3.1. Chemical Characterisation of Oils
Table 1 presents the chemical composition of the essential oil (EO) from
E. camaldulensis var obtusa and
Mentha piperita green aerial parts, where the main compounds in
E. camaldulensis var obtusa EO were eucalyptol (33.04%), spathulenol (21.15%),
p-cymene (10.49%),
γ-terpinene (6.55%), crypton (5.35%), phellandral (3.01%), thymol (2.69%) and terpinen-4-ol (2.45%), while the main compounds in
M. piperita were pulegone (29.38%), isomenthone (17.23%), levomenthol (16.36%), eucalyptol or 1,8-cineole (7.46%), menthone (6.90%), aromadendrene (3.64%), endo-borneol (3.15%) and piperitone (3.02%). Previously, the EO from
M. piperita leaves showed the presence of menthone, 1,8-cineole, menthyl acetate, caryophyllene,
β-pinene and D-limonene as main compounds with values of 20.18, 15.48, 13.13, 4.82, 4.37 and 2.81%, respectively [
19]. In addition, several studies reported that the EO from
Mentha species were composed of different chemical compositions, with the abundant compounds menthol, eucalyptol, menthone, limonene,
trans-carveol, pulegone,
β-caryophyllene and pipertitinone oxide [
52,
53,
54]. The Algerian
Mentha plant EO showed the presence of menthol, menthone, and menthyl acetate as major constituents [
55].
For the chemical composition of
M. oleifera seeds fixed oil (FO), the full chemical analysis can be found in our previous work [
32] as shown in
Table 2, where the main compounds were oleic acid,
β-sitosterol and
α-tocopherol (vitamin E) with percentages of 59.7, 21.4 and 4.9%, respectively, as measured by GC-MS. After the methylation of fatty acids (FAs), the main fatty acid methyl esters (FAMEs) were oleic, palmitic, stearic, linolenic and arachidic, with values of 78.72%, 6.27, 5.63, 3.72 and 3.29%, respectively. The main FAMEs from
M. oleifera oil were oleic and palmitic at percentages of 74.99 and 12.51%, respectively [
56]. FAs of oleic, palmitic, stearic and linoleic with values 77.40, 12.97, 2.95 and 1.40%, respectively, were found in moringa seed FO [
57].
3.2. Fruit Visual and Chemical Parameters
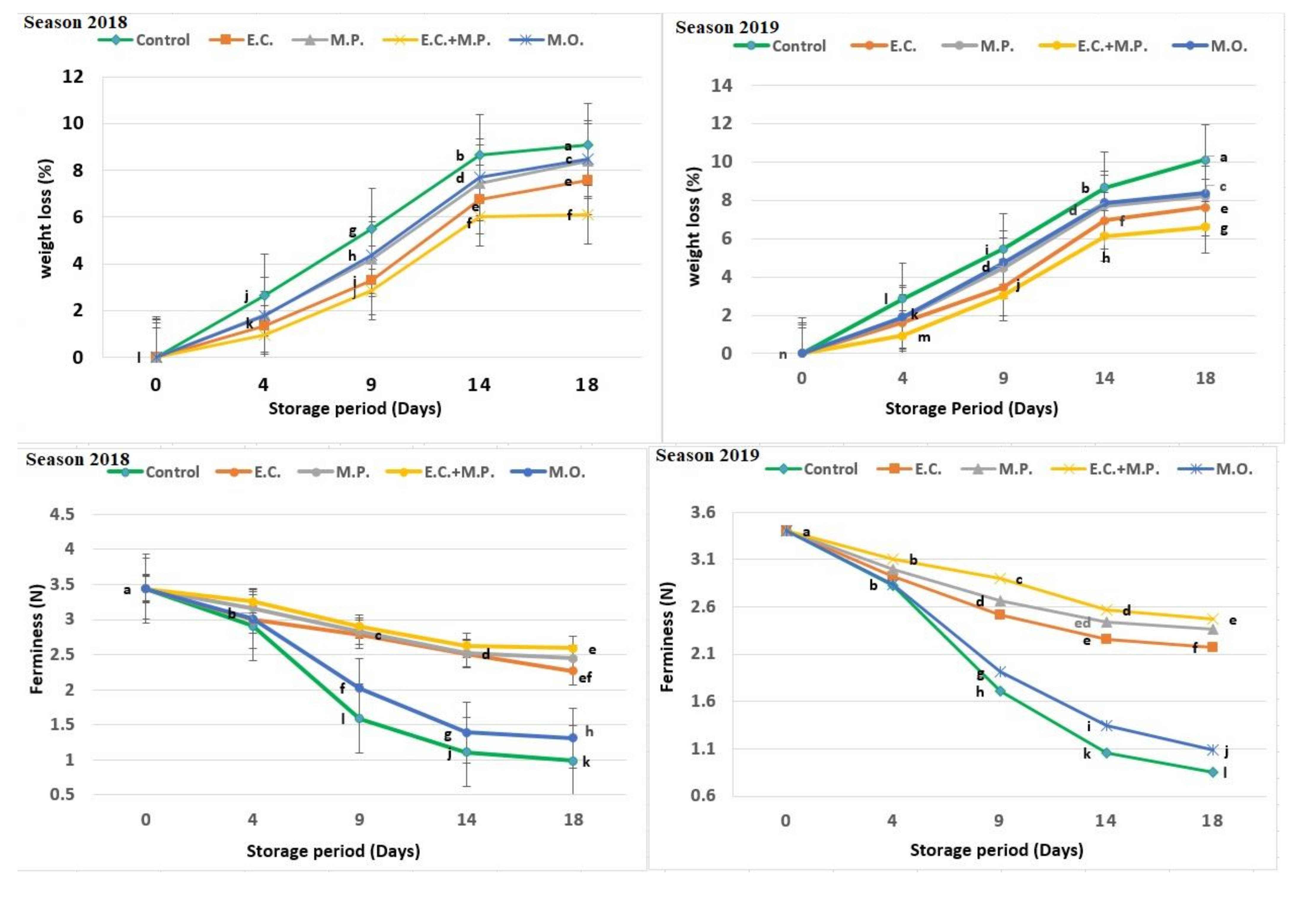
