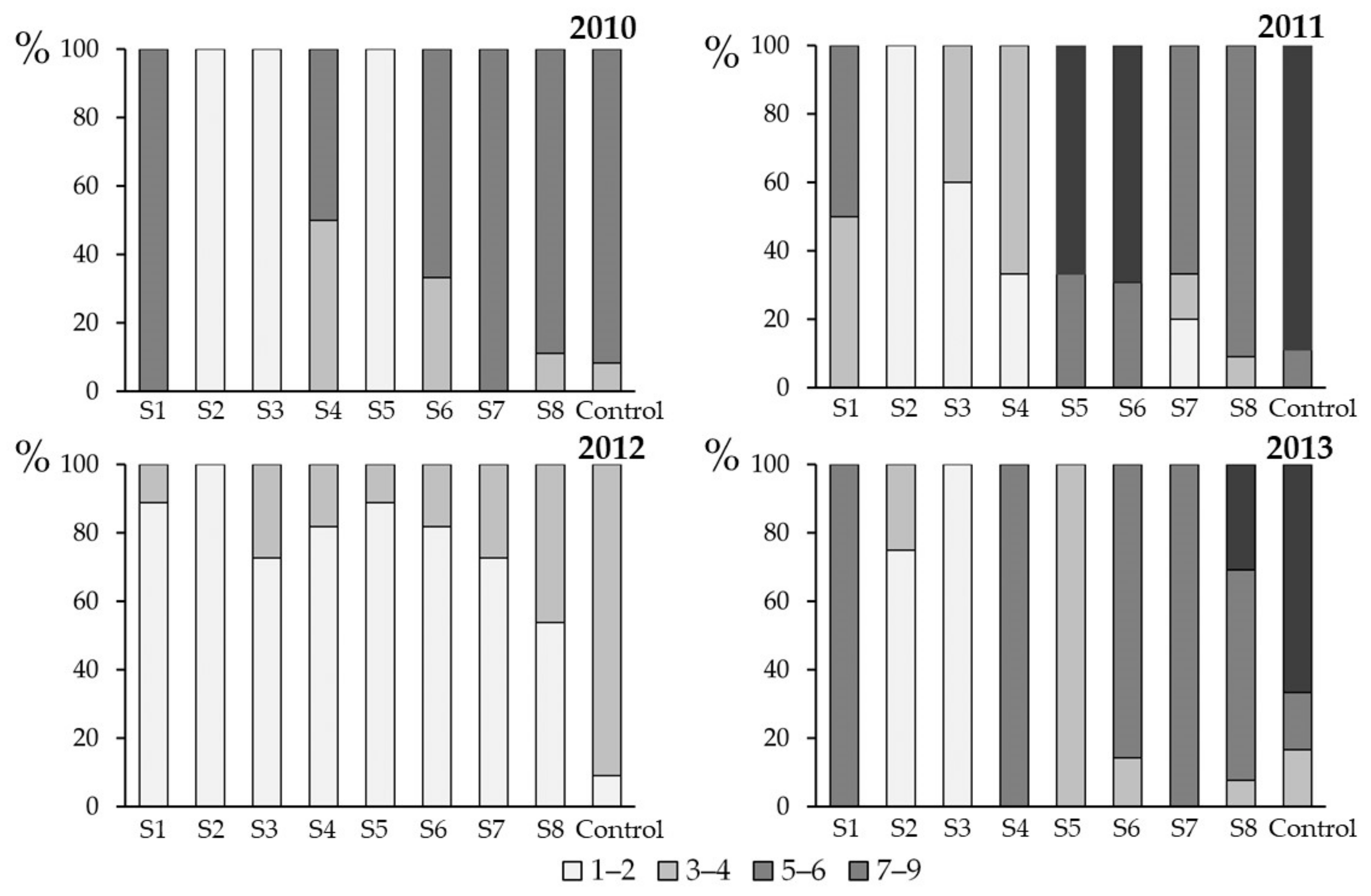
3.3. Effect of Spray Time on Plant Health
3.4. Effect of Spray Time on Seed Yield
3.5. Effect of Spray Time on Seed Quality
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carre, P.; Pouzet, A. Rapeseed market, worldwide and in Europe. OCL 2014, 21, D102. [Google Scholar] [CrossRef]
- EU-28. Oilseeds and Products Annual 2018. Global Agricultural Information Network Report, 2018. Available online: https://apps.fas.usda.gov/newgainapi/api/report/downloadreportbyfilename?filename=Oilseeds%20and%20Products%20Annual_Vienna_EU-28_3-29-2018.pdf (accessed on 30 March 2021).
- Grain Central. Rapeseed Oil Consumption Growing Globally. Available online: https://www.graincentral.com/cropping/oilseeds/rapeseed-oil-consumption-growing-globally/ (accessed on 30 March 2021).
