
Influence of Harvesting Time on Phenolic and Mineral Profiles and Their Association with the Antioxidant and Cytotoxic Effects of Atractylodes japonica Koidz




 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cultivation of A. japonica
Sample Collection and Preparation of Plant Extracts
2.3. Determination of Total Phenolic Contents (TPC)
2.4. Determination of Total Flavonoid Contents (TFC)
2.5. Quantification of Phenolic Compound Contents by Liquid Chromatography–Mass Spectrometry/Mass Spectrometry (LC–MS/MS)
2.6. Antioxidant Activity
2.6.1. 1,1-Diphenyl–2-Picrylhydrazyl (DPPH) Radical Scavenging Assay
2.6.2. 2,2′-Azino-bis-3-Ethylbenzthiazoline-6-Sulphonic Acid (ABTS+) Assay
2.7. Sample Digestion
2.8. Instrumentation and Quantification of Minerals
2.9. Assessment of Toxicity on Fibroblasts
2.10. Assessment of Irritation Potential of A. japonica Root Extracts in Fertile Chicken Egg
2.11. Statistical Analysis
3. Results
3.1. Total Phenolic and Flavonoid Contents
3.2. Phenolic Compound Contents
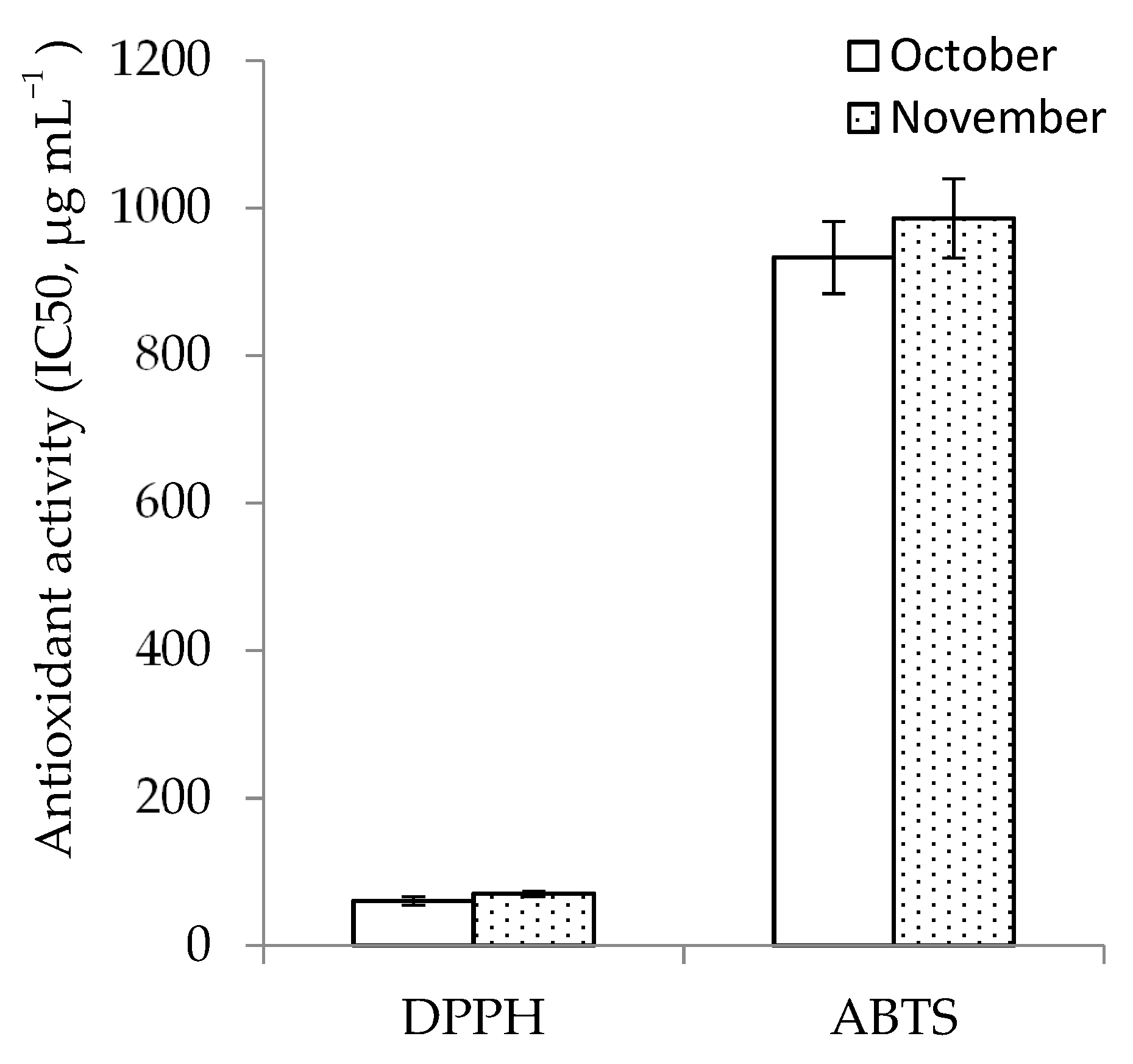
3.3. Antioxidant Activity
3.4. Mineral Content
3.5. Cytotoxicity Activity
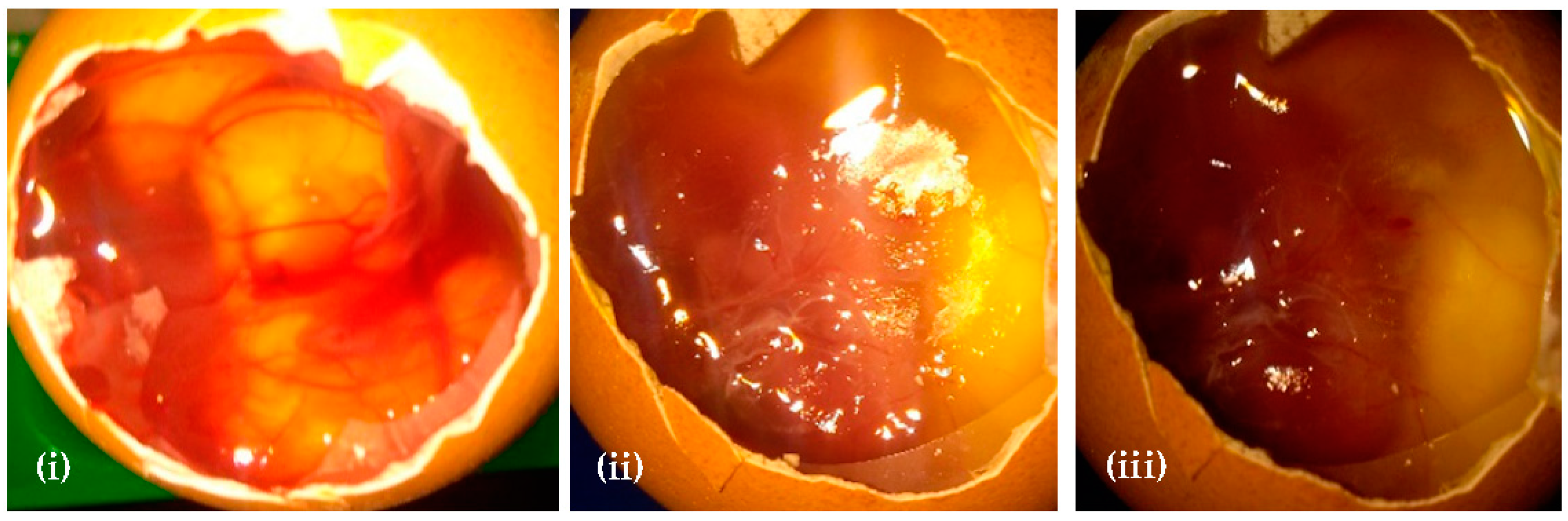
3.6. Assessment of Irritation Potential of Root extracts by the HET–CAM Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, C.M.; Shin, M.K.; Ahn, D.G.; Lee, K.S. Chungyak Daesajun; Jungdam Publisher: Seoul, Korea, 1997; Volume 8, pp. 3969–3976. [Google Scholar]
- Kwak, J.S.; Seong, H.G.; Chang, K.J. Medicinal Plant Cultivation; Blue Happy Press: Seoul, Korea, 2011; pp. 1–423. [Google Scholar]
- Park, J.H.; Seong, S.H. Medicinal Plant; Shinilbooks: Seoul, Korea, 2007; pp. 1–568. [Google Scholar]
- Satoh, K.; Nagai, F.; Ushiyama, K.; Kano, I. Specific inhibition of Na+, K(+)-ATPase activity by atractylon, a major component of byaku-jutsu, by interaction with enzyme in the E2 state. Biochem. Pharmacol. 1996, 51, 339–343. [Google Scholar] [CrossRef]
- Jang, M.H.; Shin, M.C.; Kim, Y.J.; Kim, C.J.; Kim, Y.H.; Kim, E.H. Atractylodes japonica suppresses lipopolysaccharide-stimulated expression of inducible nitric oxide synthase and cyclooxygenase-2 in RAW 264.7 macrophages. Biol. Pharm. Bull. 2004, 27, 324–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, M.H.; Kim, J.-H.; Bae, H.; Lee, N.-Y.; Shin, Y.-C.; Kim, S.-H.; Ko, S.-G. Atractylodes japonica Koidzumi inhibits the production of pro-inflammatory cytokines through inhibition of the NF-κB/IκB signal pathway in HMC-1 human mast cells. Arch. Pharm. Res. 2020, 33, 843–851. [Google Scholar] [CrossRef]
- Jeong, S.I.; Kim, S.Y.; Kim, S.J.; Hwang, B.S.; Kwon, T.H.; Yu, K.Y.; Hang, S.H.; Suzuki, K.; Kim, K.J. Antibacterial activity of phytochemicals isolated from Atractylodes japonica against methicillin-resistant Staphylococcus aureus. Molecules 2010, 15, 7395–7402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, B.S.; Kim, Y.H.; Tomiyama, M.; Nakamura, N.; Miyashiro, H.; Otake, T.; Hattori, M. Inhibitory effects of Korean plants on HIV-1 activities. Phytother. Res. 2001, 15, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Sugawara, H.; Saito, K.; Kano, Y. Effects of the acetylene compound from Atractylodes rhizome on experimental gastric ulcers induced by active oxygen species. Biol. Pharm. Bull. 1994, 17, 1364–1368. