
Applications and Potential of Genome-Editing Systems in Rice Improvement: Current and Future Perspectives

 ,
,  , ,
, , 
Abstract
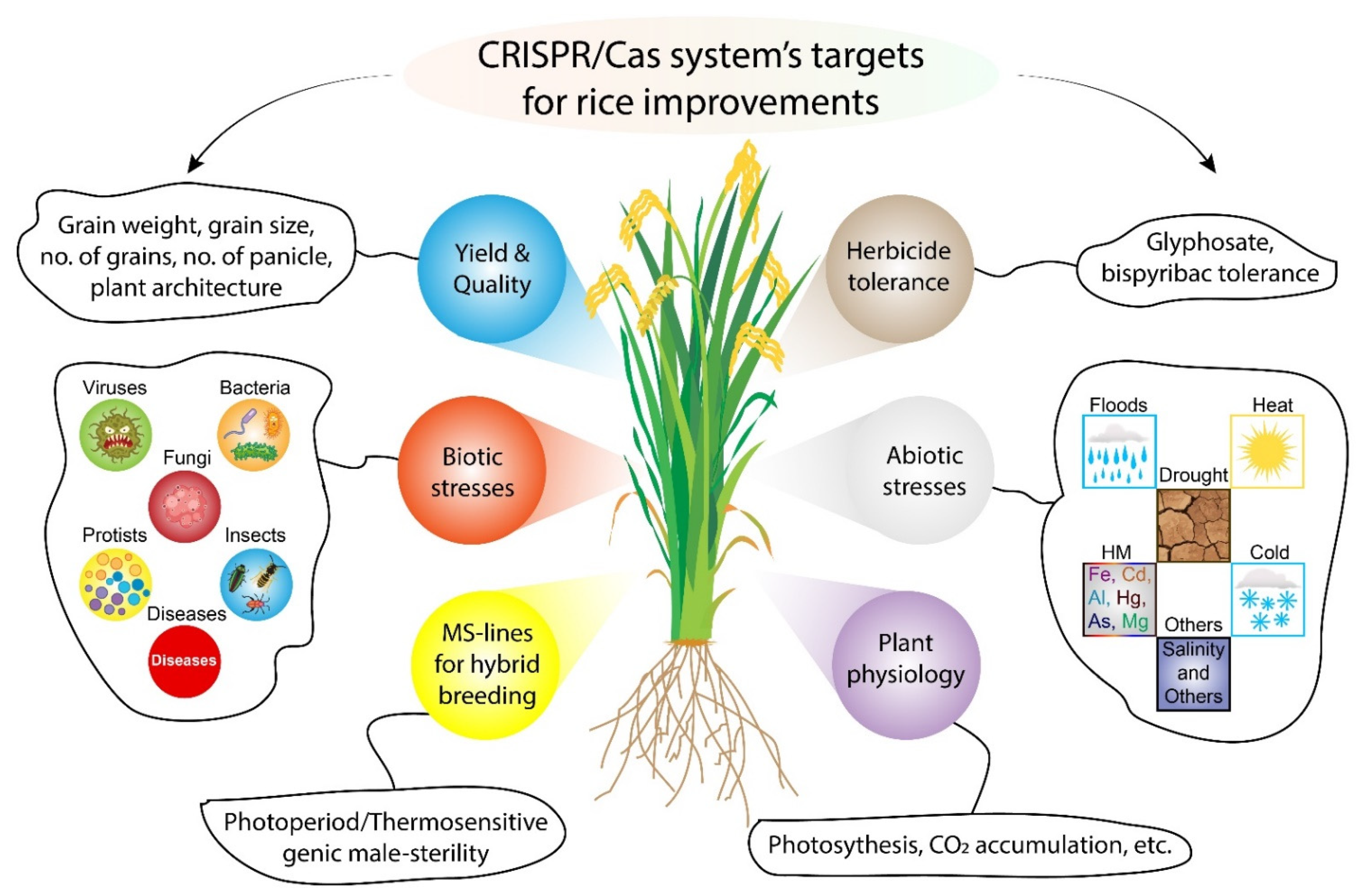
:1. Introduction
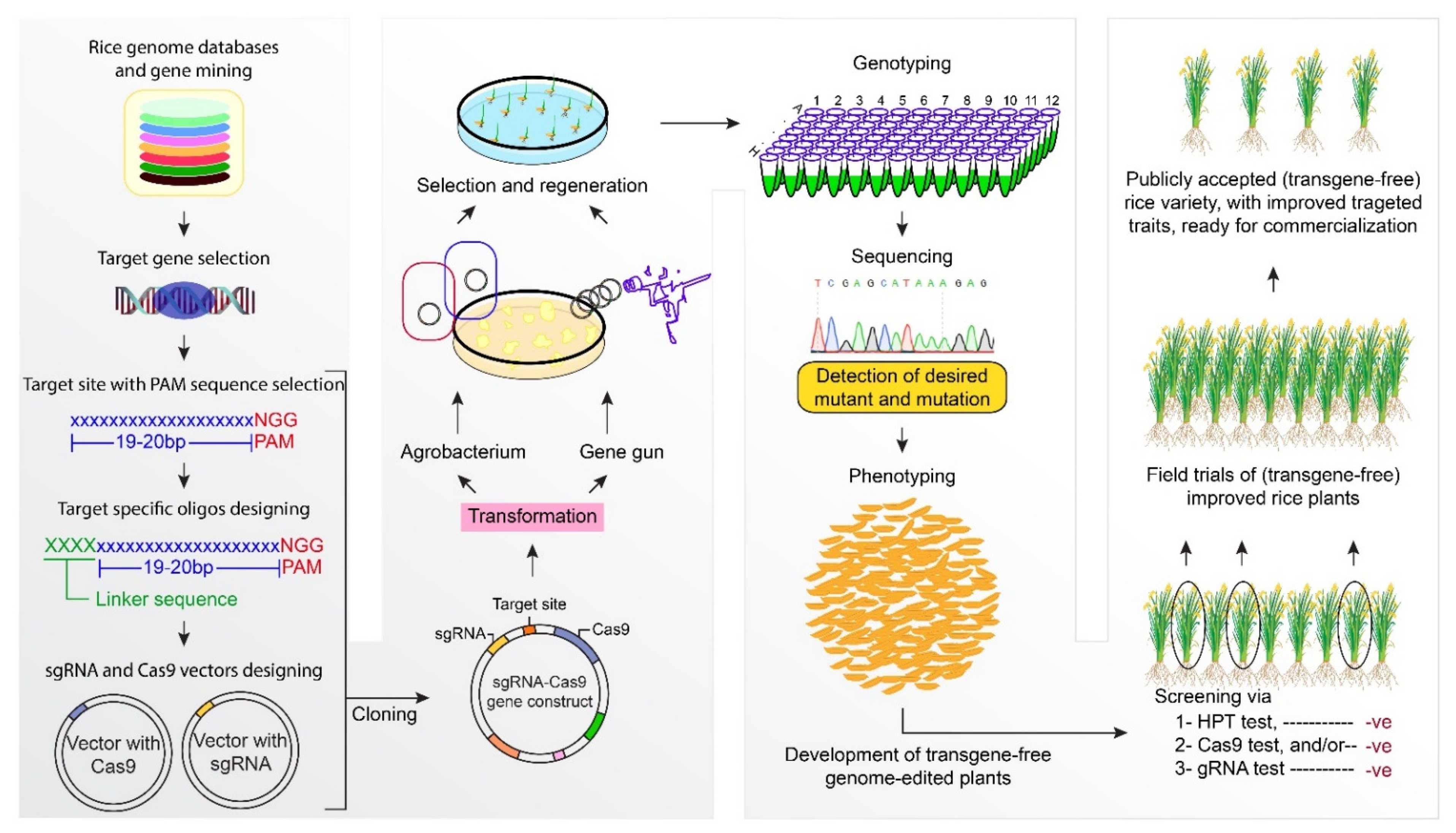
2. CRISPR/Cas9 Based Rice Crop Improvement
2.1. CRISPR/Cas9 for Improving Grain Yield of Rice
2.1.1. Improving the Plant Architecture
2.1.2. Improving the Panicle Architecture
2.1.3. Improving the ABA Signaling Pathway
2.2. CRISPR/Cas9 for Abiotic Stress Tolerant Rice
2.2.1. Targeting the ABA Signaling Pathway
2.2.2. Improving the Leaf Morphology
2.2.3. Targeting the microRNA and Transcription Factors
2.3. CRISPR/Cas9 for Improving Disease Resistance of Rice
2.3.1. Gene Disruption by Targeting the Coding Sequence
2.3.2. Gene Disruption via Promotor Sequence
2.4. CRISPR/Cas9 for Herbicide Resistant Rice
2.5. CRISPR/Cas9 for Improving Rice Quality Parameters
2.5.1. Cooking and Eating Quality Traits
2.5.2. Physical Appearance and Milling Quality
2.5.3. Nutritional Quality Traits
3. Prime Editing and Cas Variants for Rice Crop Improvement
4. Base Editing for Rice Crop Improvement
4.1. Grain Yield and Related Traits
4.2. Grain Quality
4.3. Herbicide Resistance
5. Regulatory Aspects and Risks Associated with Genome Editing
6. Limitations and Solutions
- Disruption/mutation of the targeted gene may cost some fitness, as it can disturb the pathway of the product or any product or element involved in this pathway. A gene has a linkage with many other genes and regulating pathways. This fitness cost may affect genes regulating plant growth and development, deficiency of essential nutrients leading to visual abnormalities. In order to overcome this limitation, the BE method is applicable, targeting a single nucleotide mutation, escaping disruption of other genes, or targeting promoter to generate alleles [116].
