
How Do Different Cocoa Genotypes Deal with Increased Radiation? An Analysis of Water Relation, Diffusive and Biochemical Components at the Leaf Level

 , and
, and 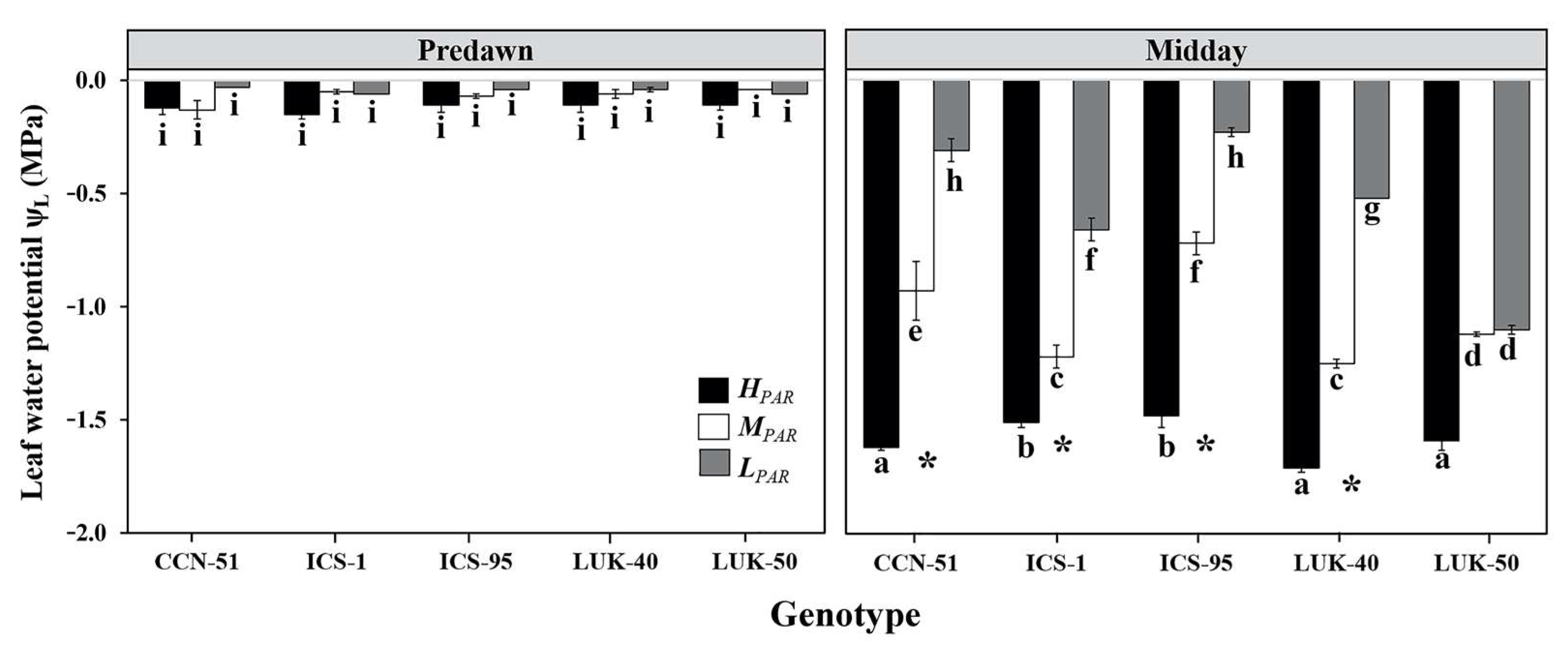{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Leaf Water Potential (ΨL) and Specific Leaf Area
2.3. Photosynthetic Light- and CO2-Response Curves of Different Genotypes of Cacao Grown under Varying Levels of Radiation
2.4. Chlorophyll (Chla) Parameters of Different Genotypes of Cacao Grown under Varying Levels of Radiation
2.5. Biochemical Assays
2.6. Data Analysis
3. Results
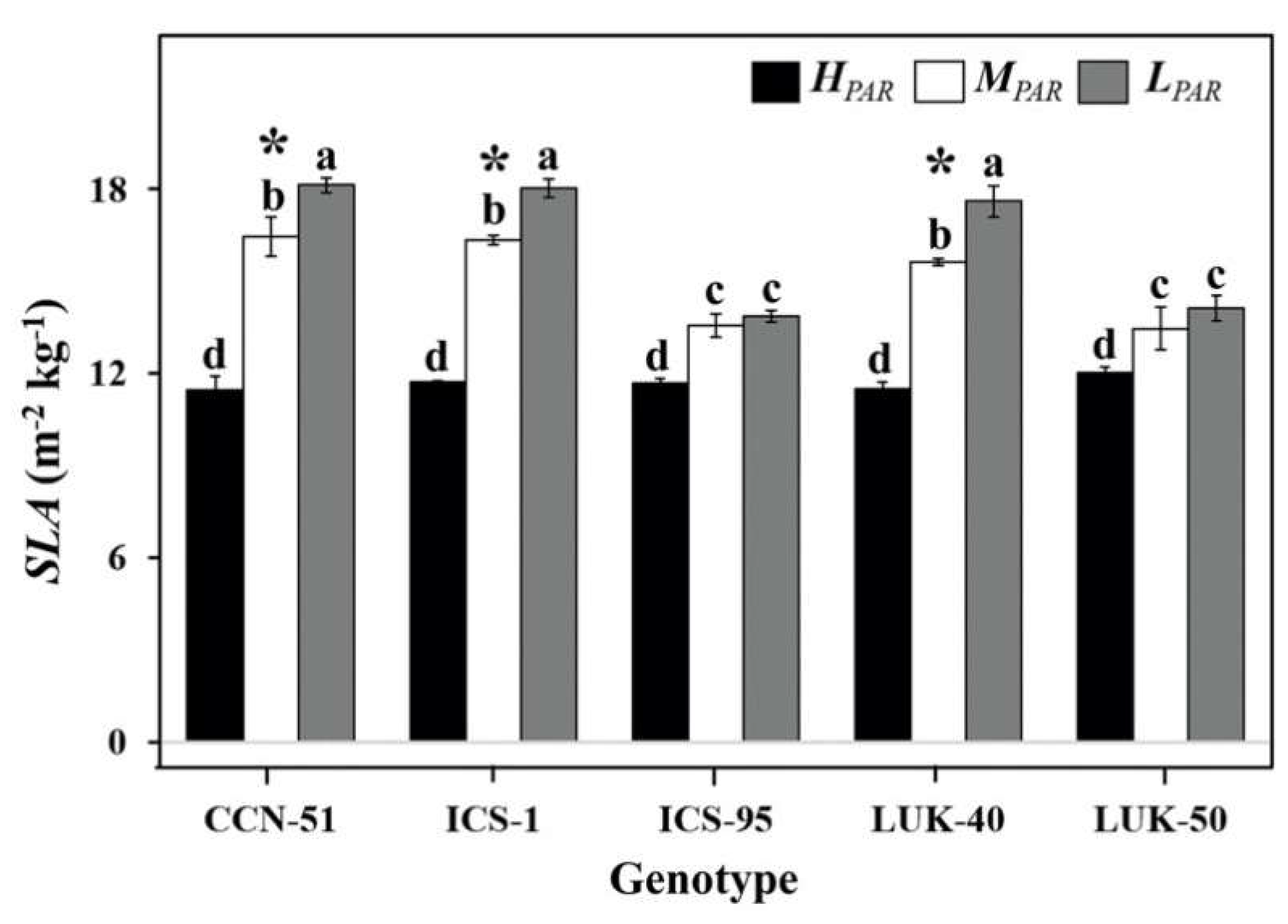
3.1. Leaf Water Potential and Specific Leaf Area
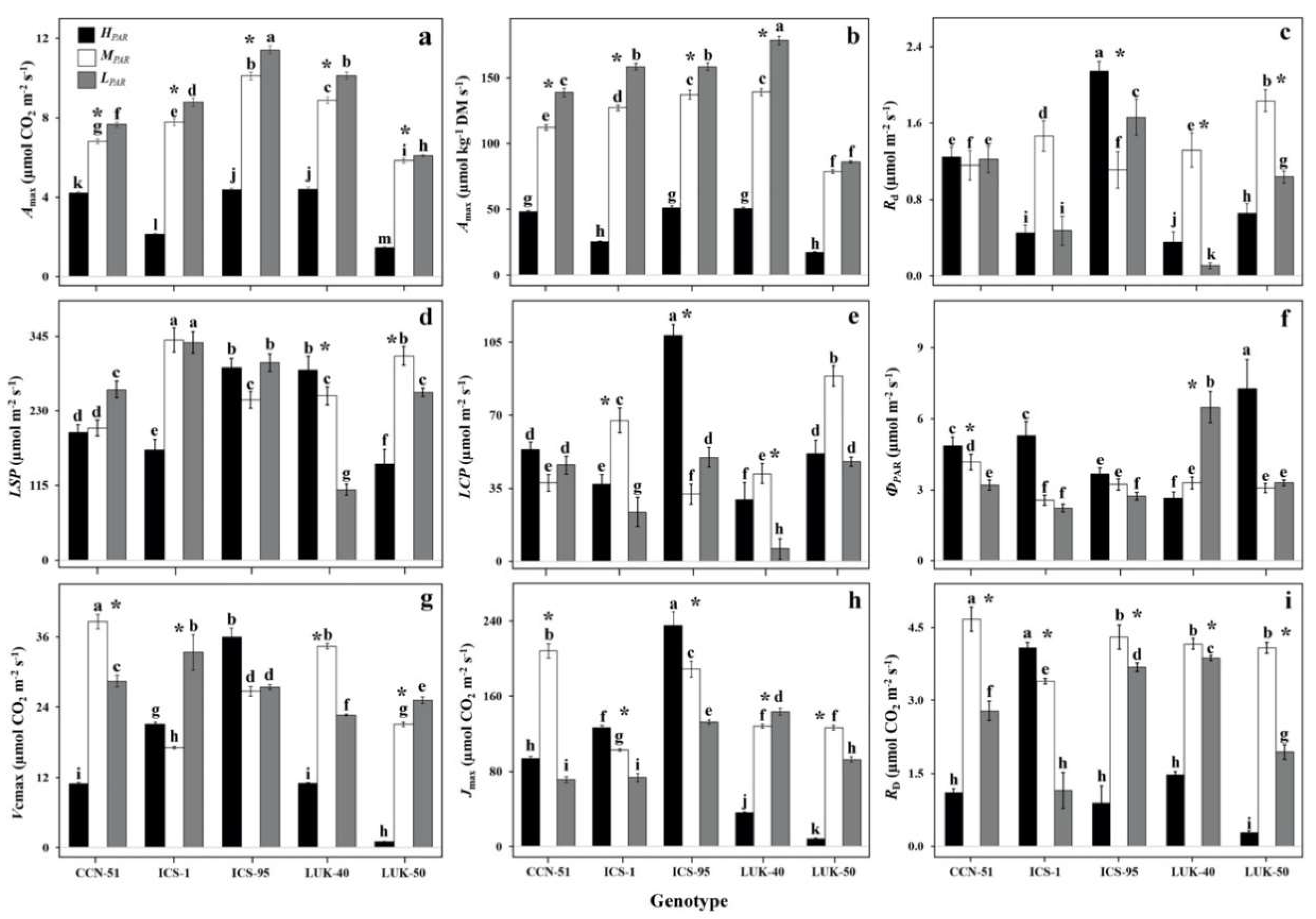
3.2. Photosynthetic Light- and CO2-Response Curves of Different Genotypes of Cacao Grown under Varying Levels of Radiation
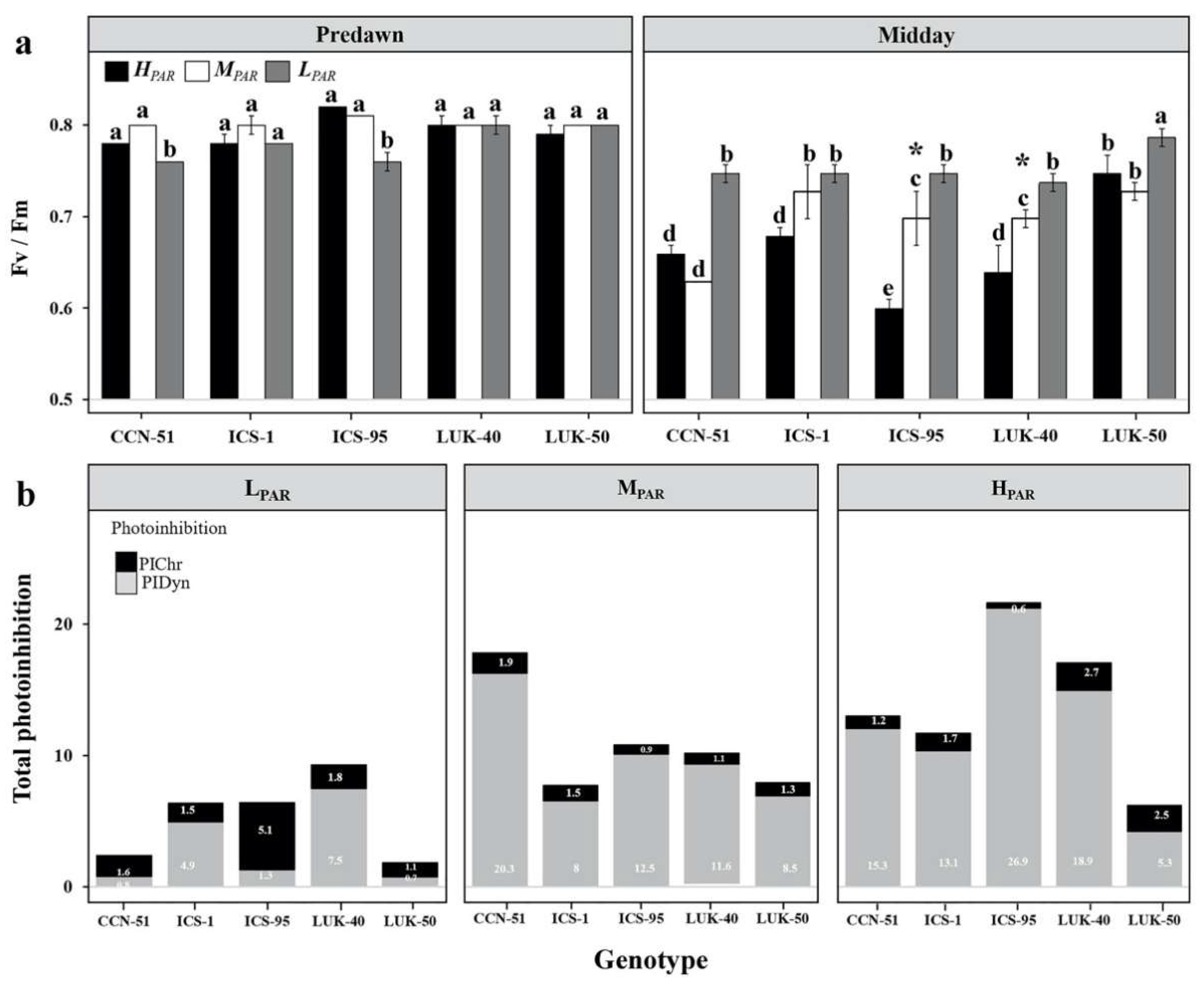
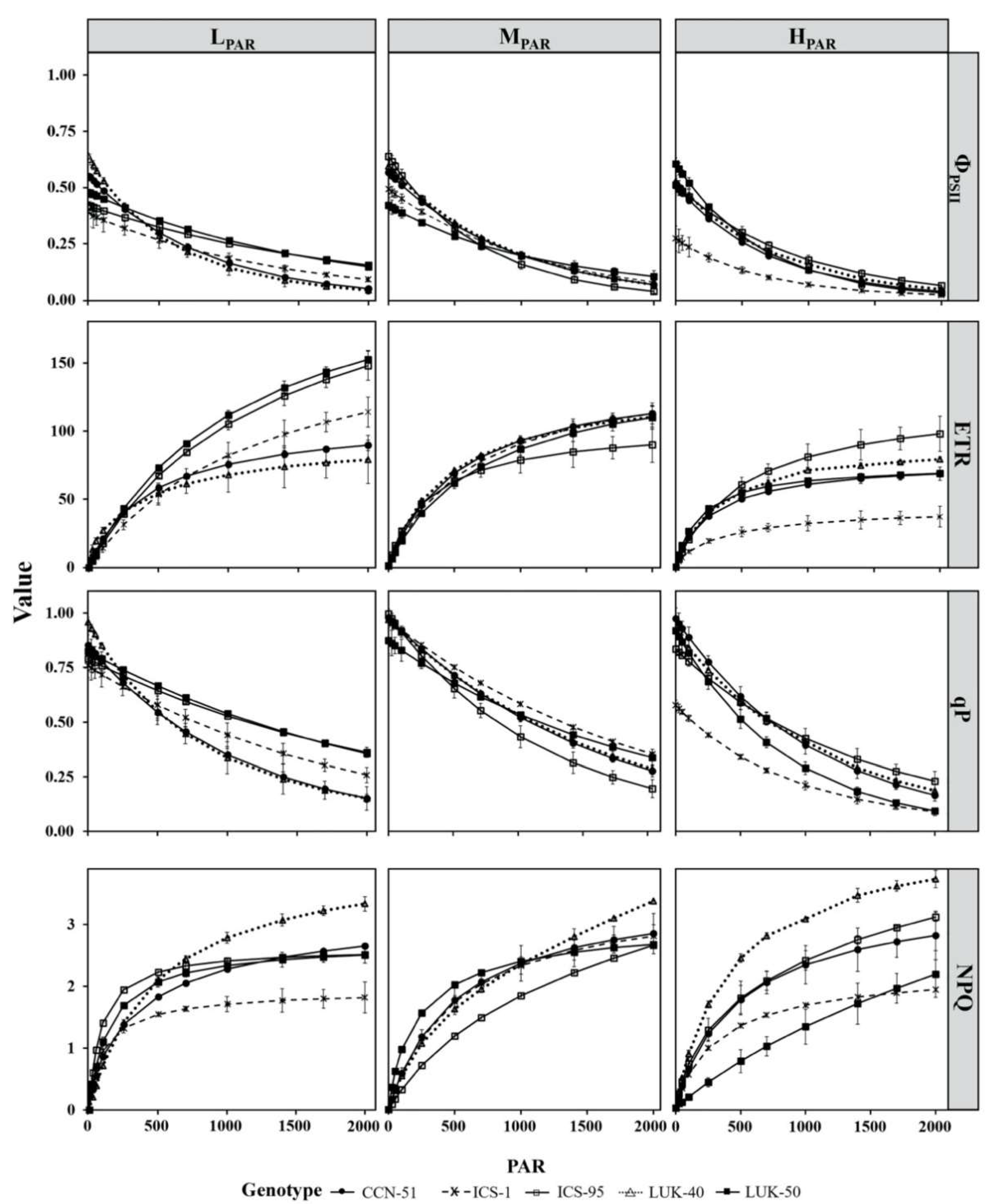
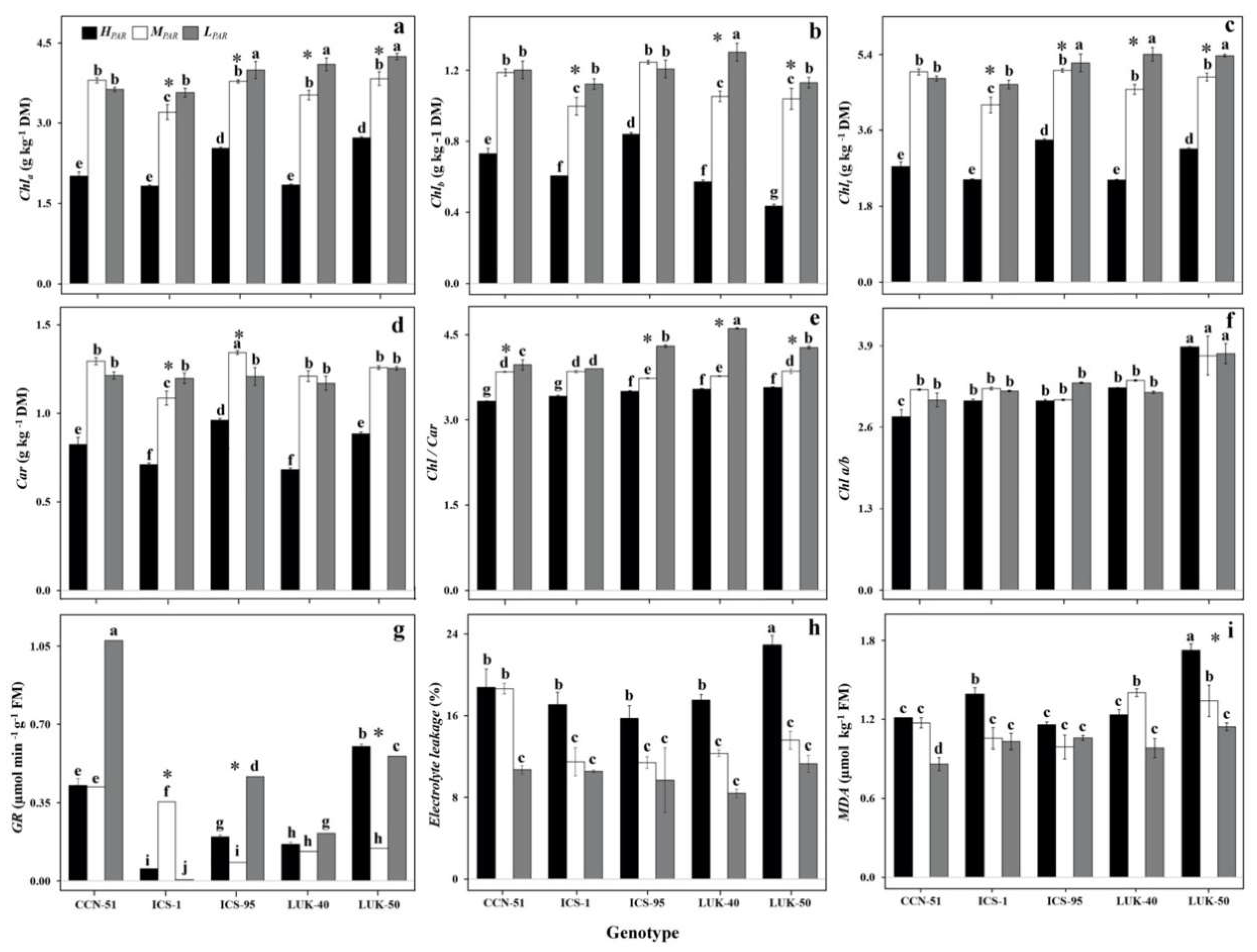
3.3. Chlorophyll (Chla) Parameters of Different Genotypes of Cacao Grown under Varying Levels of Radiation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Morales, A.; Kaiser, E. Photosynthetic Acclimation to Fluctuating Irradiance in Plants. Front. Plant Sci. 2020, 11, 268. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, F. Ecophysiological constraints on the production of shaded and unshaded coffee: A review. Field Crops Res. 2004, 86, 99–114. [Google Scholar] [CrossRef]