Figure 1 shows all the studied visual and chemical parameters of strawberry fruits treated with EOs and FO and different periods of storage. Significantly, there were different effects among natural oils and the storage days in all the studied parameters. The use of EOs observed the greatest effects in slowing down the weight loss, while moringa FO and the control sample showed the lowest effects. However, at the end of the storage period, a highly significant difference was found between treated and untreated strawberry samples after 18 days from the start of the storage period. The loss of weight in fresh fruit primarily reflects the respiration rate and moisture evaporation between the fruit tissue and surrounding air storage [
58], which are influenced by post-harvest treatment and storage temperature [
15]. Strawberry fruits are highly susceptible to a rapid loss of water due to the extremely thin skins of these fruits.
The effect of EO vapours could be associated with the formation of the fruit surface coating that modifies gas permeation and decreases respiration rate and water loss [
59]. The EO fumigation was shown to decrease the dehydration process in fruit [
60]. Our result was in agreement with a previous study on strawberries [
26,
41], cherries [
13], grapes [
61] and peaches [
59] treated with eugenol, thymol, menthol and cinnamon vapours. Strawberry fruits’ firmness is a very important visual quality of fresh market fruit in determining their post-harvest quality and shelf life [
41].
Generally, firmness decreased during storage conditions in both seasons. The EOs’ vapours also influenced the preservation of firmness of these fruits compared with untreated fruit (control) and fruit treated with moringa FO at 1%, which showed a significant decrease in firmness at 9, 14 and 18 days of storage, while vapour with mixed EOs from E. camaldulensis var obtusa and M. piperita (1/1 v/v) resulted in higher firmness than other treatments at the end of the storage period for both seasons.
Similar to our findings, EO vapours decreased the firmness losses during cold storage conditions in the strawberry [
26]. Furthermore, strawberry firmness was amended when lemon or orange EOs were used at shelf life storage of the fruit [
2]. The marked effect observed in previous studies might be related to storage. This was probably due to the selective permeability of the coating material to gas and water transmission, thus reducing respiration rates, enzyme activities and most of the metabolic changes, thereby delaying ripening and over softening of the strawberry [
62,
63,
64,
65].
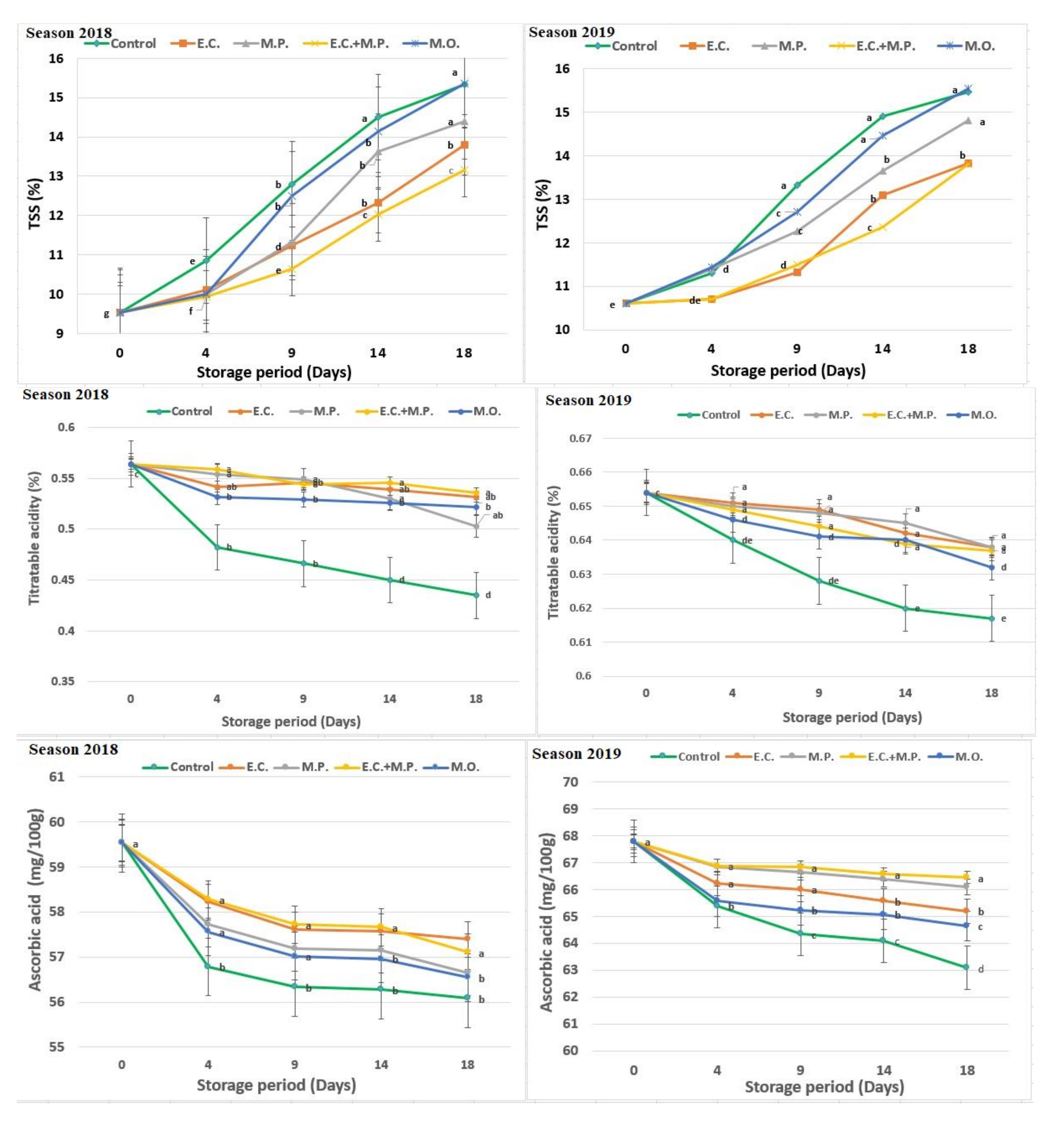