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.H.; Jeong, S.I.; Lee, J.H.; Hwang, B.S.; Kim, S.J.; Lee, S.; Choi, B.K.; Jung, K.Y. Pharmacological mechanism responsible for the Atractylodes japonica-induced distal colonic contraction in rats. Phytomedicine 2011, 18, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.W.; Kiyohara, H.; Matsumoto, T.; Yang, H.C.; Yamada, H. Intestinal immune system modulating polysaccharides from rhizomes of Atractylodes lancea. Planta Med. 1998, 64, 714–719. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.G.; Jan, Y.S.; Tsai, P.W.; Norimoto, H.; Michihara, S.; Murayama, C.; Wang, C.C. Anti-inflammatory and antinociceptive constituents of Atractylodes japonica koidzumi. J. Agric. Food Chem. 2016, 64, 2254–2262. [Google Scholar] [CrossRef]
- Kitajima, J.; Kamoshita, A.; Ishikawa, T.; Takano, A.; Fukuda, T.; Isoda, S.; Ida, Y. Glycosides of Atractylodes japonica. Chem. Pharm. Bull. 2003, 51, 152–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.C.; Lin, S.Y.; Cheng, H.C.; Hou, W.C. Pro-oxidant and cytotoxic activities of atractylenolide I in human promyeloleukemic HL-60 cells. Food Chem. Toxicol. 2006, 44, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.H.; Jeong, S.I.; Lee, J.H.; Hwang, B.S.; Lee, S.; Choi, B.K.; Jung, K.Y. Acetylene compound isolated from Atractylodes japonica stimulates the contractility of rat distal colon via inhibiting the nitrergic-purinergic relaxation. J. Ethnopharmacol. 2011, 134, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Karakaya, S. Bioavailability of phenolic compounds. Critical Rev. Food Sci. Nutr. 2004, 44, 453–464. [Google Scholar] [CrossRef]
- Ninfali, P.; Mea, G.; Giorgini, S.; Rocchi, M.; Bacchiocca, M. Antioxidant capacity of vegetables, spices and dressings relevant to nutrition. Br. J. Nutr. 2005, 93, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haslam, E. Natural polyphenols (vegetable tannins) as drugs: Possible modes of action. J. Nat. Prod. 1996, 59, 205. [Google Scholar] [CrossRef]
- Soares, S.E. Phenolic acids as antioxidants. Rev. Nutr. 2002, 15, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Szymczycha-Madeja, A.; Welna, M.; Pohl, P. Elemental analysis of teas and their infusions by spectrometric methods. Trends Anal. Chem. 2012, 35, 165–181. [Google Scholar] [CrossRef]
- Valmorbida, J.; Boaro, C.S.F.; Scavroni, J.; David, E.F.S. Crescimento de Mentha piperita L. cultivada em solução nutritiva com diferentes doses de potássio. Revista Brasileira de Plantas Medicinais 2007, 9, 27–31. [Google Scholar]
- Treutter, D. Biosynthesis of phenolic compounds and its regulation in apple. Plant Growth Regul. 2001, 34, 71–89. [Google Scholar] [CrossRef]
- Dong, T.T.; Cui, X.M.; Song, Z.H.; Zhao, K.J.; Ji, Z.N.; Lo, C.K.; Tsim, K.W. Chemical assessment of roots of Panax notoginseng in China: Regional and seasonal variations in its active constituents. J. Agric. Food Chem. 2003, 51, 4617–4623. [Google Scholar] [CrossRef]
- Pęksa, A.; Golubowska, G.; Rytel, E.; Lisińska, G.; Aniolowski, K. Influence of harvest date on glycoalkaloid content of three potato varieties. Food Chem. 2002, 87, 313–317. [Google Scholar] [CrossRef]
- Lemos, M.F.; Lemos, M.F.; Pacheco, H.P.; Guimarães, A.C.; Fronza, M.; Endringer, D.C.; Scherer, R. Seasonal variation affects the composition and antibacterial and antioxidant activities of Thymus vulgaris. Ind. Crops Prod. 2017, 95, 543–548. [Google Scholar] [CrossRef]