- Piato, K.; Lefort, F.; Subía, C.; Caicedo, C.; Calderón, D.; Pico, J.; Norgrove, L. Effects of shade trees on robusta coffee growth, yield and quality. A meta-analysis. Agron. Sustain. Dev. 2020, 40. [Google Scholar] [CrossRef]
- Lennon, A.M.; Lewis, V.R.; Farrell, A.D.; Umaharan, P. Photochemical responses to light in sun and shade leaves of Theobroma cacao L. (West African Amelonado). Sci. Hortic. 2021, 276, 109747. [Google Scholar] [CrossRef]
- Lahive, F.; Hadley, P.; Daymond, A.J. The physiological responses of cacao to the environment and the implications for climate change resilience. A review. Agron. Sustain. Dev. 2019, 39, 5. [Google Scholar] [CrossRef] [Green Version]
- Suárez, J.C.; Melgarejo, L.M.; Casanoves, F.; Di Rienzo, J.A.; DaMatta, F.; Armas, C. Photosynthesis limitations in cacao leaves under different agroforestry systems in the Colombian Amazon. PLoS ONE 2018, 13, e0206149. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Branco, M.C.; de Almeida, A.A.F.; Dalmolin, Â.C.; Ahnert, D.; Baligar, V.C. Influence of low light intensity and soil flooding on cacao physiology. Sci. Hortic. 2017, 217, 243–257. [Google Scholar] [CrossRef]