Total soluble solids (TSS%) were increased by time passed from harvest day in all treatments. At the end of the storage period, the lowest TSS was observed in fruits treated with mix of
E. camaldulensis var obtusa and
M. piperita EOs (1/1
v/
v). It seems that the increase in TSS was due to respiration during the storage period. EO fumigation reduced TSS consumption in strawberry fruits compared to control because of the decreased respiration as a result of reducing gas exchange [
2,
66,
67]. Other reasons for the increase in TSS are degradation of carbohydrate, other material changes such as acids, increasing soluble pectin and fruit corruption [
68,
69]. Furthermore, it can be observed that the TSS increase is also correlated to the weight loss.
The percentage of titratable acidity (TA) value was decreased through the storage period and the lowest value was recorded in the last four days of the storage period, where all treatments had a reduced acidity value compared to the control treatment. The lowest value of acidity was obtained due to E. camaldulensis var obtusa and M. piperita mix EOs (1/1 v/v), while the highest value was obtained with the control treatment in both seasons.
TA is an important factor in maintaining the quality of fruits, which is directly related to the organic acid content present in the fruit [
70,
71], and the decrease in TA content could be due to the consumption of organic acids in fruits during respiration. In a similar study, treatments have a significant effect on the respiration process, which could result in reduction or delay of respiration and results in maintenance of TA content [
58].
Ascorbic acid (AA) contents of all treated and untreated strawberry fruits were significantly decreased with the increase of storage periods from 0 to 18 days. This reduction could be related to its oxidation through superoxide and hydroxyl radicals in the strawberry fruits [
72]. However, at the end of the storage period, the concentrations of AA were higher in fruits treated with oils compared to the control (untreated fruits). The maximum values of AA were observed in the strawberry fruits treated with the mix of
E. camaldulensis var obtusa and
M. piperita EOs (1/1
v/
v) followed by
E. camaldulensis var obtusa EO and
M. piperita EO compared with untreated fruits (control). This result might be attributed to the antioxidant properties of the EOs, which reduce the diffusion of oxygen, decrease the rate of respiration and consequently reduce the AA oxidation [
62].