- “Off-target mutations” is another major limitation and significant support for improving the CRISPR system [117]. Unintended or undesired DNA modifications created by deceptive gRNA or a gRNA- independent method or non-specific sites fall under off-target mutations [118]. Possible solutions to cope with off-target mutations and production of transgene-free crops are improving the CRISPR system for precise and reliable editing, or developing an approach to identify off-target mutation. Some bioinformatics tools have been established that can detect off-targets, i.e., Cas-OFFinder (http://www.rgenome.net/cas-offinder/, accessed on 29 June 2021) and CCTop (https://crispr.cos.uniheidelberg.de, accessed on 29 June 2021), and also some systems, such as SELEX, IDLV capture, Guide-seq, HTGTS, BLESS, Digenome-seq [119] and DISCOVER-seq [120]. Still, researchers must use these tools according to the requirements due to their specific pros and cons. In contrast, Cas9 proteins have been modified for improved target specificity, including eSpCas9 [121], HF-Cas9 [122], HypaCas9 [123], and Sniper Cas9 [124]. These engineered Cas proteins had an incredible reduction in off-target activity. Cytosine, instead of adenine, is responsible for an off-target mutation in rice hence needs further improvement in tools like base editors [118]. Moreover, PE is also a reliable tool for reducing off-target mutations e.g., targeting of 179 predicted off-target sites with 12 pegRNAs and nCas9 nickase [91] resulted in 0.00~0.23% of off-target mutations.
- Another limiting factor is the commercial adaptation of genome-edited crops in some countries and has been discussed in detail earlier (see regulatory aspects and risks associated with genome editing). Although the GM crop decision and utilization of GETs is pending, there is great potential for robust, efficient, and environmentally friendly breeding for improved variety development.
- Immune DNA/RNA viruses towards eukaryotes is a crucial limitation of the CRISPR/Cas9 system due to outflow and instant replication of viruses [125]. There is a dire need for a widely acceptable CRISPR version, such as Cas13, to overwhelm this issue. Among all three proteins (Cas13a, Cas13b, and Cas13c), Cas13a is referred for its precise, robust RNA replications, and can exert against RNA viruses [126]. In short, Cas13 would be a better choice for targeting viral RNA against CRISPR/Cas9.
- In case of BE, many obstacles, such as high off-target activity, huge editing window, and limited PAM sites limit its efficiency. Several approaches have been used to minimize these limitations, including application of the REPAIR and RESCUE system in plants, alteration of the CBE and ABE system, generating mutations simultaneously at multiple loci in rice [116], such as multiplex BE for crop improvement. RNP approach had also overcome the regulatory obstacles of base editors by increased efficiency to improve agronomic traits.
- PE also exhibits some key issues, including cell type determinants, state of cell, DNA repair mechanisms deciding the fate of productive or unproductive PE or transport of PE protein or pegRNA for regulation of in vivo applications. This issue could be resolved by manipulating DNA repair favoring to replace the edited strand over the non-edited strand subsequent to successful insertion of a 3′ flap, or alternatively use smaller reverse transcriptase enzymes.
7. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Q.; Chen, Q.; Wang, S.; Hong, Y.; Wang, Z. Rice and cold stress: Methods for its evaluation and summary of cold tolerance-related quantitative trait loci. Rice 2014, 7, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Lin, Y.; Chen, H. Improving nutritional quality of rice for human health. Theor. Appl. Genet. 2020, 133, 1397–1413. [Google Scholar] [CrossRef]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield Trends Are Insufficient to Double Global Crop Production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [Green Version]
- Budak, H.; Hussain, B.; Khan, Z.; Ozturk, N.Z.; Ullah, N. From genetics to functional genomics: Improvement in drought signaling and tolerance in wheat. Front. Plant Sci. 2015, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Oladosu, Y.; Rafii, M.Y.; Samuel, C.; Fatai, A.; Magaji, U.; Kareem, I.; Kamarudin, Z.S.; Muhammad, I.; Kolapo, K. Drought Resistance in Rice from Conventional to Molecular Breeding: A Review. Int. J. Mol. Sci. 2019, 20, 3519. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.L.; Che Lah, W.A.; Abd. Kadir, N.; Mustaqim, M.; Rahmat, Z.; Ahmad, A.; Lam, S.D.; Ismail, M.R. Susceptibility and tolerance of rice crop to salt threat: Physiological and metabolic inspections. PLoS ONE 2018, 13, e0192732. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and Applications of CRISPR-Cas9 for Genome Engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Cai, Z.; Xie, W.; Long, T.; Yu, H.; Zhang, Q. Rice functional genomics research: Progress and implications for crop genetic improvement. Biotechnol. Adv. 2012, 30, 1059–1070. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, H.J.; Kim, H.; Cho, S.W.; Kim, J.-S. Targeted genome editing in human cells with zinc finger nucleases constructed via modular assembly. Genome Res. 2009, 19, 1279–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cebrian-Serrano, A.; Davies, B. CRISPR-Cas orthologues and variants: Optimizing the repertoire, specificity and delivery of genome engineering tools. Mamm. Genome 2017, 28, 247–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naeem, M.; Majeed, S.; Hoque, M.Z.; Ahmad, I. Latest Developed Strategies to Minimize the Off-Target Effects in CRISPR-Cas-Mediated Genome Editing. Cells 2020, 9, 1608. [Google Scholar] [CrossRef]
- Hussain, B.; Lucas, S.J.; Budak, H. CRISPR/Cas9 in plants: At play in the genome and at work for crop improvement. Brief. Funct. Genom. 2018, 17, 319–328. [Google Scholar] [CrossRef]
- Adli, M. The CRISPR tool kit for genome editing and beyond. Nat. Commun. 2018, 9, 1911. [Google Scholar] [CrossRef]
- Zhang, Y.; Massel, K.; Godwin, I.D.; Gao, C. Applications and potential of genome editing in crop improvement. Genome Biol. 2018, 19, 210. [Google Scholar] [CrossRef] [PubMed]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, K.; Tao, X.; Yuan, F.; Wang, D.; Zhu, J.-K. Precise A·T to G·C Base Editing in the Rice Genome. Mol. Plant 2018, 11, 627–630. [Google Scholar] [CrossRef] [Green Version]