- De Araújo, R.P.; de Almeida, A.A.F.; Barroso, J.P.; de Oliveira, R.A.; Gomes, F.P.; Ahnert, D.; Baligar, V. Molecular and morphophysiological responses cocoa leaves with different concentrations of anthocyanin to variations in light levels. Sci. Hortic. 2017, 224, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008; pp. 1–604. [Google Scholar]
- Chaves, A.R.M.; Ten-Caten, A.; Pinheiro, H.A.; Ribeiro, A.; DaMatta, F.M. Seasonal changes in photoprotective mechanisms of leaves from shaded and unshaded field-grown coffee (Coffea arabica L.) trees. Trees 2007, 22, 351–361. [Google Scholar] [CrossRef]
- Werner, C.; Correia, O.; Beyschlag, W. Characteristic patterns of chronic and dynamic photoinhibition of different functional groups in a Mediterranean ecosystem. Funct. Plant Biol. 2002, 29, 999. [Google Scholar] [CrossRef]
- He, J.; Lim, R.M.P.; Dass, S.H.J.; Yam, T.W. Photosynthetic acclimation of Grammatophyllum speciosum to growth irradiance under natural conditions in Singapore. Bot. Stud. 2017, 58, 58. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Martins, S.C.V.; Araújo, W.L.; Tohge, T.; Fernie, A.R.; DaMatta, F.M. In High-Light-Acclimated Coffee Plants the Metabolic Machinery Is Adjusted