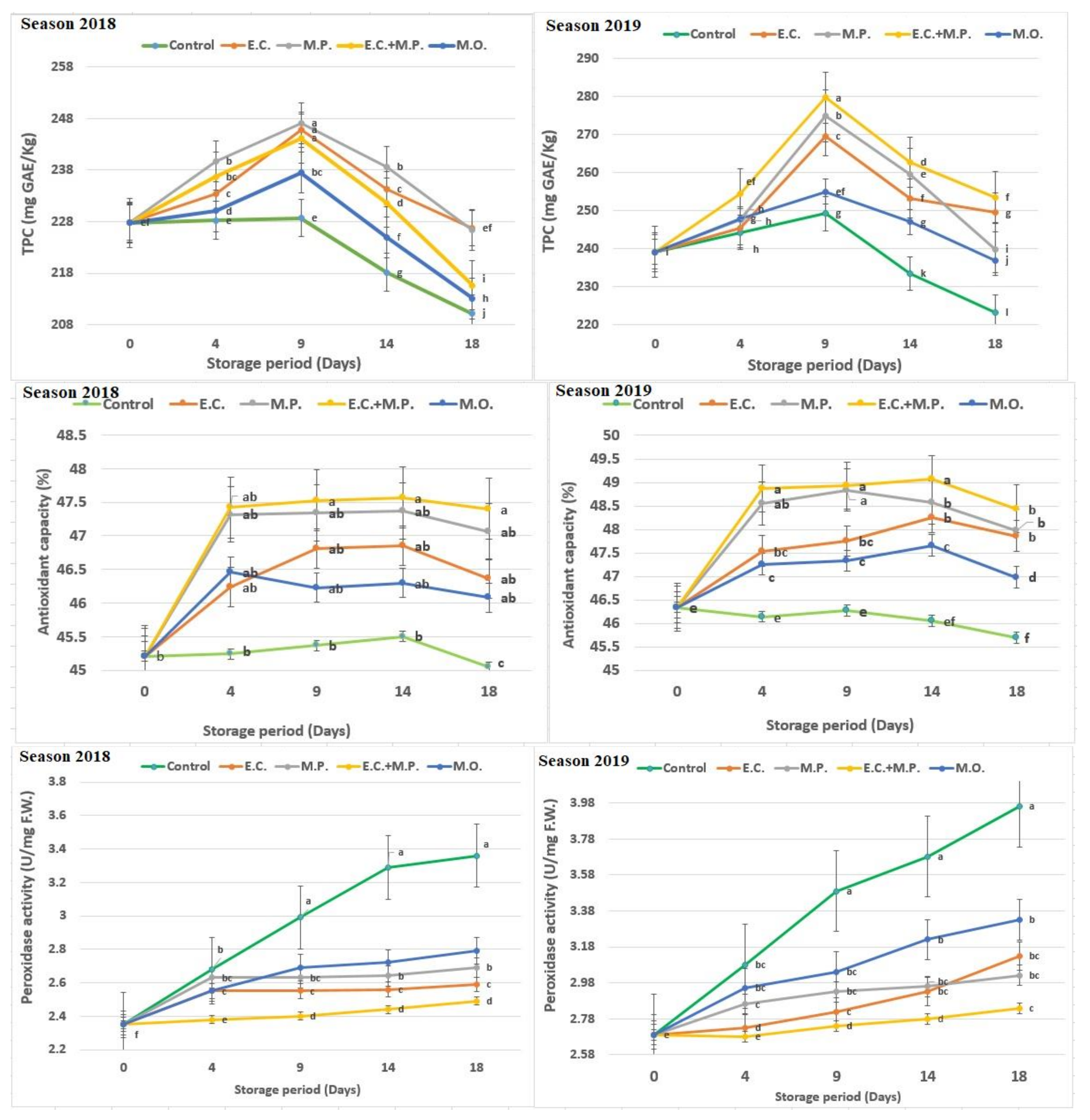
Total phenolic content (TPC, mg GAE/kg) continuously increased in the fruits treated with oils until 9 days from the start of the storage period, at which point the trend slowed down gradually and peaked on day 18. Mahmoud et al. [
73] found that the TPC of the Hollywood plum was increased during cold storage. Piljac-Žegarac and Šamec [
74] reported that small fruits like strawberries, raspberries, cherries and sour cherries stored at 4 °C exhibited slightly higher antioxidant activity values. Moreover, significant correlations between antioxidant capacity (AOC) and phenolic components in different fruits were established [
75,
76]. AOC was reduced during the storage time but not between treatments, and the changing trend in the control was more than with other treatments of the fruit. The fruits treated with
E. camaldulensis var obtusa and
M. piperita mix EOs (1/1
v/
v) and
E. camaldulensis var obtusa EO retained a higher AOC, reinforcing this attribute in the fruit at the end of the storage period in both seasons. During the storage period, the reduction in this property at the end of the storage time could be due to senescence and decomposition [
77]. In addition, AOC decreases due to cell protection against the damage caused by free radicals, where EOs decrease the respiration rate and free radical production by means of moisture maintenance and CO
2 and O
2 exchange control [
2,
25]. On the other hand, the AOC was maintained with a high percentage in the oil-treated fruits compared to the untreated fruits at 5 °C, with an increase in the storage period, whereas a previous report showed that the fruits stored at 10 °C had higher antioxidant enzyme activities and AOC than those stored at 0 or 5 °C [
78].