- Sartor, T.; Xavier, V.B.; Falcão, M.A.; Mondin, C.A.; Santos, M.A.; Cassel, E.; Santarém, E.R. Seasonal changes in phenolic compounds and in the biological activities of Baccharis dentata (Vell.) GM Barroso. Ind. Crops Prod. 2013, 51, 355–359. [Google Scholar] [CrossRef]
- Yao, X.H.; Zhang, Z.B.; Song, P.; Hao, J.Y.; Zhang, D.Y.; Zhang, Y.F. Different harvest seasons modify bioactive compounds and antioxidant activities of Pyrola incarnata. Ind. Crops Prod. 2016, 94, 405–412. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Moreno, M.I.N.; Isla, M.I.; Sampietro, A.R.; Vattuone, M.A. Comparison of the free radical-scavenging activity of propolis from several regions of Argentina. J. Ethnopharmacol. 2000, 71, 109–114. [Google Scholar] [CrossRef]
- Chung, I.M.; Chelliah, R.; Oh, D.H.; Kim, S.H.; Yu, C.Y.; Ghimire, B.K. Tupistra nutans wall. root extract, rich in phenolics, inhibits microbial growth and α-glucosidase activity, while demonstrating strong antioxidant potential. Braz. J. Bot. 2019, 42, 383–397. [Google Scholar] [CrossRef]
- Xing, Q.; Kadota, S.; Tadata, T.; Namba, T. Antioxidative effect of phenylethanoids from Cistanche deserticola. Biol. Pharm. Bull. 1996, 19, 1580–1585. [Google Scholar] [CrossRef] [Green Version]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Hawkins Byrne, D. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Sandra, F.; Sudiono, J.; Trisfilha, P.; Pratiwi, D. Cytotoxicity of Alpinia galanga rhizome crude extract on NIH-3T3 cells. Indones. Biomed. J. 2017, 9, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Routray, W.; Orsat, V. Variation of phenolic profile and antioxidant activity of North American highbush blueberry leaves with variation of time of harvest and cultivar. Ind. Crop Prod. 2014, 62, 147–155. [Google Scholar] [CrossRef]
- Salawu, S.O.; Bester, M.J.; Duodu, K.G. Phenolic composition and bioactive properties of cell wall preparations and whole grains of selected cereals and legumes. J. Food Biochem. 2014, 38, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Remorini, D.; Tavarini, S.; Degl, E.; Loreti, F.; Massai, R.; Guidi, L. Effect of rootstocks and harvesting time on the nutritional quality of peel and flesh of peach fruits. Food Chem. 2008, 110, 361–367. [Google Scholar] [CrossRef]
- Jaakola, L.; Määttä, K.; Pirttilä, A.M.; Torronen, R.; Karenlampi, S.; Hohtola, A. Expression of genes involved in anthocyanin biosynthesis in relation to anthocyanin, proanthocyanidin, and flavonol levels during bilberry fruit development. Plant Phys. 2002, 130, 729–739. [Google Scholar] [CrossRef] [Green Version]
- Francini, A.; Sebastiani, L. Phenolic compounds in apple (Malus domestica Borkh.): Compounds characterization and stability during postharvest and after processing. Antioxidants 2013, 2, 181–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krüger, E.; Dietrich, H.; Hey, M.; Patz, C.D. Effects of cultivar, yield, berry weight, temperature and ripening stage on bioactive compounds of black currants. J. App. Bot. Food Qual. 2011, 84, 40–46. [Google Scholar]