to Avoid Oxidative Stress Rather than to Benefit from Extra Light Enhancement in Photosynthetic Yield. PLoS ONE 2014, 9, e94862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Q.; Wang, H.; Guo, H.; Zhou, A.; Huang, Y.; Sun, Y.; Li, M. Effects of Shade Treatments on Photosynthetic Characteristics, Chloroplast Ultrastructure, and Physiology of Anoectochilus roxburghii. PLoS ONE 2014, 9, e85996. [Google Scholar] [CrossRef]
- Wang, L.-F. Physiological and Molecular Responses to Variation of Light Intensity in Rubber Tree (Hevea brasiliensis Muell. Arg.). PLoS ONE 2014, 9, e89514. [Google Scholar] [CrossRef]
- Sharma, S.; Kataria, S.; Joshi, J.; Guruprasad, K.N. Antioxidant defense response of fenugreek to solar UV. Int. J. Veg. Sci. 2018, 25, 40–57. [Google Scholar] [CrossRef]
- Motamayor, J.C.; Lachenaud, P.; da Silva e Mota, J.W.; Loor, R.; Kuhn, D.N.; Brown, J.S.; Schnell, R.J. Geographic and Genetic Population Differentiation of the Amazonian Chocolate Tree (Theobroma cacao L). PLoS ONE 2008, 3, e3311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ICCO. ICCO Quarterly Bulletin of Cocoa Statistics. The International Cocoa Organization (ICCO) Cocoa Producing and Cocoa Consuming Countries. 2020. Available online: https://www.icco.org/ (accessed on 7 December 2020).