- Hua, K.; Tao, X.; Liang, W.; Zhang, Z.; Gou, R.; Zhu, J. Simplified adenine base editors improve adenine base editing efficiency in rice. Plant Biotechnol. J. 2020, 18, 770–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimatani, Z.; Kashojiya, S.; Takayama, M.; Terada, R.; Arazoe, T.; Ishii, H.; Teramura, H.; Yamamoto, T.; Komatsu, H.; Miura, K.; et al. Targeted base editing in rice and tomato using a CRISPR-Cas9 cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 441–443. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Zong, Y.; Xue, C.; Wang, S.; Jin, S.; Zhu, Z.; Wang, Y.; Anzalone, A.V.; Raguram, A.; Doman, J.L.; et al. Prime genome editing in rice and wheat. Nat. Biotechnol. 2020, 38, 582–585. [Google Scholar] [CrossRef]
- Xu, W.; Zhang, C.; Yang, Y.; Zhao, S.; Kang, G.; He, X.; Song, J.; Yang, J. Versatile Nucleotides Substitution in Plant Using an Improved Prime Editing System. Mol. Plant 2020, 13, 675–678. [Google Scholar] [CrossRef]
- Tang, X.; Sretenovic, S.; Ren, Q.; Jia, X.; Li, M.; Fan, T.; Yin, D.; Xiang, S.; Guo, Y.; Liu, L.; et al. Plant Prime Editors Enable Precise Gene Editing in Rice Cells. Mol. Plant 2020, 13, 667–670. [Google Scholar] [CrossRef]
- Li, H.; Li, J.; Chen, J.; Yan, L.; Xia, L. Precise Modifications of Both Exogenous and Endogenous Genes in Rice by Prime Editing. Mol. Plant 2020, 13, 671–674. [Google Scholar] [CrossRef]
- Hua, K.; Jiang, Y.; Tao, X.; Zhu, J. Precision genome engineering in rice using prime editing system. Plant Biotechnol. J. 2020, 18, 2167–2169. [Google Scholar] [CrossRef]
- Hussain, B. Modernization in plant breeding approaches for improving biotic stress resistance in crop plants. Turkish J. Agric. For. 2015, 39, 515–530. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, Y.; Xue, D.; Wang, J.; Yan, M.; Liu, G.; Dong, G.; Zeng, D.; Lu, Z.; Zhu, X.; et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 2010, 42, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.; Sasaki, M.; Yamasaki, M. A novel Rice QTL qOPW11 Associated with Panicle Weight Affects Panicle and Plant Architecture. Rice 2018, 11, 53. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Tan, L.; Zhu, Z.; Xiao, L.; Xie, D.; Sun, C. PAY 1 improves plant architecture and enhances grain yield in rice. Plant J. 2015, 83, 528–536. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Cui, Y.; Dong, G.; Feng, A.; Wang, D.; Zhao, C.; Zhang, Y.; Hu, J.; Zeng, D.; Guo, L.; et al. Using CRISPR-Cas9 to generate semi-dwarf rice lines in elite landraces. Sci. Rep. 2019, 9, 19096. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Hu, X.; Liang, G.; Feng, A.; Wang, F.; Ruan, S.; Dong, G.; Shen, L.; Zhang, B.; Chen, D.; et al. Production of novel beneficial alleles of a rice yield-related QTL by CRISPR/Cas9. Plant Biotechnol. J. 2020, 18, 1987–1989. [Google Scholar] [CrossRef]
- Han, Y.; Teng, K.; Nawaz, G.; Feng, X.; Usman, B.; Wang, X.; Luo, L.; Zhao, N.; Liu, Y.; Li, R. Generation of semi-dwarf rice (Oryza sativa L.) lines by CRISPR/Cas9-directed mutagenesis of OsGA20ox2 and proteomic analysis of unveiled changes caused by mutations. 3 Biotech 2019, 9, 387. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, X.; Zhou, Z.; Wu, P.; Fang, M.; Pan, X.; Lin, Q.; Luo, W.; Wu, G.; Li, H. Reassessment of the Four Yield-related Genes Gn1a, DEP1, GS3, and IPA1 in Rice Using a CRISPR/Cas9 System. Front. Plant Sci. 2016, 7, 377. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Li, G.; Sun, H.; Xu, M.; Wang, H.; Ji, J.; Wang, D.; Yuan, C.; Zhao, X. Targeted Mutagenesis of the Rice FW 2.2-Like Gene Family Using the CRISPR/Cas9 System Reveals OsFWL4 as a Regulator of Tiller Number and Plant Yield in Rice. Int. J. Mol. Sci. 2020, 21, 809. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Wen, J.; Zhao, W.; Wang, Q.; Huang, W. Rational Improvement of Rice Yield and Cold Tolerance by Editing the Three Genes OsPIN5b, GS3, and OsMYB30 With the CRISPR–Cas9 System. Front. Plant Sci. 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Zhang, R.; Huang, G.; Li, Y.; Melaku, G.; Zhang, S.; Chen, H.; Zhao, Y.; Zhang, J.; Zhang, Y.; et al. Developing superior alleles of yield genes in rice by artificial mutagenesis using the CRISPR/Cas9 system. Crop J. 2018, 6, 475–481. [Google Scholar] [CrossRef]
- Shen, L.; Wang, C.; Fu, Y.; Wang, J.; Liu, Q.; Zhang, X.; Yan, C.; Qian, Q.; Wang, K. QTL editing confers opposing yield performance in different rice varieties. J. Integr. Plant Biol. 2018, 60, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Yang, Y.; Qin, R.; Li, H.; Qiu, C.; Li, L.; Wei, P.; Yang, J. Rapid improvement of grain weight via highly efficient CRISPR/Cas9-mediated multiplex genome editing in rice. J. Genet. Genom. 2016, 43, 529–532. [Google Scholar] [CrossRef]
- Zhou, J.; Xin, X.; He, Y.; Chen, H.; Li, Q.; Tang, X.; Zhong, Z.; Deng, K.; Zheng, X.; Akher, S.A.; et al. Multiplex QTL editing of grain-related genes improves yield in elite rice varieties. Plant Cell Rep. 2019, 38, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Usman, B.; Nawaz, G.; Zhao, N.; Liao, S.; Qin, B.; Liu, F.; Liu, Y.; Li, R. Programmed editing of rice (Oryza sativa l.) osspl16 gene using crispr/cas9 improves grain yield by modulating the expression of pyruvate enzymes and cell cycle proteins. Int. J. Mol. Sci. 2021, 22, 249. [Google Scholar] [CrossRef]
- Usman, B.; Nawaz, G.; Zhao, N.; Liu, Y.; Li, R. Generation of High Yielding and Fragrant Rice (Oryza sativa L.) Lines by CRISPR/Cas9 Targeted Mutagenesis of Three Homoeologs of Cytochrome P450 Gene Family and OsBADH2 and Transcriptome and Proteome Profiling of Revealed Changes Triggered by Mutations. Plants 2020, 9, 788. [Google Scholar] [CrossRef] [PubMed]
- Usman, B.; Nawaz, G.; Zhao, N.; Liao, S.; Liu, Y.; Li, R. Precise Editing of the OsPYL9 Gene by RNA-Guided Cas9 Nuclease Confers Enhanced Drought Tolerance and Grain Yield in Rice (Oryza sativa L.) by Regulating Circadian Rhythm and Abiotic Stress Responsive Proteins. Int. J. Mol. Sci. 2020, 21, 7854. [Google Scholar] [CrossRef]
- Miao, C.; Xiao, L.; Hua, K.; Zou, C.; Zhao, Y.; Bressan, R.A.; Zhu, J.-K. Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity. Proc. Natl. Acad. Sci. USA 2018, 115, 6058–6063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, B.; KHAN, A.S.; Ali, Z. Genetic variation in wheat germplasm for salinity tolerance at seedling stage: Improved statistical inference. Turkish J. Agric. For. 2015, 39, 182–192. [Google Scholar] [CrossRef]