- Martz, F.; Jaakola, L.; Julkunen-Tiitto, R.; Stark, S. Phenolic composition and antioxidant capacity of bilberry (Vaccinium myrtillus) leaves in Northern Europe following foliar development and along environmental gradients. J. Chem. Ecol. 2010, 36, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Nurmi, K.; Ossipov, V.; Haukioja, E.; Pihlaja, K. Variation of total phenolic content and individual low molecular weight phenolics in foliage of mountain birch trees (Betula pubescens spp. tortuosa). J. Chem. Ecol. 1996, 22, 2023–2040. [Google Scholar] [CrossRef] [PubMed]
- Samanta, A.; Das, G.; Das, S.K. Roles of flavonoids in plants. Int. J. Pharm. Sci. Technol. 2011, 6, 12–35. [Google Scholar]
- Agati, G.; Tattini, M. Multiple functional roles of flavonoids in photoprotection. New Phytol. 2010, 186, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Tattini, M.; Galardi, C.; Pinelli, P.; Massai, R.; Remorini, D.; Agati, G. Differential accumulation of flavonoids and hydroxycinnamates in leaves of Ligustrum vulgare under excess light and drought stress. New Phytol. 2004, 163, 547–561. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.R.; Chen, Y.H.; Guo, Q.S.; Wang, W.M.; Liu, L.; Fan, J.; Cao, L.P.; Li, C. Short-term UV-B radiation effects on morphology, physiological traits and accumulation of bioactive compounds in Prunella vulgaris L. J. Plant Interact. 2017, 12, 348–354. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.; Giannini, A.; Lercari, B. Photomorphogenic responses to UV radiation III: A comparative study of UVB effects on anthocyanin and flavonoid accumulation in wild type and aurea mutant of tomato (Lycopersicon esculentum Mill.). Photochem. Photobiol. 1995, 62, 1081–1087. [Google Scholar] [CrossRef]
- Wu, S.; Li, J.; Wang, Q.; Cao, H.; Cao, J.; Xiao, J. Seasonal dynamics of the phytochemical constituents and bioactivities of extracts from Stenoloma chusanum (L.) Ching. Food Chem. Toxicol. 2017, 108, 458–466. [Google Scholar] [CrossRef]
- Armaleo, D.; Zhang, Y.; Cheung, S. Light might regulate divergently depside and depsidone accumulation in the lichen Parmotrema hypotropum by affecting thallus temperature and water potential. Mycologia 2008, 100, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Zeni, A.L.B.; Albuquerque, C.A.C.D.; Gonçalves, F.; Latini, A.; Tasca, C.I.; Podestá, R.; Pagliosa, C.M.; Duarte, F.S.; Lima, T.; Maraschin, M. Phytochemical profile, toxicity and antioxidant activity of Aloysia gratissima (Verbenaceae). Quimica Nova 2013, 36, 69–73. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, S.; Bhanger, M.I. Effect of season and production location on antioxidant activity of Moringa oleifera leaves grown in Pakistan. J. Food Compos. Anal. 2006, 19, 544–551. [Google Scholar] [CrossRef]
- Pirvu, L.; Grigore, A.; Bubueanu, C.; Draghici, E. Comparative analytical and antioxidant activity studies on a series of Fagus sylvatica L. leaves extracts. J. Planar Chromatogr. Mod. TLC 2013, 26, 237–242. [Google Scholar] [CrossRef]
- Sati, P.; Pandey, A.; Rawat, S.; Rani, A. Phytochemicals and antioxidants in leaf extracts of Ginkgo biloba with reference to location, seasonal variation and solvent system. J. Pharm. Res. 2013, 7, 804–809. [Google Scholar] [CrossRef]
- Oboh, G.; Rocha, J.B.T. Polyphenols in red pepper [Capsicum annuum var. aviculare (Tepin)] and their protective effect on some pro-oxidants induced lipid peroxidation in brain and liver. Eur. Food Res. Technol. 2006, 225, 239–247. [Google Scholar] [CrossRef]