- Ávila-Lovera, E.; Coronel, I.; Jaimez, R.E.; Urich, R.; Pereyra, G.; Araque, O.; Chacón, I.; Tezara, W. Ecophysiological traits of adult trees of Criollo cocoa cultivars (Theobroma cacao L.) from a germplasm bank in Venezuela. Exp. Agric. 2015, 52, 137–153. [Google Scholar] [CrossRef]
- Almeida, A.-A.F.; Gomes, F.P.; Araujo, R.P.; Santos, R.C.; Valle, R.R. Leaf gas exchange in species of the Theobroma genus. Photosynthetica 2014, 52, 16–21. [Google Scholar] [CrossRef]
- Araque, O.; Jaimez, R.E.; Tezara, W.; Coronel, I.; Urich, R.; Espinoza, W.; Araque, O.; Jaimez, R.E.; Tezara, W.; Coronel, I.; et al. Comparative photosynthesis, water relations, growth and survival rates in juvenile Criollo cacao cultivars (Theobroma cacao) during dry and wet seasons. Exp. Agric. 2012, 48, 513–522. [Google Scholar] [CrossRef]
- Baligar, V.C.; Bunce, J.A.; Machado, R.C.R.; Elson, M.K. Photosynthetic photon flux density, carbon dioxide concentration, and vapor pressure deficit effects on photosynthesis in cacao seedlings. Photosynthetica 2008, 46, 216–221. [Google Scholar] [CrossRef]
- Balasimha, D.; Daniel, E.; Bhat, P.G. Influence of environmental factors on photosynthesis in cocoa trees. Agric. For. Meteorol. 1991, 55, 15–21. [Google Scholar] [CrossRef]