- Lin, L.; Allemekinders, H.; Dansby, A.; Campbell, L.; Durance-Tod, S.; Berger, A.; Jones, P. Evidence of health benefits of canola oil. Nutr. Rev. 2013, 71, 370–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gül, M.K.; Amar, S. Sterols and the phytosterol content in oilseed rape (Brassica napus L.). J. Cell Mol. Biol. 2006, 5, 71–79. [Google Scholar]
- Jakobsen, G.V.; Jensen, B.B.; Knudsen, K.E.B.; Canibe, N. Improving the nutritional value of rapeseed cake and wheat dried distillers grains with solubles by addition of enzymes during liquid fermentation. Anim. Feed Sci. Technol. 2015, 208, 198–213. [Google Scholar] [CrossRef]
- Mansour, E.H.; Dworschák, E.; Lugasi, A.; Gaál, Ö.; Barna, É.; Gergely, A. Effect of processing on the antinutritive factors and nutritive value of rapeseed products. Food Chem. 1993, 47, 247–252. [Google Scholar] [CrossRef]
- Dawidziuk, A.; Kaczmarek, J.; Podlesna, A.; Kasprzyk, I.; Jedryczka, M. Influence of meteorological parameters on Leptosphaeria maculans and L. biglobosa spore release in central and eastern Poland. Grana 2012, 51, 240–248. [Google Scholar] [CrossRef]
- Fitt, B.D.L.; Brun, H.; Barbetti, M.J.; Rimmer, S.R. World-wide importance of phoma stem canker (Leptosphaeria maculans and L. biglobosa) on oilseed rape (Brassica napus). Eur. J. Plant Pathol. 2006, 114, 3–15. [Google Scholar] [CrossRef]
- Aubertot, J.N.; Pinochet, X.; Doré, T. The effects of sowing date and nitrogen availability during vegetative stages on Leptosphaeria maculans development on winter oilseed rape. J. Crop Prot. 2004, 23, 635–645. [Google Scholar] [CrossRef]
- Fitt, B.D.L.; Huang, Y.J.; van den Bosch, F.; West, J.S. Coexistence of related pathogen species on arable crops in space and time. Annu. Rev. Phytopathol. 2006, 44, 163–182. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.W.; Fernando, W.G.D.; Entz, M. Effects of crop rotation and tillage on blackleg disease of canola. Can. J. Plant Pathol. 2005, 27, 53–57. [Google Scholar] [CrossRef]
- Eckert, M.; Gout, L.; Rouxel, T.; Blaise, F.; Jedryczka, M.; Fitt, B.; Balesdent, M.H. Identification and characterization of polymorphic minisatellites in the phytopathogenic ascomycete Leptosphaeria maculans. Curr. Genet. 2005, 47, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Marcroft, S.J.; Sosnowski, M.R.; Scott, E.S.; Ramsey, M.D.; Salisbury, P.A.; Howlett, B.J. Brassica napus plants infected by Leptosphaeria maculans after the third to fifth leaf growth stage in south-eastern Australia do not develop blackleg stem canker. Eur. J. Plant Pathol. 2005, 112, 289–292. [Google Scholar] [CrossRef]
- Fernando, W.; Chen, Y.; Ghanbarnia, K. Breeding for blackleg resistance: The biology and epidemiology. Adv. Bot. Res. 2007, 45, 271–311. [Google Scholar]
- Salam, M.U.; Fitt, B.D.L.; Aubertot, J.N.; Diggle, A.J.; Huang, Y.J.; Barbetti, M.J.; Gladders, P.; Jedryczka, M.; Khangura, R.K.; Wratten, N.; et al. Two weather-based models for predicting the onset of seasonal release of ascospores of Leptosphaeria maculans or L. biglobosa. Plant Pathol. 2007, 56, 412–423. [Google Scholar] [CrossRef]
- Savage, D.; Barbetti, M.J.; MacLeod, W.; Salam, M.U.; Renton, M. Seasonal and diurnal patterns of spore release can significantly affect the proportion of spores expected to undergo long-distance dispersal. Microb. Ecol. 2012, 63, 578–585. [Google Scholar] [CrossRef]
- West, J.S.; Kharbanda, P.D.; Barbetti, M.J.; Fitt, B.D.L. Epidemiology and management of Leptosphaeria maculans (phoma stem canker) on oilseed rape in Australia, Canada and Europe. Plant Pathol. 2001, 50, 10–27. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-J.; Evans, N.; Li, Z.-Q.; Eckert, M.; Chevre, A.-M.; Renard, M.; Fitt, B.D.L. Temperature and leaf wetness duration affect phenotypic expression of Rlm6-mediated resistance to Leptosphaeria maculans in Brassica napus. New Phytol. 2006, 170, 129–141. [Google Scholar] [CrossRef]
- Sadys, M.; Kaczmarek, J.; Grinn-Gofron, A.; Rodinkova, V.; Prikhodko, A.; Bilous, E.; Strzelczak, A.; Herbert, R.J.; Jedryczka, M. Dew point temperature affects ascospore release of allergenic genus Leptosphaeria. Int. J. Biometeorol. 2018, 62, 979–990. [Google Scholar] [CrossRef] [Green Version]
- Evans, N.; Baierl, A.; Semenov, M.A.; Gladders, P.; Fitt, B.D.L. Range and severity of a plant disease increased by global warming. J. R. Soc. Interface 2008, 5, 525–533. [Google Scholar] [CrossRef] [Green Version]
- Pérès, A.; Poisson, B.; Le Sourne, V.; Maisonneuve, C. Leptosphaeria maculans: Effect of temperature, rainfall and humidity on the formation of pseudothecia. In Proceedings of the 10th International Rapeseed Congress, Canberra, Australia, 26–29 September 1999; Available online: http://www.regional.org.aa/au/gcirc/index.html (accessed on 30 March 2021).
- Petrie, G.A. Effects of temperature and moisture on the number, size and septation of ascospores produced by Leptosphaeria maculans (Blackleg) on rapeseed stubble. Can. Plant Dis. Surv. 1994, 74, 141–151. [Google Scholar]
- Toscano-Underwood, C.; Huang, Y.J.; Fitt, B.D.L.; Hall, A.M. Effects of temperature on maturation of pseudothecia of Leptosphaeria maculans and L. biglobosa on oilseed rape stem debris. Plant Pathol. 2003, 52, 726–736. [Google Scholar] [CrossRef]
- Huang, Y.J.; Liu, Z.; West, J.S.; Todd, A.D.; Hall, A.M.; Fitt, B.D.L. Effects of temperature and rainfall on date of release of ascospores of Leptosphaeria maculans (phoma stem canker) from winter oilseed rape (Brassica napus) debris in the UK. Ann. Appl. Biol. 2007, 151, 99–111. [Google Scholar] [CrossRef]
- Piliponyte-Dzikiene, A.; Kaczmarek, J.; Petraitiene, E.; Kasprzyk, I.; Brazauskiene, I.; Brazauskas, G.; Jedryczka, M. Microscopic and molecular detection of airborne ascospores of Leptosphaeria maculans and L. biglobosa in Lithuania and Poland. Zemdirbyste 2014, 101, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Penaud, A.; Jain, L.; Poisson, B.; Balesdent, M.H. Structure of populations of Leptosphaeria maculans in France. In Proceedings of the 10th International Rapeseed Congress, Canberra, Australia, 26–29 September 1999; Available online: http://www.regional.org.au/au/gcirc (accessed on 30 March 2021).