- Nishimura, F.C.; de Almeida, A.C.; Ratti, B.A.; Ueda-Nakamura, T.; Nakamura, C.V.; Ximenes, V.F.; Silva, S.O. Antioxidant effects of quercetin and naringenin are associated with impaired neutrophil microbicidal activity. Evid. Based Complement. Alternat. Med. 2013, 2013, 795916. [Google Scholar] [CrossRef]
- Abidille, M.D.H.; Singh, R.P.; Jayaprakashna, G.K.; Jena, B.S. Antioxidant activity of the extracts from Dillenia indica fruits. Food Chem. 2005, 90, 891–896. [Google Scholar] [CrossRef]
- Kur, C.; Kapoor, H.C. Anti-oxidant activity and total phenolic content of some Asian vegetables. Int. J. Food Sci. Technol. 2002, 37, 153–161. [Google Scholar] [CrossRef]
- Melo, E.A.; Lima, V.L.A.; Maciel, M.I.S.; Caetano, A.C.S.; Leal, F.L.L. Polyphenol, ascorbic acid and total carotenoid contents in common fruits and vegetables. Braz. J. Food Technol. 2006, 9, 89–94. [Google Scholar]
- Sato, Y.; Itagaki, S.; Kurokawa, T.; Ogura, J.; Kobayashi, M.; Hirano, T.; Sugawara, M.; Iseki, K. In vitro and in vivo antioxidant properties of chlorogenic acid and caffeic acid. Int. J. Pharm. 2011, 403, 136–138. [Google Scholar] [CrossRef]
- Hidalgo, M.; Sánchez-Moreno, C.; Pascual-Teresa, S. Flavonoid–flavonoid interaction and its effect on their antioxidant activity. Food Chem. 2010, 121, 691–696. [Google Scholar] [CrossRef]
- Close, D.C.; McArthur, C. Rethinking the role of many plant phenolics—Protection from photodamage, not herbivores? Oikos 2002, 99, 166–172. [Google Scholar] [CrossRef]
- Gobbo-neto, L.; Lopes, N.P. Plantas medicinais: Fatores de influência no conteúdo de metabólitos secundários. Quimica Nova 2007, 30, 374–381. [Google Scholar] [CrossRef]
- Gomes, A.F.; Almeida, M.P.; Leite, M.F.; Schwaiger, S.; Stuppner, H.; Halabalaki, M.; Amaral, J.G.; David, J.M. Seasonal variation in the chemical composition of two chemotypes of Lippia alba. Food Chem. 2019, 273, 186–193. [Google Scholar] [CrossRef]
- Askin, M.A.; Balta, M.F.; Tekintas, F.E.; Kazankaya, A.; Balta, F. Fatty acid composition affected by kernel weight in almond [Prunus dulcis (Mill.) D.A. Webb] genetic resources. J. Food Compos. Anal. 2007, 20, 7–12. [Google Scholar] [CrossRef]
- Rout, G.R.; Sahoo, S. Role of iron in plant growth and metabolism. Rev. Agric. Sci. 2015, 3, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Marschner, P. Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; Volume 651. [Google Scholar]
- Camargo, P.N.; Silva, O. Manual de Adubação Foliar; Herba: Sao Paulo, Brazil, 1975; Volume 258. [Google Scholar]
- Epstein, E. Nutrição Mineral das Plantas: Princípios e Perspectivas; EDUSP: Sao Paulo, Brazil, 1975; Volume 344. [Google Scholar]
- Stateras, D.C.; Moustakas, N.K. Seasonal changes of macro- and micro-nutrients concentration in olive leaves. J. Plant Nutr. 2018, 4, 186–196. [Google Scholar] [CrossRef]
- Baloyi1, B.M.; Ayodele, V.I.; Addo-Bediako, A. Effects of leaf harvest on crude protein and mineral contents of selected early maturing lines of lablab (Lablab purpureus). Afr. J. Agric. Res. 2013, 8, 449–453. [Google Scholar]
- Christian, D.G.; Yates, N.E.; Riche, A.B. The effect of harvest date on the yield and mineral content of Phalaris arundinacea L. (reed canary grass) genotypes screened for their potential as energy crops in southern England. J. Sci Food Agric. 2006, 86, 1181–1188. [Google Scholar] [CrossRef]