The peroxidase enzyme activity in all fruits increased with the extension of storage periods, and peroxidase in treated fruits with different EOs showed a lower level compared to control fruits. At the end of the storage period, the lowest peroxidase enzyme was in strawberries treated with the mix of
E. camaldulensis var obtusa and
M. piperita EOs (1/1
v/
v) followed by
E. camaldulensis var obtusa and
M. piperita EOs. Similar findings were reported by Badawy et al. [
79], who observed that the use of EOs containing thymol (0.02%) or geraniol (0.04%) increased catalase (CAT) activity and reduced polyphenol oxidase (PPO) activity and peroxidase. These results suggested that the use of EOs improves the levels of oxyradical detoxification enzymes including CAT and decreases the PPO activity that prevents the oxidation damage and peroxidase, thus promoting prolongation of the shelf-life and preserving the quality of strawberries during storage.
3.4. Phenolic Compounds of Oil-Treated Fruits by HPLC
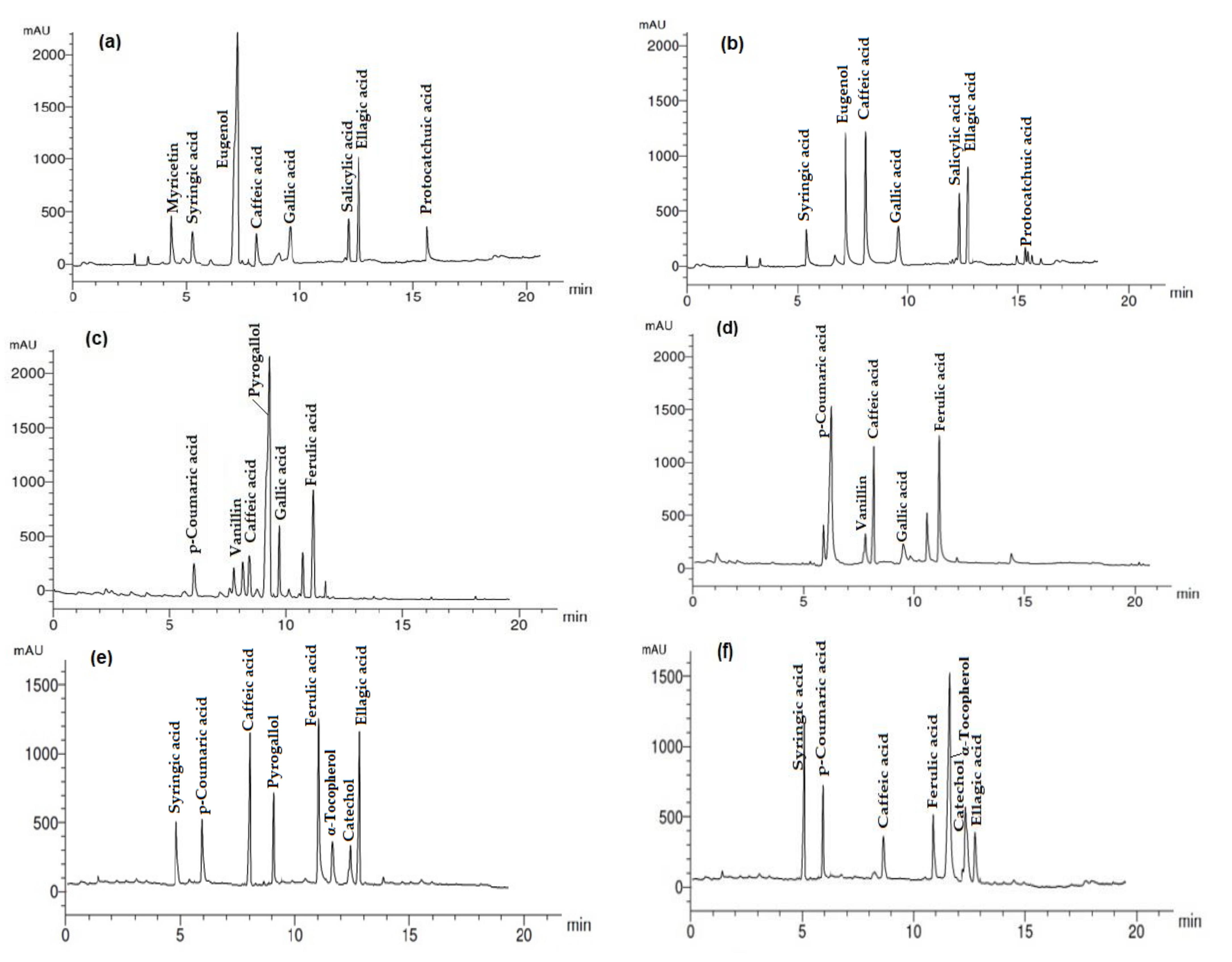
Figure 2 and
Table 4 show the changes in phenolic compounds in the methanol extract (ME) from the oil-treated or untreated strawberry fruits stored at 5 °C for 18 days compared to the original fruit sample (not stored). Myricetin was found only in the ME from the original fruit sample. Syringic acid ranged from 8.12 to 13.30 µg/mL,
p-coumaric acid from 8.09 to 25.51 µg/mL and eugenol was decreased from 35.16 µg/mL (original fruit) to 18.05 µg/mL (untreated fruit), but it was not detected in the ME from all the fruits treated with oils. Vanillin acid was detected only in the ME from fruits treated with the EOs from
E. camaldulensis var obtusa and
M. piperita. Caffeic acid was found in the ME of all fruit samples and ranged from 5.36 µg/mL (treated from with Moringa FO) to 19.63 µg/mL (treated fruits with the mixture of EOs from
E. camaldulensis var obtusa and
M. piperita (1/1
v/
v)).