- Kuswinanti, T.; Koopmann, B.; Hoppe, H.H. Virulence pattern of aggressive isolates of Leptosphaeria maculans on an extended set of Brassica differentials. J. Plant Dis. Prot. 1999, 106, 12–20. [Google Scholar]
- Humpherson-Jones, F.M. Pathogenicity studies on isolates of Leptosphaeria maculans from brassica seed production crops in south-east England. Ann. Appl. Biol. 1983, 103, 37–44. [Google Scholar] [CrossRef]
- Gugel, R.K.; Petrie, G.A. History, occurrence, impact, and control of blackleg of rapeseed. Can. J. Plant Pathol. 1992, 14, 36–45. [Google Scholar] [CrossRef]
- Moreno-Rico, O.; Frias-Trevino, A.G.; Luna-Ruiz, J.J.; Manzano-Flores, D.E.; Romero-Cova, S.; Seguin-Swartz, G. Characterization and pathogenicity of isolates of Leptosphaeria maculans from Aguascalientes and Zacatecas, Mexico. Can. J. Plant Pathol. 2001, 23, 270–278. [Google Scholar] [CrossRef]
- Fernando, W.G.D.; Parks, P.S. First report of blackleg disease caused by Leptosphaeria maculans on canola in Brazil. Plant Dis. 2003, 87, 314. [Google Scholar] [CrossRef]
- Gaetan, S.A. First outbreak of blackleg caused by Phoma lingam in commercial canola fields in Argentina. Plant Dis. 2005, 89, 435. [Google Scholar] [CrossRef]
- Jedryczka, M.; Kaczmarek, J.; Dawidziuk, A.; Brachaczek, A. System for forecasting disease epidemics—Aerobiological methods in Polish agriculture. Asp. Appl. Biol. 2008, 89, 65–70. [Google Scholar]
- Lô-Pelzer, E.; Bousset, L.; Jeuffroy, M.H.; Salam, M.U.; Pinochet, X.; Boillot, M.; Aubertot, J.N. SIPPOM WOSR: A simulator for integrated pathogen population management of phoma stem canker on winter oilseed rape: I. Description of the model. Field Crop. Res. 2010, 118, 73–81. [Google Scholar] [CrossRef]
- West, J.S.; Evans, N.; Leech, P.K.; Fitt, B.D.L.; Welham, S.J.; Jedryczka, M.; Penaud, A. Predicting leaf infection by Leptosphaeria maculans on oilseed rape. Bull. IOBC 2000, 23, 23–27. [Google Scholar]
- Steed, J.M.; Baierl, A.; Fitt, B.D.L. Relating plant and pathogen development to optimise fungicide control of phoma stem canker (Leptosphaeria maculans) on winter oilseed rape (Brassica napus). Eur. J. Plant Pathol. 2007, 118, 359–373. [Google Scholar] [CrossRef]
- Stonard, J.F.; Latunde-Dada, A.O.; Huang, Y.J.; West, J.S.; Evans, N.; Fitt, B.D.L. Geographic variation in severity of phoma stem canker and Leptosphaeria maculans/L. biglobosa populations on UK winter oilseed rape (Brassica napus). Eur. J. Plant Pathol. 2010, 126, 97–109. [Google Scholar] [CrossRef]
- Eckert, M.R.; Rossall, S.; Selley, A.; Fitt, B.D.L. Effects of fungicides on in vitro spore germination and mycelial growth of the phytopathogens Leptosphaeria maculans and L. biglobosa (phoma stem canker of oilseed rape). Pest Manag. Sci. 2010, 66, 396–405. [Google Scholar] [CrossRef]
- Kaczmarek, J.; Jędryczka, M. Wpływ wybranych fungicydów oraz ich substancji aktywnych na wzrost grzybów Leptosphaeria maculans i L. biglobosa w warunkach in vitro. Prog. Plant Prot. 2010, 50, 648–651. [Google Scholar]
- Brachaczek, A.; Kaczmarek, J.; Jędryczka, M. Monitoring blackleg (Leptosphaeria spp.) ascospore release timing and quantity enables optimal fungicide application to improved oilseed rape yield and seed quality. Eur. J. Plant Pathol. 2016, 145, 643–657. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.K. Toxicity of Fungicides. In Veterinary Toxicology, 3rd ed.; Gupta, R.C., Ed.; Academic Press: San Diego, CA, USA, 2018; Volume 3, pp. 569–580. [Google Scholar]
- Roberts, J.R.; Reigart, J.R. Fungicides. In Recognition and Management of Pesticide Poisonings, 6th ed.; U.S. Environmental Protection Agency: Washington, DC, USA, 2013; pp. 143–160. [Google Scholar]
- Petit, A.N.; Fontaine, F.; Vatsa, P.; Clément, C.; Vaillant-Gaveau, N. Fungicide impacts on photosynthesis in crop plants. Photosynth. Res. 2012, 111, 315–326. [Google Scholar] [CrossRef]
- Baćmaga, M.; Wyszkowska, J.; Kucharski, J. The influence of chlorothalonil on the activity of soil microorganisms and enzymes. Ecotoxicology 2018, 27, 1188–1202. [Google Scholar] [CrossRef]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Brühl, C.A.; Imfeld, G.; Knäbel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmüller, A.; et al. Fungicides: An overlooked pesticide class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef]
- Fustinoni, S.; Rosa Mercadante, R.; Polledri, E.; Rubino, F.M.; Mandic-Rajcevic, S.; Vianello, G.; Colosio, C.; Moretto, A. Biological monitoring of exposure to tebuconazole in winegrowers. J. Expo. Sci. Environ. Epidemiol. 2014, 24, 643–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Green Deal. Available online: https://ec.europa.eu/info/strategy/priorities-2019-2024/european-green-deal_en (accessed on 30 March 2021).