- Yanai, N.; Shiotani, S.; Hagiwara, S.; Nabetani, H.; Nakajima, M. Antioxidant combination inhibits reactive oxygen species mediated damage. Biosci. Biotechnol. Biochem. 2008, 72, 3100–3106. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Li, S.; Li, H.B.; Deng, G.F.; Ling, W.H.; Xu, X.R. Antiproliferative activities of tea and herbal infusions. Food Funct. 2013, 4, 530–538. [Google Scholar] [CrossRef]
- Estevinho, L.; Pereira, A.P.; Moreira, L.; Dias, L.G.; Pereira, E. Antioxidant and antimicrobial effects of phenolic compounds extracts of northeast Portugal honey. Food Chem. Toxicol. 2008, 46, 3774–3779. [Google Scholar] [CrossRef]
- Aleksic, V.; Knezevic, P. Antimicrobial and antioxidative activity of extracts and essential oils of Myrtus communis L. Microbiol. Res. 2014, 169, 240–254. [Google Scholar] [CrossRef]
- Llorent-Martínez, E.J.; Ortega-Barrales, P.; Zengin, G.; Mocan, A.; Simirgiotis, M.J.; Ceylan, R.; Uysal, S.; Aktumsek, A. Evaluation of antioxidant potential, enzyme inhibition activity and phenolic profile of Lathyrus cicera and Lathyrus digitatus: Potential sources of bioactive compounds for the food industry. Food Chem. Toxicol. 2017, 107, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Palchaudhuri, R.; Hergenrother, P.J. DNA as a target for anticancer compounds: Methods to determine the mode of binding and the mechanism of action. Curr. Opin. Biotechnol. 2007, 18, 497–503. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Ghimire, B.K.; Kim, S.H.; Yu, C.Y.; Oh, D.H.; Chelliah, R.; Kwon, C.; Kim, Y.J.; Chung, I.M. Assessment of phenolic and mineral profiles and their association with the antimicrobial potential, antioxidant, and cytotoxic effect of Lycium chinense Miller. Plants 2020, 9, 1023. [Google Scholar] [CrossRef] [PubMed]
- Araújo, K.C.F.; Costa, E.M.D.M.; Pazini, F.; Valadares, M.C.; De Oliveira, V. Bioconversion of quercetin and rutin and the cytotoxicity activities of the transformed products. Food Chem. Toxicol. 2013, 51, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Bonechi, C.; Donati, A.; Tamasi, G.; Leone, G.; Consumi, M.; Rossi, C.; Lamponi, S.; Magnani, A. Protective effect of quercetin and rutin encapsulated liposomes on induced oxidative stress. Biophys. Chem. 2018, 233, 55–63. [Google Scholar] [CrossRef]
- Tamasi, G.; Baratto, M.C.; Bonechi, C.; Byelyakova, A.; Pardini, A.; Donati, A.; Leone, G.; Consumi, M.; Lamponi, S.; Magnani, A.; et al. Chemical characterization and antioxidant properties of products and by-products from Olea europaea L. Food Sci. Nutr. 2019, 7, 2907–2920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christian, L.; Freia, F.S.; Angela, R.; Szymon, K.; Daniel, K.; Bart, D.W.; Heike, W.; Florian, K.G. Alternative methods for the replacement of eye irritation testing. ALTEX 2016, 33, 55–67. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | RT | Q1 (m/z) 1 | Q3-1(m/z) 2 | DP (V) 3 | EP (V) 4 | CEP (V) 5 | CE (eV) 6 | CXP (V) 7 | 8 Phenolic Compounds µgg−1 Dry Weight of Root Extract.) | |
|---|---|---|---|---|---|---|---|---|---|---|
| October November | ||||||||||
| Protocatechuic acid | 9.61 | 152.89 | 108.90 | −31.00 | −9.00 | −8.00 | −18.00 | −22.00 | 1166.70 ± 20.90 g | 636.54 ± 87.72 g |
| p-Hydroxybenzoic acid | 11.71 | 136.88 | 92.90 | −16.00 | −9.50 | −14.00 | −18.00 | −18.00 | LOD | LOD |
| Chlorogenic acid | 9.95 | 352.84 | 191.00 | −16.00 | −9.50 | −18.00 | −18.00 | −36.00 | 133833.30 ± 50.00 i | 5298.23 ± 38.82i |