p-Hydroxybenzoic acid was detected only in the ME from the fruits treated with
E. camaldulensis var obtusa EO (7.12 µg/mL). Pyrogallol was detected in the ME from fruits treated with
E. camaldulensis var obtusa EO (28.5 µg/mL) and a mix of
E. camaldulensis var obtusa and
M. piperita mix EOs (1/1
v/
v) (14.51 µg/mL).
Gallic acid ranged from 5.12 to 12.66 µg/mL but was not detected in fruits treated with a mix of E. camaldulensis var obtusa and M. piperita (1/1 v/v)) EOs or moringa FO. Ferulic acid ranged from 6.12 µg/mL to 21.12 µg/mL in the ME from fruits treated with M. oleifera FO and the mix of E. camaldulensis var obtusa and M. piperita EOs (1/1 v/v), respectively. A high amount of α-tocopherol was detected in the ME of fruits treated with M. oleifera FO (22.01 µg/mL) and in the fruits treated with a mix of EOs from E. camaldulensis var obtusa and M. piperita EOs (1/1 v/v) (7.45 µg/mL) and was not detected in the ME from other fruits. Salicylic acid was not detected in the ME from the fruits treated with oils, but it was detected in the original fruit sample (9.12 µg/mL) and the untreated fruit (9.56 µg/mL). Catechol was identified in the ME from fruits treated with mix of E. camaldulensis var obtusa and M. piperita EOs (1/1 v/v) (5.18 µg/mL) and M. oleifera FO (6.23 µg/mL).
According to the above results concerning the HPLC analysis of the changes of phenolic compounds, there are differences in the concentrations of the identified compounds among the fruits treated with EOs or FO. Phenolic compounds such as quercetin and kaempferol glycosides,
p-coumaric acid and ellagic acid were identified in strawberry fruits [
80,
81,
82,
83]. It was reported that ellagic acid was the most abundant phenolic compound in the strawberry [
80,
84], but it was found strictly as a compound or combined with glycosides and ellagitannins [
85].
p-OH benzoic acid,
p-coumaric acid glucoside, ferulic acid derivative and caffeic acid were identified in strawberry cultivars [
86]. Phenolic compounds identified in strawberry fruits containing ellagic acid were reported to have a high antioxidant capacity [
87].