- Hussain, B.; Lucas, S.J.; Ozturk, L.; Budak, H. Mapping QTLs conferring salt tolerance and micronutrient concentrations at seedling stagein wheat. Sci. Rep. 2017, 7, 15662. [Google Scholar] [CrossRef]
- Ahmad, S.; Sheng, Z.; Jalal, R.S.; Tabassum, J.; Ahmed, F.K.; Hu, S.; Shao, G.; Wei, X.; Abd-Elsalam, K.A.; Hu, P.; et al. CRISPR–Cas technology towards improvement of abiotic stress tolerance in plants. In CRISPR and RNAi Systems; Elsevier: Amsterdam, The Netherlands, 2021; pp. 755–772. [Google Scholar]
- Lou, D.; Wang, H.; Liang, G.; Yu, D. OsSAPK2 Confers Abscisic Acid Sensitivity and Tolerance to Drought Stress in Rice. Front. Plant Sci. 2017, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogata, T.; Ishizaki, T.; Fujita, M.; Fujita, Y. CRISPR/Cas9-targeted mutagenesis of OsERA1 confers enhanced responses to abscisic acid and drought stress and increased primary root growth under nonstressed conditions in rice. PLoS ONE 2020, 15, e0243376. [Google Scholar] [CrossRef]
- Liao, S.; Qin, X.; Luo, L.; Han, Y.; Wang, X.; Usman, B.; Nawaz, G.; Zhao, N.; Liu, Y.; Li, R. CRISPR/Cas9-Induced Mutagenesis of Semi-Rolled Leaf1,2 Confers Curled Leaf Phenotype and Drought Tolerance by Influencing Protein Expression Patterns and ROS Scavenging in Rice (Oryza sativa L.). Agronomy 2019, 9, 728. [Google Scholar] [CrossRef] [Green Version]
- Shahzad, R.; Jamil, S.; Ahmad, S.; Nisar, A.; Amina, Z.; Saleem, S.; Zaffar Iqbal, M.; Muhammad Atif, R.; Wang, X. Harnessing the potential of plant transcription factors in developing climate resilient crops to improve global food security: Current and future perspectives. Saudi J. Biol. Sci. 2021, 28, 2323–2341. [Google Scholar] [CrossRef]
- Santosh Kumar, V.V.; Verma, R.K.; Yadav, S.K.; Yadav, P.; Watts, A.; Rao, M.V.; Chinnusamy, V. CRISPR-Cas9 mediated genome editing of drought and salt tolerance (OsDST) gene in indica mega rice cultivar MTU1010. Physiol. Mol. Biol. Plants 2020, 26, 1099–1110. [Google Scholar] [CrossRef]
- Yue, E.; Cao, H.; Liu, B. OsmiR535, a Potential Genetic Editing Target for Drought and Salinity Stress Tolerance in Oryza sativa. Plants 2020, 9, 1337. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, Y.; Wang, F.; Li, T.; Chen, Z.; Kong, D.; Bi, J.; Zhang, F.; Luo, X.; Wang, J.; et al. Enhanced rice salinity tolerance via CRISPR/Cas9-targeted mutagenesis of the OsRR22 gene. Mol. Breed. 2019, 39, 47. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Que, Z.; Xia, Y.; Tang, N.; Li, D.; He, R.; Cao, M. Knock out of the annexin gene OsAnn3 via CRISPR/Cas9-mediated genome editing decreased cold tolerance in rice. J. Plant Biol. 2017, 60, 539–547. [Google Scholar] [CrossRef]
- Asibi, A.E.; Chai, Q.; Coulter, J.A. Rice Blast: A Disease with Implications for Global Food Security. Agronomy 2019, 9, 451. [Google Scholar] [CrossRef] [Green Version]
- Pandolfi, V.; Neto, J.; Silva, M.; Amorim, L.; Wanderley-Nogueira, A.; Silva, R.; Kido, E.; Crovella, S.; Iseppon, A. Resistance (R) Genes: Applications and Prospects for Plant Biotechnology and Breeding. Curr. Protein Pept. Sci. 2017, 18, 323–334. [Google Scholar] [CrossRef]
- Hussain, B.; Akpınar, B.A.; Alaux, M.; Algharib, A.M.; Sehgal, D.; Ali, Z.; Appels7, R.; Aradottir, G.I.; Batley, J.; Bellec, A.; et al. Wheat genomics and breeding: Bridging the gap. Agrirxiv 2021, 1–57. [Google Scholar] [CrossRef]
- Oliva, R.; Ji, C.; Atienza-Grande, G.; Huguet-Tapia, J.C.; Perez-Quintero, A.; Li, T.; Eom, J.-S.; Li, C.; Nguyen, H.; Liu, B.; et al. Broad-spectrum resistance to bacterial blight in rice using genome editing. Nat. Biotechnol. 2019, 37, 1344–1350. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Xu, X.; Gong, Q.; Li, Z.; Li, Y.; Wang, S.; Yang, Y.; Ma, W.; Liu, L.; Zhu, B.; et al. Engineering Broad-Spectrum Bacterial Blight Resistance by Simultaneously Disrupting Variable TALE-Binding Elements of Multiple Susceptibility Genes in Rice. Mol. Plant 2019, 12, 1434–1446. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Luo, Y.; Vu, N.T.Q.; Shen, S.; Xia, K.; Zhang, M. CRISPR/Cas9-mediated mutation of OsSWEET14 in rice cv. Zhonghua11 confers resistance to Xanthomonas oryzae pv. oryzae without yield penalty. BMC Plant Biol. 2020, 20, 313. [Google Scholar] [CrossRef] [PubMed]
- Zafar, K.; Khan, M.Z.; Amin, I.; Mukhtar, Z.; Yasmin, S.; Arif, M.; Ejaz, K.; Mansoor, S. Precise CRISPR-Cas9 Mediated Genome Editing in Super Basmati Rice for Resistance Against Bacterial Blight by Targeting the Major Susceptibility Gene. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Wei, X.; Sheng, Z.; Hu, P.; Tang, S. CRISPR/Cas9 for development of disease resistance in plants: Recent progress, limitations and future prospects. Brief. Funct. Genom. 2020, 19, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, C.; Liu, P.; Lei, C.; Hao, W.; Gao, Y.; Liu, Y.G.; Zhao, K. Enhanced rice blast resistance by CRISPR/ Cas9-Targeted mutagenesis of the ERF transcription factor gene OsERF922. PLoS ONE 2016, 11, e0154027. [Google Scholar] [CrossRef]
- Foster, A.J.; Martin-Urdiroz, M.; Yan, X.; Wright, H.S.; Soanes, D.M.; Talbot, N.J. CRISPR-Cas9 ribonucleoprotein-mediated co-editing and counterselection in the rice blast fungus. Sci. Rep. 2018, 8, 14355. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Shen, L.; Hu, P.; Liu, Q.; Zhu, X.; Qian, Q.; Wang, K.; Wang, Y. Developing disease-resistant thermosensitive male sterile rice by multiplex gene editing. J. Integr. Plant Biol. 2019, 61, 1201–1205. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, G.; Usman, B.; Peng, H.; Zhao, N.; Yuan, R.; Liu, Y.; Li, R. Knockout of Pi21 by CRISPR/Cas9 and iTRAQ-Based Proteomic Analysis of Mutants Revealed New Insights into M. oryzae Resistance in Elite Rice Line. Genes (Basel) 2020, 11, 735. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-A.; Moon, H.; Park, C.-J. CRISPR/Cas9-targeted mutagenesis of Os8N3 in rice to confer resistance to Xanthomonas oryzae pv. oryzae. Rice 2019, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Macovei, A.; Sevilla, N.R.; Cantos, C.; Jonson, G.B.; Slamet-Loedin, I.; Čermák, T.; Voytas, D.F.; Choi, I.-R.; Chadha-Mohanty, P. Novel alleles of rice eIF4G generated by CRISPR/Cas9-targeted mutagenesis confer resistance to Rice tungro spherical virus. Plant Biotechnol. J. 2018, 16, 1918–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef]
- Li, C.; Li, W.; Zhou, Z.; Chen, H.; Xie, C.; Lin, Y. A new rice breeding method: CRISPR/Cas9 system editing of the Xa13 promoter to cultivate transgene-free bacterial blight-resistant rice. Plant Biotechnol. J. 2020, 18, 313–315. [Google Scholar] [CrossRef] [Green Version]