- Daymond, A.J.; Tricker, P.; Hadley, P. Genotypic variation in photosynthesis in cacao is correlated with stomatal conductance and leaf nitrogen. Biol. Plant. 2011, 55, 99–104. [Google Scholar] [CrossRef]
- Miyaji, K.-I.; Da Silva, W.S.; Alvim, P.D.T. Longevity of leaves of a tropical tree, Theobroma cacao, grown under shading, in relation to position within the canopy and time of emergence. New Phytol. 1997, 135, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Galyuon, I.K.A.; McDavid, C.R.; Lopez, F.B.; Spence, J.A. The effect of irradiance level on cocoa (Theobroma cacao L.): II. Gas exchange and chlorophyll fluorescence. Trop. Agric. 1996, 73, 29–33. [Google Scholar]
- Raja, H.; Hardwick, K. The effects of prolonged exposure to different light intensities on the photosynthesis of cocoa leaves. In Proceedings of the 10th International Cocoa Research Conference, Santo Domingo, Dominican Republic, 17–23 May 1987; Cocoa Producers’ Alliance: Lagos, Nigeria, 1988; pp. 205–209. [Google Scholar]
- Jaimez, R.E.; Puyutaxi, F.A.; Vasco, A.; Loor, R.G.; Tarqui, O.; Quijano, G.; Jimenez, J.C.; Tezara, W. Photosynthetic response to low and high light of cacao growing without shade in an area of low evaporative demand. Acta Biol. Colomb. 2018, 23, 95–103. [Google Scholar] [CrossRef]