- Brachaczek, A.; Kamiński, M.; Kaczmarek, J.; Jędryczka, M. Economic value of oilseed rape cultivars under production conditions with full fungicide protection technology using the SPEC forecasting system. Oilseed Crop. 2010, 31, 67–83. [Google Scholar]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Kaczmarek, J.; Jedryczka, M. Characterization of two coexisting pathogen populations of Leptosphaeria spp., the cause of stem canker of brassicas. Acta Agrobot. 2011, 64, 3–14. [Google Scholar] [CrossRef]
- Kaczmarek, J. Rozwój Stadium Generatywnego Grzybów Leptosphaeria maculans ([Desm.] Ces. et de Not.) i L. biglobosa (Shoemaker i Brun 2001) Oraz Ochrona Rzepaku Przed Tymi Patogenami. Ph.D. Thesis, Institute of Plant Genetics PAS, Poznan, Poland, 2010. [Google Scholar]
- Jedryczka, M. Epidemiology and damage caused by stem canker of oilseed rape in Poland. Phytopathol. Pol. 2007, 45, 73–75. [Google Scholar]
- Soxhlet, F. Die gewichtsanalytische Bestimmung des Milchfettes. Dinglers Polytech. J. 1879, 232, 461–465. [Google Scholar]
- Kjeldahl, J. Neue Methode zur Bestimmung des Stickstoffs in organischen Körpern. Fresenius Zeitschriftf. Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef] [Green Version]
- Sprague, S.J.; Kirkegaard, J.A.; Howlett, B.J.; Graham, J. Effect of root rot and stem canker caused by Leptosphaeria maculans on yield of Brassica napus and measures for control in the field. Crop Pasture Sci. 2010, 61, 50–58. [Google Scholar] [CrossRef]
- Zamani-Noor, N.; Knüfer, J. Effects of host plant resistance and fungicide application on phoma stem canker, growth parameters and yield of winter oilseed rape. Crop Prot. 2018, 112, 313–321. [Google Scholar] [CrossRef]
- Huang, Y.-J.; Qi, A.; King, G.J.; Fitt, B.D. L Assessing quantitative resistance against Leptosphaeria maculans (phoma stem canker) in Brassica napus (oilseed rape) in young plants. PLoS ONE 2014, 9, e84924. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek, J.; Brachaczek, A.; Jedryczka, M. The effect of fungicide spray time on the incidence of stem canker of Brassicas and seed yield of winter oilseed rape in Pomerania. J. Plant Dis. Prot. 2014, 121, 58–63. [Google Scholar] [CrossRef]
- Sewell, T.R.; Hawkins, N.J.; Stotz, H.U.; Huang, Y.-J.; Kelly, S.L.; Kelly, D.E. Azole sensitivity in Leptosphaeria pathogens of oilseed rape: The role of lanosterol 14α-demethylase. Sci. Rep. 2017, 7, 15849. [Google Scholar] [CrossRef] [PubMed]
- Jajor, E.; Mrówczyński, M.; Bartkowiak-Broda, I.; Broniarz, J.; Danielewicz, J.; Dobrzycka, A.; Dworzańska, D.; Fiedler, Ż.; Gorzała, G.; Horoszkiewicz-Janka, J.; et al. Metodyka Integrowanej Ochrony i Produkcji Rzepaku Ozimego Oraz Jarego Dla Doradców; IOR–PIB: Poznań, Poland, 2017; pp. 1–264. [Google Scholar]
- Pińskwar, I.; Choryński, A.; Kundzewicz, Z.W. Severe drought in the spring of 2020 in Poland—More of the same? Agronomy 2020, 10, 1646. [Google Scholar] [CrossRef]
- McCartney, H.A.; Doughty, K.J.; Norton, G.; Booth, E.J.; Kightley, S.P.J.; Landon, G.; West, G.; Walker, K.C.; Thomas, J.E. A study of the effect of disease on seed quality parameters of oilseed rape. In Proceedings of the 10th International Rapeseed Congress, Canberra, Australia, 26–29 September 1999; Available online: http://www.regional.org.aa/au/gcirc/index.html (accessed on 5 June 2021).
- Wenda-Piesik, A.; Hoppe, S. Evaluation of hybrid and population cultivars on standard and high-input technology in winter oilseed rape. Acta Agric. Scand. Sect. B Soil Plant Sci. 2018, 68, 678–689. [Google Scholar]
- Jankowski, K.J.; Sokólski, M.; Szatkowski, A. The effect of autumn foliar fertilization on the yield and quality of winter oilseed rape seeds. Agronomy 2019, 9, 849. [Google Scholar] [CrossRef] [Green Version]
- Gugała, M.; Sikorska, A.; Zarzecka, K. The effect of fertilization with sulphur, boron, and amino acids on the content of glucosinolate in winter rape seeds. Agronomy 2020, 10, 519. [Google Scholar] [CrossRef] [Green Version]