- Sang, Y.; Mejuto, J.-C.; Xiao, J.; Simal-Gandara, J. Assessment of Glyphosate Impact on the Agrofood Ecosystem. Plants 2021, 10, 405. [Google Scholar] [CrossRef]
- Hussain, B.; Mahmood, S. Development of transgenic cotton for combating biotic and abiotic stresses. In Cotton Production and Uses: Agronomy, Crop Protection, and Postharvest Technologies; Springer: Singapore, 2020; pp. 527–545. ISBN 9789811514722. [Google Scholar]
- Li, J.; Meng, X.; Zong, Y.; Chen, K.; Zhang, H.; Liu, J.; Li, J.; Gao, C. Gene replacements and insertions in rice by intron targeting using CRISPR–Cas9. Nat. Plants 2016, 2, 16139. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, X.; Wu, C.; He, Y.; Ma, Y.; Hou, H.; Guo, X.; Du, W.; Zhao, Y.; Xia, L. Engineering Herbicide-Resistant Rice Plants through CRISPR/Cas9-Mediated Homologous Recombination of Acetolactate Synthase. Mol. Plant 2016, 9, 628–631. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Xu, Y.; Li, W.; Chen, Z.; Wang, J.; Fan, F.; Tao, Y.; Jiang, Y.; Zhu, Q.-H.; Yang, J. Creating a novel herbicide-tolerance OsALS allele using CRISPR/Cas9-mediated gene editing. Crop J. 2021, 9, 305–312. [Google Scholar] [CrossRef]
- Fiaz, S.; Ahmad, S.; Noor, M.; Wang, X.; Younas, A.; Riaz, A.; Riaz, A.; Ali, F. Applications of the CRISPR/Cas9 System for Rice Grain Quality Improvement: Perspectives and Opportunities. Int. J. Mol. Sci. 2019, 20, 888. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Cheng, Z.; Zhang, X.; Guo, X.; Su, N.; Jiang, L.; Mao, L.; Wan, J. Double repression of soluble starch synthase genes SSIIa and SSIIIa in rice ( Oryza sativa L.) uncovers interactive effects on the physicochemical properties of starch. Genome 2011, 54, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Wambugu, P.; Ndjiondjop, M.-N.; Furtado, A.; Henry, R. Sequencing of bulks of segregants allows dissection of genetic control of amylose content in rice. Plant Biotechnol. J. 2018, 16, 100–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, H.; Botella, J.R.; Zhu, J.-K. Generation of new glutinous rice by CRISPR/Cas9-targeted mutagenesis of the Waxy gene in elite rice varieties. J. Integr. Plant Biol. 2018, 60, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Li, Q.; Zhang, C.; Chu, R.; Gu, Z.; Tan, H.; Zhao, D.; Fan, X.; Liu, Q. Creating novel Wx alleles with fine-tuned amylose levels and improved grain quality in rice by promoter editing using CRISPR/Cas9 system. Plant Biotechnol. J. 2020, 18, 2164–2166. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, Y.; Guo, M.; Zhong, C.; Yan, C.; Sun, S. Targeted mutagenesis of amino acid transporter genes for rice quality improvement using the CRISPR/Cas9 system. Crop J. 2020, 8, 457–464. [Google Scholar] [CrossRef]
- Sun, Y.; Jiao, G.; Liu, Z.; Zhang, X.; Li, J.; Guo, X.; Du, W.; Du, J.; Francis, F.; Zhao, Y.; et al. Generation of High-Amylose Rice through CRISPR/Cas9-Mediated Targeted Mutagenesis of Starch Branching Enzymes. Front. Plant Sci. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Ashokkumar, S.; Jaganathan, D.; Ramanathan, V.; Rahman, H.; Palaniswamy, R.; Kambale, R.; Muthurajan, R. Creation of novel alleles of fragrance gene OsBADH2 in rice through CRISPR/Cas9 mediated gene editing. PLoS ONE 2020, 15, e0237018. [Google Scholar] [CrossRef]
- Shufen, C.; Yicong, C.; Baobing, F.; Guiai, J.; Zhonghua, S.; Ju, L.; Shaoqing, T.; Jianlong, W.; Peisong, H.; Xiangjin, W. Editing of Rice Isoamylase Gene ISA1 Provides Insights into Its Function in Starch Formation. Rice Sci. 2019, 26, 77–87. [Google Scholar] [CrossRef]
- Li, Y.; Fan, C.; Xing, Y.; Yun, P.; Luo, L.; Yan, B.; Peng, B.; Xie, W.; Wang, G.; Li, X.; et al. Chalk5 encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice. Nat. Genet. 2014, 46, 398–404. [Google Scholar] [CrossRef]
- Zhao, D.-S.; Li, Q.-F.; Zhang, C.-Q.; Zhang, C.; Yang, Q.-Q.; Pan, L.-X.; Ren, X.-Y.; Lu, J.; Gu, M.-H.; Liu, Q.-Q. GS9 acts as a transcriptional activator to regulate rice grain shape and appearance quality. Nat. Commun. 2018, 9, 1240. [Google Scholar] [CrossRef]
- Liu, J.; Wu, X.; Yao, X.; Yu, R.; Larkin, P.J.; Liu, C.-M. Mutations in the DNA demethylase OsROS1 result in a thickened aleurone and improved nutritional value in rice grains. Proc. Natl. Acad. Sci. USA 2018, 115, 11327–11332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y.; et al. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.-D.; Huang, S.; Konishi, N.; Wang, P.; Chen, J.; Huang, X.-Y.; Ma, J.F.; Zhao, F.-J. Overexpression of the manganese/cadmium transporter OsNRAMP5 reduces cadmium accumulation in rice grain. J. Exp. Bot. 2020, 71, 5705–5715. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Liu, Y.; Liu, Y.; Tan, Y.; Huang, J.; Shu, Q. Mutation of Inositol 1,3,4-trisphosphate 5/6-kinase6 Impairs Plant Growth and Phytic Acid Synthesis in Rice. Plants 2019, 8, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, A.; Saika, H.; Takemura, M.; Misawa, N.; Toki, S. A novel approach to carotenoid accumulation in rice callus by mimicking the cauliflower Orange mutation via genome editing. Rice 2019, 12, 81. [Google Scholar] [CrossRef] [PubMed]
- Dong, O.X.; Yu, S.; Jain, R.; Zhang, N.; Duong, P.Q.; Butler, C.; Li, Y.; Lipzen, A.; Martin, J.A.; Barry, K.W.; et al. Marker-free carotenoid-enriched rice generated through targeted gene insertion using CRISPR-Cas9. Nat. Commun. 2020, 11, 1178. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Lin, Q.; Luo, Y.; Zhu, Z.; Liu, G.; Li, Y.; Chen, K.; Qiu, J.-L.; Gao, C. Genome-wide specificity of prime editors in plants. Nat. Biotechnol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Li, J.; Liu, X.; Shan, T.; Qin, R.; Wei, P. Development of Plant Prime-Editing Systems for Precise Genome Editing. Plant Commun. 2020, 1, 100043. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Liang, J.; Li, Z.; Gong, B.; Li, J. Multiplex and optimization of dCas9-TV-mediated gene activation in plants. J. Integr. Plant Biol. 2021, 63, 634–645. [Google Scholar] [CrossRef]
- Lowder, L.G.; Zhou, J.; Zhang, Y.; Malzahn, A.; Zhong, Z.; Hsieh, T.-F.; Voytas, D.F.; Zhang, Y.; Qi, Y. Robust Transcriptional Activation in Plants Using Multiplexed CRISPR-Act2.0 and mTALE-Act Systems. Mol. Plant 2018, 11, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zong, Y.; Lin, Q.; Zhang, H.; Chai, Z.; Zhang, D.; Chen, K.; Qiu, J.-L.; Gao, C. Precise, predictable multi-nucleotide deletions in rice and wheat using APOBEC–Cas9. Nat. Biotechnol. 2020, 38, 1460–1465. [Google Scholar] [CrossRef]