| p-Coumaric acid | 13.95 | 162.86 | 118.90 | −16.00 | −5.00 | −8.00 | −20.00 | −22.00 | 36.80 ± 2.00 a | 170.33 ± 26.52 b |
| Ferulic acid | 14.27 | 192.88 | 133.90 | −6.000 | −6.50 | −12.00 | −22.00 | −26.00 | LOQ | LOQ |
| Rutin | 11.90 | 609.00 | 299.70 | −96.00 | −8.00 | −26.00 | −48.00 | −52.00 | 639.20 ± 10.00 e | 564.57 ± 13.30 e |
| Quercetin | 18.16 | 300.88 | 150.80 | −31.00 | −10.50 | −18.00 | −28.00 | −32.00 | LOD | LOQ |
| 5−Sulfosalicylic acid | 6.68 | 216.77 | 198.80 | −21.00 | −7.50 | −10.00 | −18.00 | −44.00 | LOQ | LOQ |
| Homogentisic acid | 8.62 | 166.92 | 122.90 | −21.00 | −7.00 | −14.00 | −14.00 | −28.00 | LOD | LOD |
| Salicylic acid | 18.66 | 136.87 | 92.900 | −16.00 | −6.00 | −12.00 | −20.00 | −20.00 | 536.70 ± 12.00 b | 354.40 ± 5.00 f |
| Caffeic acid | 10.85 | 178.86 | 134.80 | −11.00 | −10.00 | −12.00 | −20.00 | −30.00 | 583.66 ± 15.00 c | 146.30 ± 7.00 c |
| Orientin | 10.10 | 447.09 | 327.00 | −61.00 | −11.00 | −20.00 | −22.00 | −54.00 | 591.17 ± 17.00 cd | 309.69 ± 21.63d |
| Vitexin | 14.60 | 430.82 | 310.80 | −56.00 | −9.00 | −28.00 | −22.00 | −50.00 | 592.00 ± 11.90 d | 517.66 ± 1.30 f |
| Apigenin | 21.00 | 268.94 | 117.00 | −56.00 | −8.50 | −16.00 | −48.00 | −22.00 | 1325.00 ± 20.00 h | 67.30 ± 3.00 a |
| Luteolin | 17.66 | 284.96 | 133.20 | −66.00 | −10.00 | −16.00 | −46.00 | −30.00 | 869.30 ± 10.00 f | 946.63 ± 12.00 h |
| L-Phenylalanine | 7.00 | 163.88 | 146.80 | −51.00 | −10.00 | −12.61 | −16.00 | −30.00 | LOD | LOD |
| Assay | TPC | TFC | Protocatechuic Acid | ChlorogenicAcid | p-Coumaric Acid | Rutin | Salicylic Acid | Caffeic Acid | Orientin | Vitexin | Apigenin | Luteolin |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DPPH (October) | 0.992 * | 0.961 * | 0.500 * | 0.655 * | 0.550 ** | 0.590 * | 0.500 * | 0.817 * | 0.810 ** | 0.693 | 0.721 | 0.655 |
| ABTS (October) | 0.997 * | 0.997 * | 0.663 ** | 0.792 * | 0.663 * | 0.663 ** | 0.663 ** | 0.916 ** | 0.910 * | 0.823 | 0.845 | 0.792 |
| DPPH (November) | 0.999 * | 0.993 * | 0.311 | 0.717 * | 0.632 * | 0.767 * | 0.632 ** | 0.539 * | 0.622 * | 0.539 | 0.767 | 0.632 |
| ABTS (November) | 0.965 | 0.911 * | 0.013 | 0.475 | 0.371 | 0.540 * | 0.371 | 0.262 | 0.614 | 0.262 | 0.540 | 0.371 |
| Element | Concentration (mg L−1) a | Linearity (r2) | Slope (S) | Response SD | SD c (%) | LOD b | LOQ b | |
|---|---|---|---|---|---|---|---|---|
| μg mL−1 | ||||||||
| Macro elements | Ca | 0.5–10 | 0.999576 | 8157 | 1103.17 | 1.15 | 0.405726 | 1.352421 |
| Mg | 0.5–10 | 0.999738 | 19,920 | 2106.85 | 0.94 | 0.317297 | 1.057656 | |
| Micro elements | Al | 0.5–10 | 0.999932 | 3730 | 358.88 | 0.96 | 0.288643 | 0.962145 |
| Ba | 0.5–10 | 0.999825 | 5071 | 505.09 | 0.99 | 0.298811 | 0.996036 | |
| Cd | 0.5–10 | 0.999812 | 2007 | 156.53 | 0.78 | 0.233976 | 0.779920 | |
| Cu | 0.5–10 | 0.999852 | 19,530 | 1778.00 | 0.91 | 0.273118 | 0.910394 | |
| Fe | 0.5–10 | 0.999803 | 4031 | 352.03 | 0.87 | 0.261992 | 0.873307 | |
| Mn | 0.5–10 | 0.999901 | 31,120 | 3071.07 | 0.98 | 0.296054 | 0.986848 | |
| Zn | 0.5–10 | 0.999799 | 5112 | 378.98 | 0.74 | 0.222406 | 0.741354 | |
| Trace elements | As | 0.5–10 | 0.999876 | 47.10 | 3.54 | 0.75 | 0.225478 | 0.751592 |
| Bi | 0.5–10 | 0.999907 | 286.5 | 38.39 | 1.34 | 0.401990 | 1.339965 | |
| Co | 0.5–10 | 0.999131 | 1197 | 111.51 | 0.93 | 0.279474 | 0.931579 | |