- Rajab, Y.A.; Leuschner, C.; Barus, H.; Tjoa, A.; Hertel, D. Cacao Cultivation under Diverse Shade Tree Cover Allows High Carbon Storage and Sequestration without Yield Losses. PLoS ONE 2016, 11, e0149949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedel, J.; Kägi, N.; Armengot, L.; Schneider, M. Effects of rehabilitation pruning and agroforestry on cacao tree development and yield in an older full-sun plantation. Exp. Agric. 2019, 55, 849–865. [Google Scholar] [CrossRef]
- Bentley, J.W.; Boa, E.; Stonehouse, J. Neighbor Trees: Shade, Intercropping, and Cacao in Ecuador. Hum. Ecol. 2004, 32, 241–270. [Google Scholar] [CrossRef]
- IDEAM. Atlas Climatológico de Colombia. 2020. Available online: http://www.ideam.gov.co (accessed on 15 February 2019).
- Salazar, J.C.S.; Bieng, M.A.N.; Melgarejo, L.M.; Di Rienzo, J.A.; Casanoves, F. First typology of cacao (Theobroma cacao L.) systems in Colombian Amazonia, based on tree species richness, canopy structure and light availability. PLoS ONE 2018, 13, e0191003. [Google Scholar] [CrossRef] [Green Version]
- Niether, W.; Armengot, L.; Andres, C.; Schneider, M.; Gerold, G. Shade trees and tree pruning alter throughfall and microclimate in cocoa (Theobroma cacao L.) production systems. Ann. For. Sci. 2018, 75, 38. [Google Scholar] [CrossRef] [Green Version]