- Varga, L.; Ložek, O.; Ducsay, L.; Kováčik, P.; Lošák, T.; Hlušek, J. Effect of top dressing with nitrogen and boron on the yield and quality of rapeseed. Acta Univ. Agric. Silvic. Mendel. Brun. 2014, 58, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Baylis, A.D.; Hutley-Bull, P.D. The effects of a paclobutrazol based growth regulator on the yield, quality and ease of management of oilseed rape. Ann. Appl. Biol. 1991, 118, 445–452. [Google Scholar] [CrossRef]
- Poslušná, J.; Plachká, E.; Horáček, J.; Macháčková, I.; Ondráčková, E.; Šmirous, P., Jr.; Vrbovský, V. The harmfulness of phoma stem canker, Sclerotinia stem rot, and phytoplasma on winter oilseed rape with regard to Czech breeding programs. Agronomy 2019, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Stonard, J.F.; Marchant, B.P.; Latunde-Dada, A.O.; Liu, Z.; Evans, N.; Gladders, P.; Eckert, M.R.; Fitt, B.D.L. Geostatistical analysis of the distribution of Leptosphaeria species causing phoma stem canker on winter oilseed rape (Brassica napus) in England. Plant Pathol. 2010, 59, 200–210. [Google Scholar] [CrossRef] [Green Version]
- Jedryczka, M.; Rouxel, T.; Balesdent, M.H.; Mendes Pereira, E.; Bertrandy, J. Molecular characterization of Polish Phoma lingam isolates. Cereal Res. Commun. 1997, 25, 279–283. [Google Scholar] [CrossRef]
- Sewell, T.; Moloney, S.; Ashworth, M.; Ritchie, F.; Mashanova, A.; Huang, Y.-J.; Stotz, H.; Fitt, B.D.L. Effects of a penthiopyrad and picoxystrobin fungicide mixture on phoma stem canker (Leptosphaeria spp.) on UK winter oilseed rape. Eur. J. Plant Pathol 2016, 145, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.J.; Hood, J.; Eckert, M.R.; Stonard, J.F.; Cools, H.J.; King, G.J.; Rosall, S.; Ashworth, F.; Fitt, B.D.L. Effects of fungicide on growth of Leptosphaeria maculans and L. biglobosa in relation to development of phoma stem canker on oilseed rape (Brassica napus). Plant Path. 2011, 60, 607–620. [Google Scholar] [CrossRef] [Green Version]
- Draskau, M.K.; Boberg, J.; Taxvig, C.; Pedersen, M.; Frandsen, H.L.; Christiansen, S.; Svingen, T. In vitro and in vivo endocrine disrupting effects of the azole fungicides triticonazole and flusilazole. Environ. Pollut. 2019, 255, 113309. [Google Scholar] [CrossRef] [PubMed]
- Aubertot, J.N.; West, J.S.; Bousset-Vaslin, L.; Salam, M.U.; Barbetti, M.J.; Diggle, A.J. Improved resistance management for durable disease control: A case study of phoma stem canker of oilseed rape (Brassica napus). Eur. J. Plant Pathol. 2006, 114, 91–106. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Applications | Autumn 2009 | Autumn 2010 | Autumn 2011 | Autumn 2012 | ||||
|---|---|---|---|---|---|---|---|---|
| Glubczyce | Toszek | Glubczyce | Toszek | Glubczyce | Toszek | Glubczyce | Toszek | |
| Sowing | 19 Aug. | 21 Aug. | 24 Aug. | 19 Aug. | 16 Aug. | 19 Aug. | 23 Aug. | 24 Aug. |
| Spray 1 * | 30 Sept. | 30 Sept. | 24 Sept. | 23 Sept. | 27 Sept. | 24 Sept. | 17 Sept. | 20 Sept. |
| Spray 2 | 9 Oct. | 6 Oct. | 1 Oct. | 30 Sept. | 4 Oct. | 30 Sept. | 24 Sept. | 27 Sept. |
| Spray 3 | 14 Oct. | 15 Oct. | 7 Oct. | 7 Oct. | 11 Oct. | 6 Oct. | 30 Sept. | 3 Oct. |
| Spray 4 | 22 Oct. | 22 Oct. | 14 Oct. | 15 Oct. | 19 Oct. | 14 Oct. | 8 Oct. | 10 Oct. |
| Spray 5 | 29 Oct. | 31 Oct. | 22 Oct. | 21 Oct. | 25 Oct. | 20 Oct. | 15 Oct. | 15 Oct. |
| Spray 6 | 5 Nov. | 6 Nov. | 29 Oct. | 28 Oct. | 2 Nov. | 27 Oct. | 24 Oct. | 24 Oct. |
| Spray 7 | 13 Nov. | 13 Nov. | 4 Nov. | 5 Nov. | 8 Nov. | 4 Nov. | 30 Oct. | 30 Oct. |
| Spray 8 ** | 19 Nov. | 19 Nov. | 12 Nov. | 12 Nov. | 16 Nov. | 11 Nov. | 6 Nov. | 6 Nov. |
| Vegetative Season | Phoma Leaf Spot | Stem Canker | ||||
| L. maculans (%) | L. biglobosa (%) | No. of Isolates | L. maculans (%) | L. biglobosa (%) | No. of Isolates | |
| Glubczyce | ||||||
| 2010/2011 | 72.4 | 27.6 | 29 | 20.0 | 80.0 | 15 |
| 2011/2012 | 68.4 | 31.6 | 38 | 20.0 | 80.0 | 20 |
| 2012/2013 | 78.6 | 21.4 | 14 | 42.9 | 57.1 | 7 |
| 2013/2014 | 78.9 | 21.1 | 19 | 33.3 | 66.7 | 36 |
| Toszek | ||||||
| 2010/2011 | 66.67 | 33.33 | 6 | 31.6 | 68.4 | 19 |
| 2011/2012 | 75.00 | 25.00 | 12 | 21.4 | 78.6 | 14 |
| 2012/2013 | 80.00 | 20.00 | 20 | 35.7 | 64.3 | 14 |
| 2013/2014 | 77.78 | 22.22 | 9 | 15.0 | 85.0 | 20 |
| Fungicide Application | 2009/2010 | 2010/2011 | 2011/2012 | 2012/2013 | ||||
|---|---|---|---|---|---|---|---|---|