- Endo, A.; Masafumi, M.; Kaya, H.; Toki, S. Efficient targeted mutagenesis of rice and tobacco genomes using Cpf1 from Francisella novicida. Sci. Rep. 2016, 6, 38169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Z.; Zhang, Y.; You, Q.; Tang, X.; Ren, Q.; Liu, S.; Yang, L.; Wang, Y.; Liu, X.; Liu, B.; et al. Plant Genome Editing Using FnCpf1 and LbCpf1 Nucleases at Redefined and Altered PAM Sites. Mol. Plant 2018, 11, 999–1002. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Mao, Y.; Lu, Y.; Tao, X.; Zhu, J. Multiplex Gene Editing in Rice Using the CRISPR-Cpf1 System. Mol. Plant 2017, 10, 1011–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol. 2020, 38, 824–844. [Google Scholar] [CrossRef] [PubMed]
- Hua, K.; Tao, X.; Han, P.; Wang, R.; Zhu, J.-K. Genome Engineering in Rice Using Cas9 Variants that Recognize NG PAM Sequences. Mol. Plant 2019, 12, 1003–1014. [Google Scholar] [CrossRef] [PubMed]
- Monsur, M.B.; Shao, G.; Lv, Y.; Ahmad, S.; Wei, X.; Hu, P.; Tang, S. Base Editing: The Ever Expanding Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) Tool Kit for Precise Genome Editing in Plants. Genes (Basel) 2020, 11, 466. [Google Scholar] [CrossRef]
- Hua, K.; Tao, X.; Zhu, J.-K. Expanding the base editing scope in rice by using Cas9 variants. Plant Biotechnol. J. 2019, 17, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Zong, Y.; Wang, Y.; Li, C.; Zhang, R.; Chen, K.; Ran, Y.; Qiu, J.-L.; Wang, D.; Gao, C. Precise base editing in rice, wheat and maize with a Cas9-cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 438–440. [Google Scholar] [CrossRef]
- Lu, Y.; Zhu, J.-K. Precise Editing of a Target Base in the Rice Genome Using a Modified CRISPR/Cas9 System. Mol. Plant 2017, 10, 523–525. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Sun, Y.; Du, J.; Zhao, Y.; Xia, L. Generation of Targeted Point Mutations in Rice by a Modified CRISPR/Cas9 System. Mol. Plant 2017, 10, 526–529. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Lin, Q.; Li, X.; Wang, F.; Chen, Z.; Wang, J.; Li, W.; Fan, F.; Tao, Y.; Jiang, Y.; et al. Fine-tuning the amylose content of rice by precise base editing of the Wx gene. Plant Biotechnol. J. 2021, 19, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Y.; Li, S.; Ren, B.; Yan, F.; Spetz, C.; Li, X.; Zhou, X.; Zhou, H. Base-Editing-Mediated Artificial Evolution of OsALS1 In Planta to Develop Novel Herbicide-Tolerant Rice Germplasms. Mol. Plant 2020, 13, 565–572. [Google Scholar] [CrossRef]
- Zhang, R.; Chen, S.; Meng, X.; Chai, Z.; Wang, D.; Yuan, Y.; Chen, K.; Jiang, L.; Li, J.; Gao, C. Generating broad-spectrum tolerance to ALS-inhibiting herbicides in rice by base editing. Sci. China Life Sci. 2020. [Google Scholar] [CrossRef]
- Liu, X.; Qin, R.; Li, J.; Liao, S.; Shan, T.; Xu, R.; Wu, D.; Wei, P. A CRISPR-Cas9-mediated domain-specific base-editing screen enables functional assessment of ACCase variants in rice. Plant Biotechnol. J. 2020, 18, 1845–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Shahzad, R.; Jamil, S.; Tabassum, J.; Chaudhary, M.A.M.; Atif, R.M.; Iqbal, M.M.; Monsur, M.B.; Lv, Y.; Sheng, Z.; et al. Regulatory aspects, risk assessment, and toxicity associated with RNAi and CRISPR methods. In CRISPR and RNAi Systems; Elsevier: Amsterdam, The Netherlands, 2021; pp. 687–721. [Google Scholar]
- Podevin, N.; Davies, H.V.; Hartung, F.; Nogué, F.; Casacuberta, J.M. Site-directed nucleases: A paradigm shift in predictable, knowledge-based plant breeding. Trends Biotechnol. 2013, 31, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Lema, M.A. Regulatory aspects of gene editing in Argentina. Transgenic Res. 2019, 28, 147–150. [Google Scholar] [CrossRef]
- Gao, W.; Xu, W.-T.; Huang, K.-L.; Guo, M.; Luo, Y.-B. Risk analysis for genome editing-derived food safety in China. Food Control 2018, 84, 128–137. [Google Scholar] [CrossRef]
- Normile, D. Gene-edited foods are safe, Japanese panel concludes. Science 2019. [Google Scholar] [CrossRef]
- Thygesen, P. Clarifying the regulation of genome editing in Australia: Situation for genetically modified organisms. Transgenic Res. 2019, 28, 151–159. [Google Scholar] [CrossRef]
- Yan, F.; Kuang, Y.; Ren, B.; Wang, J.; Zhang, D.; Lin, H.; Yang, B.; Zhou, X.; Zhou, H. Highly Efficient A·T to G·C Base Editing by Cas9n-Guided tRNA Adenosine Deaminase in Rice. Mol. Plant 2018, 11, 631–634. [Google Scholar] [CrossRef] [Green Version]
- Hahn, F.; Nekrasov, V. CRISPR/Cas precision: Do we need to worry about off-targeting in plants? Plant Cell Rep. 2019, 38, 437–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Zong, Y.; Gao, Q.; Zhu, Z.; Wang, Y.; Qin, P.; Liang, C.; Wang, D.; Qiu, J.-L.; Zhang, F.; et al. Cytosine, but not adenine, base editors induce genome-wide off-target mutations in rice. Science 2019, eaaw7166. [Google Scholar] [CrossRef] [PubMed]
- Koo, T.; Lee, J.; Kim, J.-S. Measuring and Reducing Off-Target Activities of Programmable Nucleases Including CRISPR-Cas9. Mol. Cells 2015, 38, 475–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wienert, B.; Wyman, S.K.; Richardson, C.D.; Yeh, C.D.; Akcakaya, P.; Porritt, M.J.; Morlock, M.; Vu, J.T.; Kazane, K.R.; Watry, H.L.; et al. Unbiased detection of CRISPR off-targets in vivo using DISCOVER-Seq. Science 2019, 364, 286–289. [Google Scholar] [CrossRef]
- Slaymaker, I.M.; Gao, L.; Zetsche, B.; Scott, D.A.; Yan, W.X.; Zhang, F. Rationally engineered Cas9 nucleases with improved specificity. Science 2016, 351, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Kleinstiver, B.P.; Pattanayak, V.; Prew, M.S.; Tsai, S.Q.; Nguyen, N.T.; Zheng, Z.; Joung, J.K. High-fidelity CRISPR–Cas9 nucleases with no detectable genome-wide off-target effects. Nature 2016, 529, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.S.; Dagdas, Y.S.; Kleinstiver, B.P.; Welch, M.M.; Sousa, A.A.; Harrington, L.B.; Sternberg, S.H.; Joung, J.K.; Yildiz, A.; Doudna, J.A. Enhanced proofreading governs CRISPR–Cas9 targeting accuracy. Nature 2017, 550, 407–410. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.K.; Jeong, E.; Lee, J.; Jung, M.; Shin, E.; Kim, Y.; Lee, K.; Jung, I.; Kim, D.; Kim, S.; et al. Directed evolution of CRISPR-Cas9 to increase its specificity. Nat. Commun. 2018, 9, 3048. [Google Scholar] [CrossRef] [Green Version]
- Mehta, D.; Stürchler, A.; Anjanappa, R.B.; Zaidi, S.S.-A.; Hirsch-Hoffmann, M.; Gruissem, W.; Vanderschuren, H. Linking CRISPR-Cas9 interference in cassava to the evolution of editing-resistant geminiviruses. Genome Biol. 2019, 20, 80. [Google Scholar] [CrossRef] [Green Version]
- Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Ding, S.; Mahfouz, M. RNA virus interference via CRISPR/Cas13a system in plants. Genome Biol. 2018, 19, 1. [Google Scholar] [CrossRef]
- Gentzel, I.N.; Park, C.H.; Bellizzi, M.; Xiao, G.; Gadhave, K.R.; Murphree, C.; Yang, Q.; LaMantia, J.; Redinbaugh, M.G.; Balint-Kurti, P.; et al. A CRISPR/dCas9 toolkit for functional analysis of maize genes. Plant Methods 2020, 16, 133. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.; Yang, W.; Zeng, X.; Xu, X.; Xu, Y.; Fan, X.; Luo, M.; Tian, C.; Xia, K.; Zhang, M. A Rice Autophagy Gene OsATG8b Is Involved in Nitrogen Remobilization and Control of Grain Quality. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Shang, F.; Chao, X.; Meng, K.; Meng, X.; Li, Q.; Chen, L.; Wang, J. Fine Mapping of a Grain Shape Gene from a Rice Landrace Longliheinuo-Dwarf (Oryza sativa L. ssp. japonica). Agronomy 2020, 10, 380. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.; Gao, Y.; Li, Y.; Ruan, B.; Yang, S.; Liu, C.; Zhang, B.; Jiang, H.; Fang, G.; Ding, S.; et al. Genetic Analysis for Cooking and Eating Quality of Super Rice and Fine Mapping of a Novel Locus qGC10 for Gel Consistency. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Khahani, B.; Tavakol, E.; Shariati, V.; Rossini, L. Meta-QTL and ortho-MQTL analyses identified genomic regions controlling rice yield, yield-related traits and root architecture under water deficit conditions. Sci. Rep. 2021, 11, 6942. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.; Ontoy, J.; Subudhi, P. Meta-Analysis of Quantitative Trait Loci Associated with Seedling-Stage Salt Tolerance in Rice (Oryza sativa L.). Plants 2019, 8, 33. [Google Scholar] [CrossRef] [Green Version]
- Raza, Q.; Riaz, A.; Bashir, K.; Sabar, M. Reproductive tissues-specific meta-QTLs and candidate genes for development of heat-tolerant rice cultivars. Plant Mol. Biol. 2020, 104, 97–112. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targeted Trait | Targeted Gene/s | Cas9 Promoter/S | sgRNA Promoter/S | Improved Trait/s in Mutants | Ref. |
|---|---|---|---|---|---|
| Plant Architecture | SD1 | 2 × 35S pro | gRNA1SD1 gRNA3SE5 | Grain yield, plant architecture, semi-dwarf plants, resistance to lodging | [28] |
| OsGA20ox2 | Pubi-H | OsU6a OsU6b | Grain yield, plant architecture, semi-dwarf plants, reduced gibberellins and flag leaf length | [30] | |
| SCM1/SD1, SCM3/OsTB1/FC1, SCM2/APO1 | 2 × 35S pro CaMV | gRNA1, gRNA2, gRNA3, gRNA4, gRNA5, gRNA6 | Plant architecture, number of tillers, panicle architecture, larger panicles, stem cross-section area | [29] | |
| OsFWL4 | Maize Ubi1 | OsU6 | Grain yield, plant architecture, number of tillers, flag leaf area, grain length, number of cells in flag leaf | [32] | |
| IPA1 | Maize Ubi1 | U6a | Grain yield, plant architecture, number of tillers, reduced plant height | [31] | |
| Panicle Architecture | IPA1, GS3, DEP1, Gn1a | Maize Ubi1 | U6a | Grain yield, plant architecture, panicle architecture, number of tillers, grain size, dense erect panicles, grain number | [31] |
| GS3, OsGW2, Gn1a | p35S | OsU6 OsU3 | Grain yield, grain size, grain weight, number of grains per panicle | [37] | |
| OsPIN5b, GS3 | 2 × 35S pro Pubi-H | OsU6a | Grain yield, panicle architecture, panicle length, grain size | [33] | |
| GW2, 5 and 6 | pUBQ | OsU3, OsU6 TaU3 | Grain yield, grain weight | [36] | |
| OsSPL16/qGW8 | 2 × 35S pro Pubi | OsU6a | Grain yield, grain weight, grain size | [38] | |
| Gn1a, GS3 | 2 × 35S pro | U3 | Grain yield, panicle architecture, number of grains per panicle, grain size | [35] | |
| Gn1a, DEP1 | 2 × 35S pro | OsU3 | Grain yield, panicle architecture, panicle orientation, number of grains per panicle | [34] | |
| Cytochrome P450, OsBADH2 | Pubi-H | U6a U6b U6c U3m | Grain yield, grain size, aroma (2-acetyl-1-pyrroline (2AP) content) | [39] | |
| ABA Signaling Pathway | PYL1, PYL4, PYL6 | Maize Ubi1 | OsU6 OsU3 | Number of grains, grain yield | [41] |
| OsPYL9 | PubiH | OsU6a OsU6b | Grain yield under normal and limited water availability | [40] |
| Stress | Edited Gene/S | Cas9 Promoter/S | sgRNA Promoter/S | Improved Traits in Mutants | Ref. |
|---|---|---|---|---|---|
| Drought | OsSAPK2 | Pubi-H | U3 | Reduced drought, salinity, and osmotic stress tolerance; role of gene in ROS scavenging, stomatal conductance and ABA signaling | [45] |
| OsPYL9 | PubiH | OsU6a OsU6b | Drought tolerance; grain yield, antioxidant activities, chlorophyll content, ABA accumulation, leaf cuticle wax, survival rate, stomatal conductance, transpiration rate | [40] | |
| OsERA1 | Not defined | pCAMBIA1300 | Drought tolerance, stomatal conductance, increased sensitivity to ABA. | [46] | |
| OsSRL1, OsSRL2 | Pubi-H | U6a U6b U6c U3m | Improved drought tolerance; Reduced number of stomata, stomatal conductance, transpiration rate and malondialdehyde (MDA) content; Improved panicle number, abscisic acid (ABA) content, catalase (CAT), superoxide dismutase (SOD) and survival rate | [47] | |
| DST | OsUBQ | OsU3 | Drought tolerance, leaf architecture, reduced stomatal density, enhanced leaf water retention | [49] | |
| OsmiR535 | UBI 35S pro | OsU3 OsU6 | Drought tolerance, ABA insensitivity, number of lateral roots (73% more), shoot length (30% longer), primary root length | [50] | |
| Salinity and Osmotic Stress | OsSAPK2 | Pubi-H | U3 | Reduced salinity and osmotic stress tolerance, role of gene in ROS scavenging | [45] |
| OsRR22 | 2 × 35S pro Pubi-H | OsU6a | Salinity tolerance, shoot length, shoot fresh and dry weight | [51] | |
| DST | OsUBQ | OsU3 | Salinity tolerance, osmotic tolerance | [49] | |
| OsmiR535 | UBI 35S pro | OsU3 OsU6 | Salinity tolerance, osmotic tolerance, shoot length (86.8%), number of lateral roots (514% as compared with line overexpressing MIR535), primary root length (35.8%) | [50] | |
| Cold Stress | OsAnn3 | UBI 35S pro | U3 | Response to cold tolerance | [52] |
| OsMYB30 | 2 × 35S pro Pubi-H | OsU6a | Cold tolerance | [33] |
| Pathogen | Improved Disease/Pathogen Resistance | Targeted Gene/S | Cas9 Promoter/S | sgRNA Promoter/S | Ref. |
|---|---|---|---|---|---|
| Fungi | Rice blast (Magnaporthe oryzae) | OsERF922 | 2 × 35S pro Pubi-H | OsU6a | [61] |
| OsALB1, OsRSY1, | TrpC, TEF1 | SNR52, U6–1, U6-2 | [62] | ||
| OsPi21 | PubiH | OsU6a, OsU3 | [63] | ||
| OsPi21 | PubiH | OsU6a, OsU6b | [64] | ||
| Bacteria | Bacterial leaf blight (Xanthomonas oryzae pv. Oryzae) | OsSWEET14, OsSWEET11 | CaMV35S | U6 | [67] |