3.5. Inoculation and Infection Process
Firstly, after 18 days from the storage period of the treated fruits with the natural oils, it was observed a fungal growth over untreated fruits, and treated fruits with
E. camaldulensis var obtusa EO and
M. oleifera FO (
Figure 3).
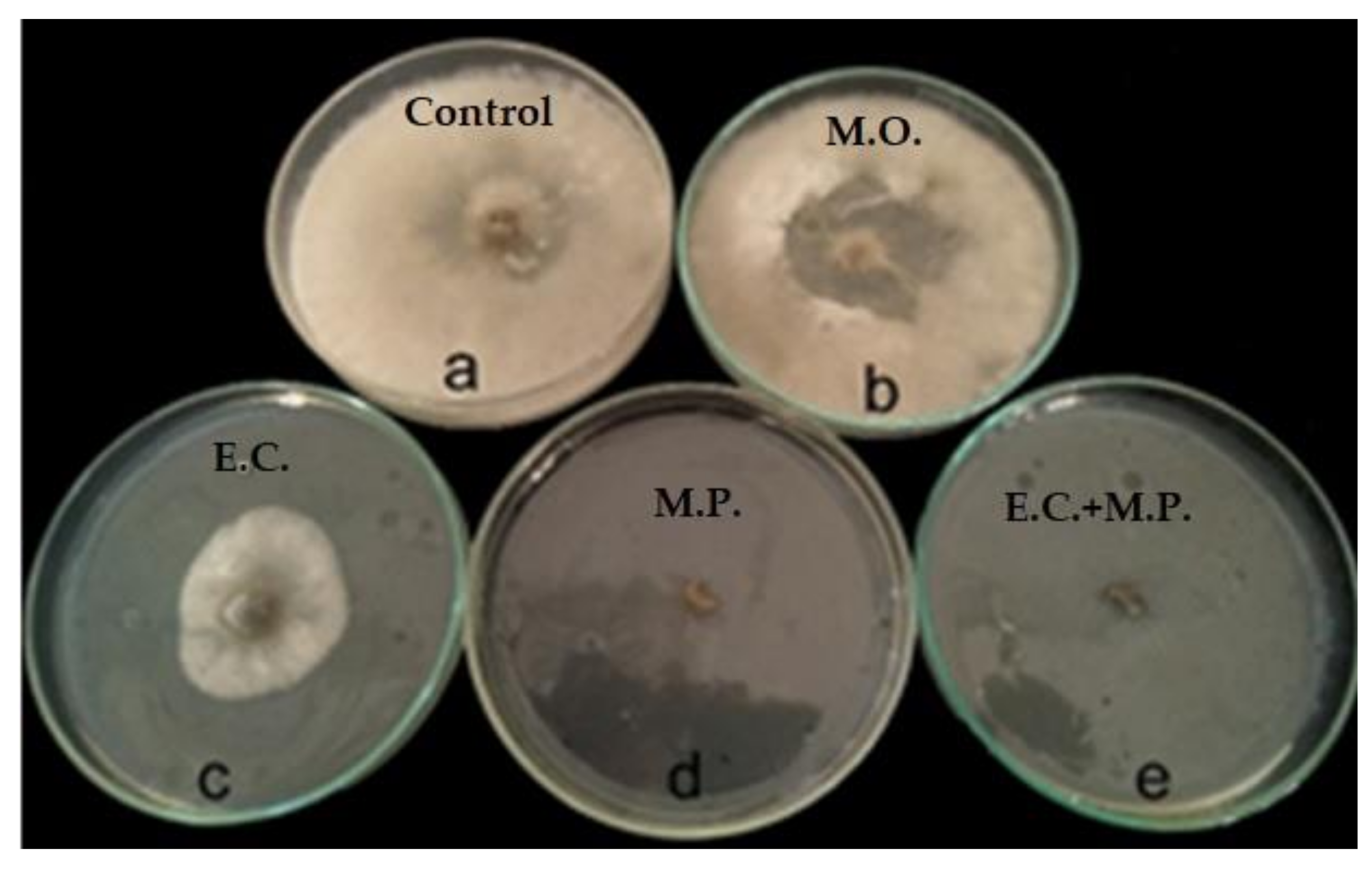
Therefore, we conducted the primary in vitro experiment of the antifungal activity of the tested oils examined at 0.05% concentration as shown in
Table 5 and
Figure 4. All tested oils had different degrees of antifungal activity against
B. cinerea in terms of the radial colony growth (cm) as measured at 2, 4 and 7 days from incubation. No radial growth from the
M. piperita and the
E. camaldulensis var obtusa and
M. piperita (1/1
v/
v) EOs in Petri dishes were noted throughout the experimental period.
The highest antifungal activity was recorded for
M. piperita EO and
E. camaldulensis var obtusa and
M. piperita mix EOs (1/1
v/
v) with an antifungal index of 100%, while
E. camaldulensis var obtusa EO showed a low to moderate effect on the radial growth of
B. cinerea with antifungal indices of 76.67, 63.7 and 31.1% at 2, 4 and 7 DAI, respectively (
Table 6). On the other hand, moringa FO demonstrated that reduction in mycelial growth of
B. cinerea was very weak and reached 4.97 and 13.42% at 2 and 4 DAI, with no difference being observed in radial growth from the non-amended oil control at day 7 (
Table 6). The application of EOs is a very attractive and eco-friendly method to control post-harvest diseases.
The infection rate (%) of strawberry fruits caused by
B. cinerea is shown in
Table 7, where it increased from 7 to 10 days after the infection, but the lowest value was observed in fruits treated with
M. piperita EO and the mix EOs after 10 days with percentages of 18% and 20%, respectively, compared to the control (87%). The lowest lesion diameters (cm) were also found in the fruits treated with
M. piperita EO and the mix EOs (
Table 8). The mycelial growth inhibition of
B. cinerea (%) in the fruits treated with
M. piperita EO and the mix EOs was 38.78 and 46.94 %, respectively (
Table 9).