| 2009 | 2010 | 2010 | 2011 | 2011 | 2012 | 2012 | 2013 | |
| Phoma Leaf Spot | Stem Canker | Phoma Leaf Spot | Stem Canker | Phoma Leaf Spot | Stem Canker | Phoma Leaf Spot | Stem Canker | |
| Glubczyce | ||||||||
| Spray 1 | 15.08 bc* | 18.30 ab | 4.66 ab | 8.00 ab | 6.33 a | 7.01 a | 8.00 ab | 3.00 a |
| Spray 2 | 7.87 a | 14.23 a | 0.67 a | 6.67 ab | 7.80 ab | 9.39 a | 6.67 a | 4.33 a |
| Spray 3 | 11.07 ab | 18.30ab | 4.67 ab | 4.00 a | 9.29 abc | 9.28 a | 6.67 a | 4.33 a |
| Spray 4 | 17.25 c | 18.63 b | 9.33 bc | 4.00 a | 11.85 bcd | 13.22 b | 10.00 ab | 5.67 a |
| Spray 5 | 29.66 c | 18.30 ab | 15.33 cd | 4.00 a | 7.37 ab | 22.37 d | 10.67 ab | 7.00 a |
| Spray 6 | 35.92 d | 23.04 c | 29.33 e | 4.00 a | 11.59 bcd | 16.27 bc | 13.00 b | 21.33 b |
| Spray 7 | 35.24 d | 27.11 cd | 26.67 de | 5.33 a | 14.55 d | 19.55 cd | 9.67 ab | 18.67 b |
| Spray 8 | 34.33 cdc | 27.79 d | 30.00 e | 10.00 b | 13.52 cd | 18.52 c | 21.00 c | 21.00 b |
| Control | 29.63 c | 30.17 d | 30.67 e | 10.67 b | 17.34 e | 26.41 e | 22.00 c | 22.00 b |
| Toszek | ||||||||
| Spray 1 | 9.00 ab | 4.33 abc | 11.67 cd | 11.30 de | 3.10 a | 5.47 a | 3.67 ab | 5.67 ab |
| Spray 2 | 7.00 a | 2.67 a | 8.22 b | 5.86 abc | 4.57 a | 7.04 ab | 2.34 a | 4.34 ab |
| Spray 3 | 7.00 a | 4.00 ab | 2.14 a | 3.14 a | 4.90 a | 6.94 ab | 2.34 a | 1.67 a |
| Spray 4 | 8.00 a | 8.00 d | 2.11 a | 7.29 bc | 8.62 b | 10.88 bcd | 5.67 abc | 1.67 a |
| Spray 5 | 5.67 a | 8.67 de | 5.56 b | 4.55 a | 10.33 bc | 10.59 bc | 6.34 abcd | 1.67 a |
| Spray 6 | 12.03 bc | 6.00 bcd | 11.22 c | 9.57 cd | 9.79 bc | 12.15 cd | 8.02 cde | 1.67 a |
| Spray 7 | 8.00 a | 7.00 d | 20.48 e | 17.43 f | 11.32 c | 12.80 cd | 6.5 bcd | 3.00 a |
| Spray 8 | 13.00 c | 11.00 ef | 12.01 cd | 12.01 de | 10.29 bc | 14.67 d | 10.33 de | 7.67 b |
| Control | 15.00 c | 13.33 f | 14.03 d | 14.44 ef | 10.11 bc | 24.07 e | 10.67 e | 14.34 d |
| Fungicide Application | Yield (dt ha−1) | Oil Content (%) | Protein Content (%) | Mass of Thousand Seeds (g) |
|---|---|---|---|---|
| Autumn 2009 | ||||
| Spray 1 | 44.62 a* | 47.82 b | 21.10 ab | 4.08 ab |
| Spray 2 | 50.83 a | 47.82 b | 21.50 b | 4.10 ab |
| Spray 3 | 50.33 a | 47.69 b | 21.70 b | 4.06 ab |
| Spray 4 | 48.30 a | 47.80 b | 21.33 ab | 4.02 a |
| Spray 5 | 50.51 a | 48.12 b | 21.12 ab | 4.18 b |
| Spray 6 | 47.78 a | 47.64 b | 21.10 ab | 4.12 ab |
| Spray 7 | 46.65 a | 47.55 ab | 21.34 ab | 4.02 a |
| Spray 8 | 45.79 a | 47.51 ab | 21.40 ab | 4.08 ab |
| Control | 47.04 a | 46.01 a | 20.53 a | 4.03 a |
| Autumn 2010 | ||||
| Spray 1 | 54.15 a | 47.41 a | 21.30 a | 5.08 ab |
| Spray 2 | 56.08 a | 47.60 a | 21.70 a | 4.88 ab |
| Spray 3 | 54.16 a | 47.64 a | 21.90 a | 4.70 a |
| Spray 4 | 54.56 a | 47.29 a | 21.53 a | 5.28 ab |
| Spray 5 | 54.75 a | 47.22 a | 21.32 a | 5.30 ab |
| Spray 6 | 54.57 a | 47.02 a | 21.30 a | 5.06 ab |
| Spray 7 | 55.07 a | 46.36 a | 21.39 a | 5.26 ab |
| Spray 8 | 54.67 a | 46.29 a | 21.31 a | 5.48 b |
| Control | 54.25 a | 46.12 a | 21.41 a | 5.00 ab |
| Autumn 2011 | ||||
| Spray 1 | 41.45 a | 47.82 a | 19.51 ab | 4.42 ab |
| Spray 2 | 42.24 a | 48.57 a | 20.22 ab | 4.38 a |
| Spray 3 | 39.18 a | 47.57 a | 20.51 b | 4.34 a |
| Spray 4 | 41.04 a | 47.15 a | 19.70 ab | 4.50 b |
| Spray 5 | 41.19 a | 47.33 a | 19.40 ab | 4.44 ab |
| Spray 6 | 40.78 a | 47.73 a | 19.24 a | 4.34 a |
| Spray 7 | 39.65 a | 46.46 a | 19.34 ab | 4.40 ab |
| Spray 8 | 38.79 a | 48.16 a | 19.63 ab | 4.40 ab |
| Control | 38.63 a | 46.11 a | 19.94 ab | 4.3 a |
| Autumn 2012 | ||||
| Spray 1 | 44.73 d | 47.71 a | 19.57 a | 4.40 a |
| Spray 2 | 44.27 d | 47.71 a | 19.58 a | 4.59 ab |
| Spray 3 | 42.60 c | 47.58 a | 19.26 a | 4.70 abc |
| Spray 4 | 41.60 c | 47.71 a | 19.1 a | 4.86 abc |
| Spray 5 | 40.17 b | 48.01 a | 19.18 a | 5.01 abc |
| Spray 6 | 41.50 cd | 47.53 a | 19.36 a | 5.03 abc |
| Spray 7 | 40.03 b | 47.44 a | 19.30 a | 5.16 bc |
| Spray 8 | 39.93 b | 47.40 a | 18.9 a | 5.29 c |
| Control | 37.10 a | 47.37 a | 18.78 a | 5.29 bc |
| Fungicide Application | Yield (dt ha−1) | Oil Content (%) | Protein Content (%) | Mass of Thousand Seeds (g) |
|---|---|---|---|---|
| Autumn 2009 | ||||
| Spray 1 | 36.56 a | 44.33 a | 21.83 c | 3.94 a |