| Cr | 0.5–10 | 0.999841 | 4921 | 522.49 | 1.06 | 0.318527 | 1.061756 | |
| Li | 0.5–10 | 0.999783 | 32,120 | 3995.40 | 1.24 | 0.373169 | 1.243898 | |
| Ni | 0.5–10 | 0.999757 | 1951 | 174.43 | 0.89 | 0.268216 | 0.894054 | |
| Pb | 0.5–10 | 0.999792 | 277.0 | 37.92 | 1.37 | 0.410686 | 1.368953 | |
| Se | 0.5–10 | 0.999913 | 72.12 | 8.72 | 1.21 | 0.362729 | 1.209096 | |
| Ag | 0.5–10 | 0.999942 | 20,240 | 1753.28 | 0.86 | 0.259874 | 0.866245 | |
| Ti | 0.5–10 | 0.999927 | 38,130 | 2907.65 | 0.76 | 0.228769 | 0.762562 | |
| Elements | Concentration (μg mL−1) | ||
|---|---|---|---|
| October | November | ||
| Macro elements | Ca | 49.34 ± 0.838 | 92.770 ± 0.654 |
| Mg | 19.66 ± 0.2750 | 28.680 ± 0.155 | |
| Micro elements | Al | 0.701 ± 0.0073 | 0.551 ± 0.027 |
| Ba | 0.120 ± 0.0015 | 0.269 ± 0.0035 | |
| Cd | 0.089 ± 0.0013 | 0.090 ± 0.0019 | |
| Cu | 0.057 ± 0.0004 | 0.031 ± 0.0011 | |
| Fe | 0.953 ± 0.0063 | 0.875 ± 0.0115 | |
| Mn | 0.928 ± 0.0134 | 1.696 ± 0.0032 | |
| Zn | 0.218 ± 0.0018 | 0.303 ± 0.0047 | |
| Trace elements | As | 0.142 ± 0.0364 | 0.037 ± 0.0562 |
| Bi | 0.076 ± 0.0442 | 0.071 ± 0.0215 | |
| Co | 0.131 ± 0.0029 | 0.134 ± 0.0022 | |
| Cr | 0.095 ± 0.0026 | 0.095 ± 0.0010 | |
| Li | 0.081 ± 0.0007 | 0.081 ± 0.0004 | |
| Ni | 0.113 ± 0.0017 | 0.116 ± 0.0017 | |
| Pb | 0.076 ± 0.0223 | 0.087 ± 0.0193 | |
| Se | 0.143 ± 0.0443 | 0.175 ± 0.0643 | |
| Ag | 0.060 ± 0.0010 | 0.060 ± 0.0007 | |
| Ti | 0.040 ± 0.0001 | 0.040 ± 0.0003 | |
| Phenolic Compounds | Cytotoxicity |
|---|---|
| Protocatechuic acid | 0.987 ** |
| Rutin | 0.899 |
| Chlorogenic acid | 0.978 * |
| p-Coumaric acid | 0.974 * |
| Salicylic acid | 0.908 * |
| Caffeic acid | 0.959 * |
| Orientin | 0.509 |
| Vitexin | 0.350 |
| Apigenin | 0.240 |
| Luteolin | 0.150 |
| Samples | Irritation Score | Irritation Assessment |
|---|---|---|
| Negative control | 0 | Non-irritant |
| NaOH (0.1 M) | 18.00 | Strong irritant |
| Root extracts (50 mg/mL) | 0 | Non-irritant |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghimire, B.K.; Seo, J.-W.; Kim, S.-H.; Ghimire, B.; Lee, J.-G.; Yu, C.-Y.; Chung, I.-M. Influence of Harvesting Time on Phenolic and Mineral Profiles and Their Association with the Antioxidant and Cytotoxic Effects of Atractylodes japonica Koidz. Agronomy 2021, 11, 1327. https://doi.org/10.3390/agronomy11071327
Ghimire BK, Seo J-W, Kim S-H, Ghimire B, Lee J-G, Yu C-Y, Chung I-M. Influence of Harvesting Time on Phenolic and Mineral Profiles and Their Association with the Antioxidant and Cytotoxic Effects of Atractylodes japonica Koidz. Agronomy. 2021; 11(7):1327. https://doi.org/10.3390/agronomy11071327
Chicago/Turabian StyleGhimire, Bimal Kumar, Ji-Won Seo, Seung-Hyun Kim, Balkrishna Ghimire, Jae-Geun Lee, Chang-Yeon Yu, and Ill-Min Chung. 2021. "Influence of Harvesting Time on Phenolic and Mineral Profiles and Their Association with the Antioxidant and Cytotoxic Effects of Atractylodes japonica Koidz" Agronomy 11, no. 7: 1327. https://doi.org/10.3390/agronomy11071327
APA StyleGhimire, B. K., Seo, J. -W., Kim, S. -H., Ghimire, B., Lee, J. -G., Yu, C. -Y., & Chung, I. -M. (2021). Influence of Harvesting Time on Phenolic and Mineral Profiles and Their Association with the Antioxidant and Cytotoxic Effects of Atractylodes japonica Koidz. Agronomy, 11(7), 1327. https://doi.org/10.3390/agronomy11071327