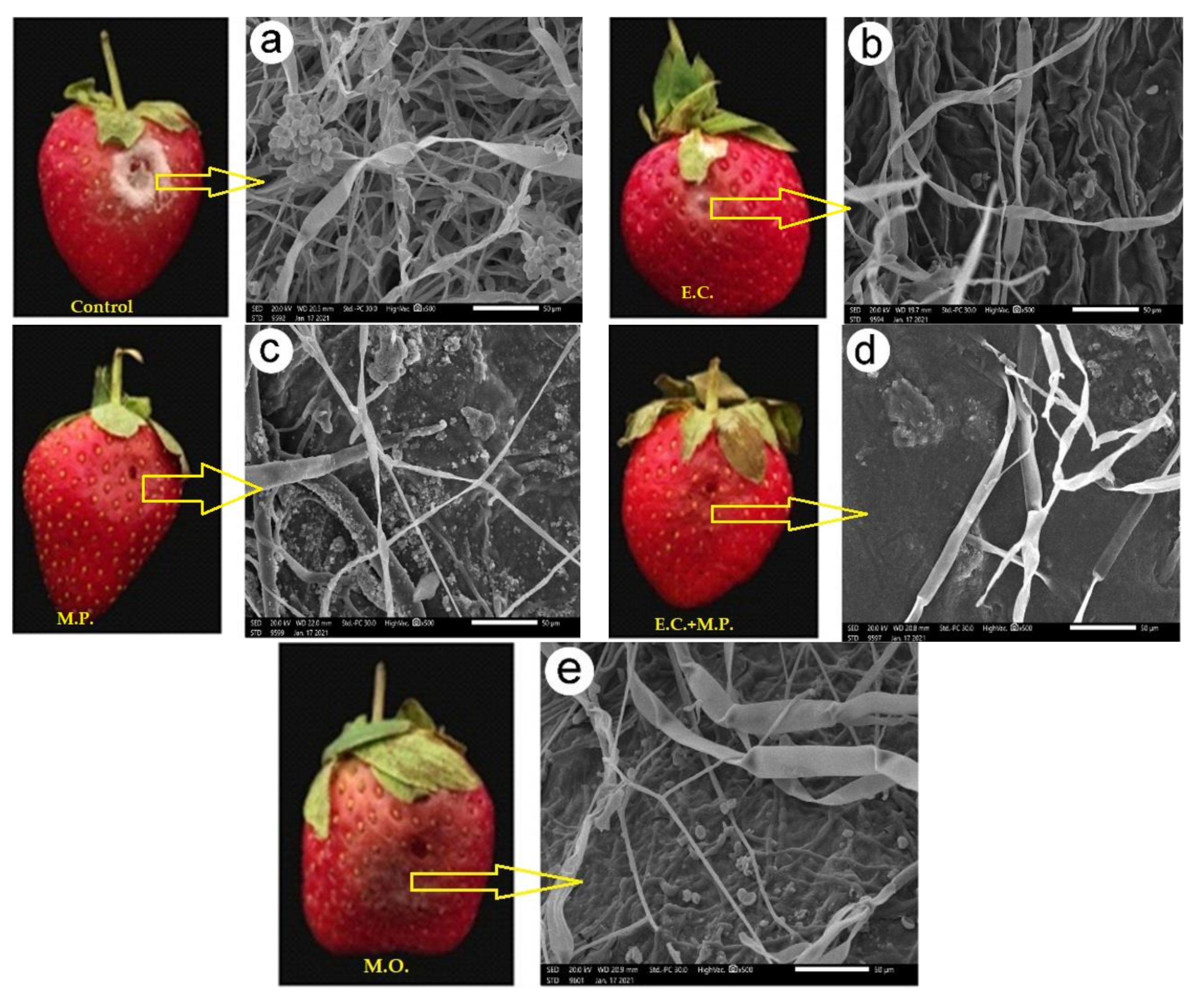
The visual observation of the fruits inoculated with
B. cinerea 10 days after inoculation is shown in
Figure 5. The mycelial morphology of
B. cinerea grown on strawberry fruit treated with the studied oils is observed by SEM at 10 days (
Figure 5). There were regular, uniform and complete mycelia with smooth surfaces, relatively strong and with high spore production in the control (
Figure 5a). The fungal mycelial growth of
B. cinerea was decreased after 10 days in the fruits treated with
E. camaldulensis var obtusa EO (
Figure 5b), but fungus hyphae are shown in dense growth. Fruit surfaces treated with
M. piperita EO (
Figure 5c) and the combined
E. camaldulensis var obtusa +
M. piperita EOs (
Figure 5d) showed great morphological changes, including irregular growth of the mycelium, formation of verrucous surface, shrinkage, collapse and hollowing of hyphae. The morphology of mycelium
B. cinerea grown on strawberry fruit surfaces treated with moringa FO (
Figure 5e) was abnormal growth, lysis, shrinkage, reduced hyphal length and diameters with lower production of conidia compared to the control treatment. Recently, our group and various publications have documented the antifungal activity of EOs and plant extracts including rosemary, peppermint, bay, basil, tea tree, celery seed fennel and
Cinnamomum camphora [
2,
16,
41,
88]. The in vitro result showed that the EOs could be candidates for a natural antifungal in food preservation technology.
The beneficial effect of EO from
M. peperita against
B. cinerea was achieved after 15 days of storage by immersing cherry tomato fruits into this oil [
89]. Medicinal and aromatic plants (MAPs) are very important in various fields, such as the pharmaceutical, perfumery and cosmetic industries [
90]. In the last few years, the food industry has primarily used EOs as flavorings, and they represent an interesting source of natural antimicrobials for food preservation [
91]. Carvacrol, cymene and
γ-terpinene, the main chemical compounds from several EOs, showed potential antifungal activity against selected fungi including
B. cinerea [
92,
93]. Their findings are in agreement with the results of this study, where GC–MS analysis showed that an EO mixture of
Eucalyptus and
M. peperita (1/1
v/
v) exhibited the strongest antifungal activity.
The antimicrobial activity of EOs has been investigated against a large number of fungi [
94,
95].
E. citriodora, EO was found to exhibit
B. cinerea fungitoxicity on grapes, with 100% growth inhibition [
96]. Thyme and lemongrass have revealed great potential in post-harvest disease control [
60].



 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}