| Spray 2 | 42.77 a | 45.03 a | 20.14 b | 3.99 a |
| Spray 3 | 42.94 a | 44.60 a | 19.89 ab | 3.81 a |
| Spray 4 | 41.64 a | 45.60 a | 19.65 ab | 3.72 a |
| Spray 5 | 42.45 a | 45.33 a | 19.54 ab | 3.66 a |
| Spray 6 | 39.72 a | 45.80 a | 19.50 ab | 3.68 a |
| Spray 7 | 38.59 a | 46.60 a | 18.64 ab | 3.69 a |
| Spray 8 | 37.73 a | 45.77 a | 18.55 a | 3.86 a |
| Control | 37.57 a | 44.70 a | 19.32 ab | 4.07 a |
| Autumn 2010 | ||||
| Spray 1 | 41.23 c | 44.70 a | 19.08 ab | 4.62 ab |
| Spray 2 | 42.21 c | 46.63 a | 18.75 ab | 4.57 ab |
| Spray 3 | 41.00 c | 46.23 a | 19.22 ab | 4.49 ab |
| Spray 4 | 41.07 c | 45.50 a | 18.86 ab | 4.47 ab |
| Spray 5 | 37.67 a | 45.17 a | 18.16 a | 4.28 a |
| Spray 6 | 37.00 a | 45.87 a | 18.96 ab | 4.33 ab |
| Spray 7 | 40.53 bc | 45.70 a | 19.65 b | 4.39 ab |
| Spray 8 | 40.00 b | 47.10 a | 18.42 a | 4.66 ab |
| Control | 37.00 a | 44.13 a | 18.44 a | 5.02 a |
| Autumn 2011 | ||||
| Spray 1 | 40.36 a | 47.20 a | 18.46 a | 4.76 c |
| Spray 2 | 41.15 a | 48.02 a | 21.14 cde | 4.74 c |
| Spray 3 | 38.09 a | 48.26 a | 21.30 de | 4.60 bc |
| Spray 4 | 39.95 a | 48.62 a | 21.29 de | 4.79 c |
| Spray 5 | 40.10 a | 48.48 a | 21.83 e | 4.31 abc |
| Spray 6 | 39.69 a | 48.21 a | 20.14 cd | 3.60 ab |
| Spray 7 | 38.56 a | 48.21 a | 19.89 abcd | 3.40 a |
| Spray 8 | 37.70 a | 47.57 a | 19.65 abc | 3.62 ab |
| Control | 37.54 a | 46.44 a | 19.50 ab | 3.42 a |
| Autumn 2012 | ||||
| Spray 1 | 40.00 bc | 45.79 a | 18.64 a | 4.93 ab |
| Spray 2 | 41.47 c | 45.50 a | 19.59 a | 4.84 ab |
| Spray 3 | 42.07 c | 44.20 a | 19.53 a | 4.89 ab |
| Spray 4 | 40.21 bc | 46.53 a | 19.24 a | 4.90 a |
| Spray 5 | 41.57 c | 47.37 a | 18.41a | 5.14 ab |
| Spray 6 | 40.90 c | 46.40 a | 18.97 a | 5.27 ab |
| Spray 7 | 40.00 bc | 46.73 a | 19.24 a | 5.39 c |
| Spray 8 | 38.00 ab | 46.50 a | 19.52 a | 5.37 b |
| Control | 37.30 a | 47.37 a | 18.90 a | 4.62 a |
| Indole Glucosinolate | Alkenyl Glucosinolate | |||||
|---|---|---|---|---|---|---|
| Fungicide Application | 4-Hydroxyglucobrassicine (μM/g) | Gucobrassicine (μM/g) | Glucobrassicanapine (μM/g) | Gluconapine (μM/g) | Napoleiferine (μM/g) | Progoitrine (μM/g) |
| Autumn 2009 | ||||||
| Spray 1 | 4.70 b | 0.12 c | 0.42 b | 1.41 c | 0.11 b | 2.76 cd |
| Spray 2 | 4.45 ab | 0.15 d | 0.57 b | 1.47 c | 0.12 c | 2.28 ab |
| Spray 3 | 4.17 a | 0.19 e | 0.24 a | 2.40 f | 0.13 c | 2.85 de |
| Spray 4 | 4.74 b | 0.23 f | 0.82 c | 2.10 e | 0,13 c | 2.42 abcd |
| Spray 5 | 4.61 ab | 0.087 b | 0.83 c | 1.51 c | 0.10 b | 2.39 abc |
| Spray 6 | 4.72 b | 0.11 bc | 0.60 b | 1.80 d | 0.09 b | 2.66 bcd |
| Spray 7 | 4.50 ab | 0.16 d | 0.20 a | 0.12 a | 0.13 a | 2.01 a |
| Spray 8 | 4.08 a | 0.16 d | 0.23 a | 0.59b | 0.02 a | 2.33 abc |
| Control | 4.08 a | 0.056 a | 0.99 d | 1.84 d | 0.18 d | 3.25 e |
| Autumn 2010 | ||||||
| Spray 1 | 4.30 a | 0.11 a | 0.80 abc | 1.37 ab | 0.12 bcd | 3.41 bc |
| Spray 2 | 4.08 a | 0.10 a | 0.90 abc | 1.43 b | 0.11 bc | 3.30 abc |
| Spray 3 | 4.55 a | 0.11 a | 1.00 abc | 2.33 e | 0.13 d | 2.68 ab |
| Spray 4 | 4.61 a | 0.12 a | 1.30 cd | 2.06 de | 0.12 bcd | 3.86 c |
| Spray 5 | 4.57a | 0.15 a | 1.10 bcd | 1.47 bc | 0.10 b | 3.52 c |
| Spray 6 | 4.48 a | 0.11 a | 0.50 a | 1.76 cd | 0.12 bcd | 3.59 c |
| Spray 7 | 4.62 a | 0.13 a | 1.10 bcd | 1.41 ab | 0.13 cd | 2.60 a |
| Spray 8 | 4.23 a | 0.13 a | 0.65 ab | 1.10 a | 0.024 a | 4.01 c |
| Control | 3.93 a | 0.10 a | 1.67 d | 1.80 d | 0.17 e | 3.84 c |
| Autumn 2011 | ||||||
| Spray 1 | 4.29 a | 0.16 ab | 0.50 a | 1.98 ab | 0.11 cd | 2.93 c |
| Spray 2 | 4.08 a | 0.23 b | 1.13b | 1.71 b | 0.10 bc | 3.20 d |
| Spray 3 | 4.55 a | 0.11 a | 0.92 ab | 1.71 b | 0.09 b | 3.00 cd |
| Spray 4 | 4.61 a | 0.13 ab | 1.80 c | 1.35 ab | 0.13 ef | 1.20 a |
| Spray 5 | 4.58 a | 0.15 ab | 1.81 c | 0.76 a | 0.09 a | 2.10 b |
| Spray 6 | 4.48 a | 0.17 ab | 0.89 ab | 0.50 a | 0.10 bc | 3.20 d |
| Spray 7 | 4.62 a | 0.14 ab | 0.93 ab | 0.71 a | 0.12 def | 3.15 cd |
| Spray 8 | 4.23 a | 0.11 a | 1.10 b | 0.80 a | 0.13 f | 2.10 b |
| Control | 3.93 a | 0.09 a | 1.09 b | 1.80 b | 0.11 cde | 1.00 a |
| Autumn 2012 | ||||||
| Spray 1 | 4.26 ab | 0.17 c | 0.42 b | 1.31 b | 0.06 ab | 2.61 c |