| OsSWEET11 or Os8N3 | 35S-p | OsU6a | [65] | ||
| OsXa13/SWEET11 | PubiH | OsU6a, OsU3 | [63] | ||
| OsSWEET11, OsSWEET13, OsSWEET14 | ZmUbiP | U6 | [56] | ||
| OsSWEET11, OsSWEET14 | 35S CaMV | SW11, SW14 | [57] | ||
| OsSWEET14 | Pubi or P35S | OsU3, OsU6b, OsU6c | [58] | ||
| OsSWEET14 | 35S, Ubi | OsU3 | [59] | ||
| OsXa13/ SWEET11 | 35S, Ubi | U3, U6a | [68] | ||
| Virus | Rice tungro spherical virus (RTSV) | eIF4G | ZmUBI1, CaMV35S | TaU6 | [66] |
| Quality Traits | Cultivar Background | Targeted Gene | Gene Function | Cas9 Promoter | sgRNA Promoter | Results | Ref. |
|---|---|---|---|---|---|---|---|
| Eating andcookingquality | Yanggeng-158 Nangeng-9108 Wuyungeng-30 | OsAAP6 OsAAP10 | Amino acid transporter for GPC | CaMV35S | OsU3 | Improved eating and cooking quality | [79] |
| Zhonghua11, XS134 | OsWaxy | GBSS (amylose synthesis) | CaMV35S | OsU6 | Decrease in amylose content (glutinous rice) | [78] | |
| XS134 (Japonica) | OsWaxy | GBSS (amylose synthesis) | CaMV35S | OsU6 | Decrease in amylose content (glutinous rice) | [77] | |
| Kitaake (Japonica) | OsBEIand OsBEIIb | Starch branching enzyme | pCXUN | OsU3 | High amylose content | [80] | |
| Zhonghua11 | ISA1 | Starch (isoamylase-type) debranching enzymes | CaMV 35S | OsU6 | Reduced amylose content; increased total soluble sugar | [82] | |
| Indica | BADH2 | Betaine aldehyde dehydrogenase (fragrant rice) | OsUbi | OsU6a | Enhanced fragrance | [81] | |
| Zhonghua 11 (Japonica) | GS9, DEP1 | Grain size, Panicle architecture | OsUbi | OsU6a | Slender grain shape, less chalkiness | [84] | |
| IR-96 | Cyt P450 homoeologs, B OsBADH2 | Grain yield and fragrance | Pubi | OsU6a, OsU6b, OsU6c, OsU3m | Increased grain size, and fragrance | [39] | |
| Physical and appearance quality | Indica (VP4892) | OsSPL16/GW8 | Grain size | pUbi | OsU6aOsU6b | Increased grain size | [38] |
| Zhonghua-11 (Japonica) | GS3/Gn la, | Grain size-3/grain number la | OsUbi | OsU6a | Increased grain length | [31] | |
| Japonica | GS3 | Grain size-3 | 2 × 35S Pubi-H | OsU6a | Increased grain size | [33] | |
| Nipponbare (Japonica) | GW2/GW5/ TGW6 | Grain weight | OsUbi | OsU3, OsU6, TaU3 | Increased grain length and width | [36] | |
| Japonica | OsFWL | Grain Length | Maize Ubi1 | OsU6 | Increased grain length | [32] | |
| Nutritional Quality | Indica | OsNramp5 | Cd accumulation | ZmUBI | OsU6a | Low Cd in grains | [87] |
| Indica (Huazhan and Longke 638S) | OsNramp5 | Cd accumulation | Pubi-H | OsU6a | Low Cd in grains | [86] | |
| Rice Protoplast | OsOr | β-carotene synthesis | 2 × 35S | OsU6-2 | Increased β-carotene (provitamin A) content | [89] | |
| Kitaake | SSU-crtI, ZmPsy | β-carotene synthesis | Increased β-carotene (provitamin A) content | [90] |
| Systems | Cultivar Used | Gene Name | Gene Function | Cas9 Promoter | sgRNA Promoter | Results | Ref. |
|---|---|---|---|---|---|---|---|
| dCas9 | Zhonghua 11 | OsGW7 OsER1 | Grain size and shape, ethylene upregulation | CaMV35S | OsU6a | Multiplex genome editing | [93] |
| Rice protoplast | OsSPL14, OsIPA, OsGRF1 | Senescence, plant architecture | Ubi | Generate larger deletions | [94] | ||
| Cas9 with APOBEC | Rice protoplast | OsAAT OsNRT1.1B OsCDC48 | senescence, cell death | Ubi | CaMV | Generate short and larger deletions | [95] |
| Cas12a/Cpf1 FnCpf1 and CRISPR/ LbCpf1 | Nipponbare | ALS (Acetolactate synthase) | Synthesis of branched chain amino acids | ZmUBI | OsU6 | Loss of ALS activity, plant death | [96] |
| Rice protoplast | DEP1 | Dense and erect Panicle | CaMV35S | OsU6 | Scattered panicle | [97] | |
| Rice protoplast | OsBEL, OsPDS OsEPSPS OsBEL OsRLK | Bentazon-sensitive-lethal, Phytoene Desaturase | CaMV35S | OsU6 | Multiplex gene editing (Albino phenotype) | [98] | |
| Prime editors | Rice protoplast | OsALS APO1 | Acetolactate synthase, Panicle organization | CaMV35S | OsU6 | Resistant to imidazolinone herbicides | [23] |
| Nipponbare | OsPDS1 OsACC1 OsWx1 | Herbicide resistance, amylose | Ubi-1 CaMV35S | OsU3 | Enhanced herbicide tolerance | [92] | |
| Rice protoplast | OsALS, OsACC OsDEP1 | Nitrogen use efficiency Herbicide tolerance | ZmUbi1 OsUbq | OsU6a OsU3 | Nucleotide substitution, herbicide resistance | [20] | |
| Rice protoplast | OsALS, OsKO2, OsDEP1, PDS | Panicle Architecture | ZmUbi1 | OsU6 OsU3 | Novel prime editing, dense panicle architecture | [21] |
| Gene Name | Gene Function | Base Editing Tool | PAM | Editing Window (nt) | Target Trait | Ref. |
|---|---|---|---|---|---|---|
| OsCDC48, OsNRT1.1B OsSPL14 | senescence, cell death, plant architecture | pnCas9-PBE | CGG | 3 to 9 | Yield | [103] |
| SPL14, SPL17, SPL16, SPL18 | Grain weight, size, shape, quality, number | ABE-P1 ABE-P2 ABE-P3 ABE-P4 ABE-P5 | GAG CAG CGA GGA AGCG GGCG | 3 to 15 | Yield | [102] |
| SLR1, SPL14, SPL16, SPL18, SPL17 | Grain weight, size, shape, quality, number | ABE-P1 ABE-P2 | NNGRRT | 4 to 9 | Yield | [17] |
| NRT1.1B | Nitrogen transporter | APOBEC1-XTEN-Cas9(D10A) | AGG GGG | 5 | High nitrogen use Efficiency | [104] |
| SBEIIb | Starch branching enzyme | CBE | CCT | 5 | High amylose | [105] |
| Wx or GBSSI | Starch synthesis | CBE | CCA CGG | 4 | High amylose | [106] |
| OsALS1 | ABC transporter | BEMGE | CGG CAG | 5–7 | Herbicide resistance | [107] |
| OsALS1 | ABC transporter | CBE | CGG CCT | 3–5 | Herbicide resistance | [108] |
| OsACC | Herbicide resistance | eABE eBE3 | TGG | 3 to 9 | Herbicide resistance | [109] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabassum, J.; Ahmad, S.; Hussain, B.; Mawia, A.M.; Zeb, A.; Ju, L. Applications and Potential of Genome-Editing Systems in Rice Improvement: Current and Future Perspectives. Agronomy 2021, 11, 1359. https://doi.org/10.3390/agronomy11071359
Tabassum J, Ahmad S, Hussain B, Mawia AM, Zeb A, Ju L. Applications and Potential of Genome-Editing Systems in Rice Improvement: Current and Future Perspectives. Agronomy. 2021; 11(7):1359. https://doi.org/10.3390/agronomy11071359
Chicago/Turabian StyleTabassum, Javaria, Shakeel Ahmad, Babar Hussain, Amos Musyoki Mawia, Aqib Zeb, and Luo Ju. 2021. "Applications and Potential of Genome-Editing Systems in Rice Improvement: Current and Future Perspectives" Agronomy 11, no. 7: 1359. https://doi.org/10.3390/agronomy11071359
APA StyleTabassum, J., Ahmad, S., Hussain, B., Mawia, A. M., Zeb, A., & Ju, L. (2021). Applications and Potential of Genome-Editing Systems in Rice Improvement: Current and Future Perspectives. Agronomy, 11(7), 1359. https://doi.org/10.3390/agronomy11071359