| Spray 2 | 4.45 b | 0.15 bc | 0.57 c | 1.37 bc | 0.06 a | 2.14 a |
| Spray 3 | 4.60 a | 0.15 bc | 0.24 a | 2.27 e | 0.07 ab | 2.70 cd |
| Spray 4 | 4.10 ab | 0.19 c | 0.43 b | 1.99 de | 0.08 abc | 2.27 ab |
| Spray 5 | 3.90 a | 0.23 d | 0.76 f | 1.41 bc | 0.08 abc | 2.24 ab |
| Spray 6 | 4.50 a | 0.09 a | 0.60 cd | 1.70 cd | 0.08 abc | 2.51 bc |
| Spray 7 | 3.90 a | 0.11 ab | 0.65 de | 0.62 a | 0.08 abc | 2.21 ab |
| Spray 8 | 4.50 a | 0.15 ab | 0.67 e | 0.48 a | 0.09 bc | 2.15 a |
| Control | 3.80 a | 0.15 bc | 0.83 g | 0.63 a | 0.10 c | 2.96 d |
| Fungicide Application | Indole Glucosinolate | Alkenyl Glucosinolate | ||||
|---|---|---|---|---|---|---|
| 4-Hydroxyglucobrassicine (μM/g) | Glucobrassicine (μM/g) | Glucobrassicanapine (μM/g) | Gluconapine (μM/g) | Napoleiferine (μM/g) | Progoitrine (μM/g) | |
| Autumn 2009 | ||||||
| Spray 1 | 3.67 a | 0.15 a | 0.57 a | 1.47 c | 0.10 a | 5.71 a |
| Spray 2 | 3.59 a | 0.15 a | 0.62 a | 2.37 e | 0.10 a | 6.23 a |
| Spray 3 | 3.61 a | 0.16 a | 0.58 a | 2.10 de | 0.10 a | 7.10 a |
| Spray 4 | 3.54 a | 0.16 a | 0.57 a | 1.51 c | 0.097 a | 6.21 a |
| Spray 5 | 3.51 a | 0.16 a | 0.59 a | 1.80 cd | 0.097 a | 7.46 a |
| Spray 6 | 3.69 a | 0.17 a | 0.62 a | 0.12 a | 0.11 a | 6.70 a |
| Spray 7 | 3.53 a | 0.17 a | 0.52 a | 0.59 ab | 0.10 a | 6.80 a |
| Spray 8 | 3.59 a | 0.17 a | 0.60 a | 0.72 b | 0.11 a | 6.11 a |
| Control | 3.75 a | 0.18 a | 0.56 a | 1.33 c | 0.10 a | 7.87 a |
| Autumn 2010 | ||||||
| Spray 1 | 3.67 a | 0.17 a | 0.60 a | 2.10 d | 0.11 a | 4.75 ab |
| Spray 2 | 3.59 a | 0.16 a | 0.65 a | 1.51 c | 0.11 a | 5.26 ab |
| Spray 3 | 3.61 a | 0.16 a | 0.59 a | 1.80 cd | 0.12 a | 3.65 a |
| Spray 4 | 3.54 a | 0.15 a | 0.58 a | 0.12 a | 0.11 a | 4.85 ab |
| Spray 5 | 3.51 a | 0.16 a | 0.58 a | 0.59 ab | 0.12 a | 5.49 ab |
| Spray 6 | 3.69 a | 0.17 a | 0.53 a | 0.72 b | 0.10 a | 4.43 ab |
| Spray 7 | 3.53 a | 0.18 a | 0.44 a | 1.40 c | 0.12 a | 4.71 ab |
| Spray 8 | 3.59 a | 0.17 a | 0.51 a | 1.60 cd | 0.11 a | 5.20 ab |
| Control | 3.75 a | 0.15 a | 0.45 a | 1.64 cd | 0.11 a | 7.11 b |
| Autumn 2011 | ||||||
| Spray 1 | 3.51 a | 0.17 a | 0.45 a | 1.60 ab | 0.10 a | 3.15 a |
| Spray 2 | 3.56 a | 0.18 a | 0.48 a | 1.60 ab | 0.10 a | 3.72 a |
| Spray 3 | 3.67 a | 0.17 a | 0.37 a | 1.53 a | 0.11 a | 3.18 a |
| Spray 4 | 3.90 a | 0.17 a | 0.46 a | 1.85 ab | 0.11 a | 3.94 a |
| Spray 5 | 3.60 a | 0.15 a | 0.52 a | 1.36 a | 0.11 a | 2.49 a |
| Spray 6 | 3.58 a | 0.15 a | 0.40 a | 1.45 a | 0.11 a | 2.75 a |
| Spray 7 | 3.62 a | 0.15 a | 0.52 a | 1.65 ab | 0.11 a | 3.16 a |
| Spray 8 | 3.75 a | 0.15 a | 0.68 a | 1.41 a | 0.11 a | 2.37 a |
| Control | 3.75 a | 0.17 a | 0.66 a | 2.15 b | 0.11 a | 5.24 b |
| Autumn 2012 | ||||||
| Spray 1 | 3.65 a | 0.18 ab | 0.59 a | 1.40 a | 0.11 a | 5.16 a |
| Spray 2 | 3.83 a | 0.20 b | 0.58 a | 2.52 a | 0.11 a | 5.14 a |
| Spray 3 | 3.77 a | 0.18 ab | 0.61 a | 2.59 a | 0.12 a | 4.87 a |
| Spray 4 | 3.84 a | 0.09 a | 0.67 a | 2.52 a | 0.12 a | 4.48 a |
| Spray 5 | 3.38 a | 0.12 ab | 0.61 a | 2.13 a | 0.12 a | 5.20 a |
| Spray 6 | 3.50 a | 0.16 ab | 0.56 a | 2.57 a | 0.12 a | 4.83 a |
| Spray 7 | 3.53 a | 0.15 ab | 0.57 a | 2.36 a | 0.12 a | 4.41 a |
| Spray 8 | 3.61 a | 0.15 ab | 0.50 a | 2.37 a | 0.11a | 4.27 a |
| Control | 4.17 a | 0.16 ab | 0.66 a | 2.81 a | 0.11a | 5.83 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brachaczek, A.; Kaczmarek, J.; Jedryczka, M. Warm and Wet Autumns Favour Yield Losses of Oilseed Rape Caused by Phoma Stem Canker. Agronomy 2021, 11, 1171. https://doi.org/10.3390/agronomy11061171
Brachaczek A, Kaczmarek J, Jedryczka M. Warm and Wet Autumns Favour Yield Losses of Oilseed Rape Caused by Phoma Stem Canker. Agronomy. 2021; 11(6):1171. https://doi.org/10.3390/agronomy11061171
Chicago/Turabian StyleBrachaczek, Andrzej, Joanna Kaczmarek, and Malgorzata Jedryczka. 2021. "Warm and Wet Autumns Favour Yield Losses of Oilseed Rape Caused by Phoma Stem Canker" Agronomy 11, no. 6: 1171. https://doi.org/10.3390/agronomy11061171
APA StyleBrachaczek, A., Kaczmarek, J., & Jedryczka, M. (2021). Warm and Wet Autumns Favour Yield Losses of Oilseed Rape Caused by Phoma Stem Canker. Agronomy, 11(6), 1171. https://doi.org/10.3390/agronomy11061171