- Acheampong, K.; Hadley, P.; Daymond, A.J. Photosynthetic activity and early growth of four cacao genotypes as influenced by different shade regimes under west african dry and wet season conditions. Exp. Agric. 2013, 49, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Rada, F.; Jaimez, R.E.; Garcia, C.; Azocar, A.; Ramirez, M.E. Water relations and gas exchange in Theobroma cacao var. Guasare under periods of water deficit. J. Biol. Chem. 1995, 270, 26723–26726. [Google Scholar]
- Da Matta, F.M.; Loos, R.A.; Rodrigues, R.; Barros, R.S. Actual and potential photosynthetic rates of tropical crop species. Rev. Bras. Fisiol. Veg. 2001, 13, 24–32. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Salazar, J.C.S.; Polanía, J.A.; Bastidas, A.T.C.; Suárez, L.R.; Beebe, S.; Rao, I.M. Agronomical, phenological and physiological performance of common bean lines in the Amazon region of Colombia. Theor. Exp. Plant Physiol. 2018, 30, 303–320. [Google Scholar] [CrossRef]
- Bauerle, W.L.; Wang, G.G.; Bowden, J.D.; Hong, C.M. An analysis of ecophysiological responses to drought in American Chestnut. Ann. For. Sci. 2006, 63, 833–842. [Google Scholar] [CrossRef]
- Long, S.P. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef] [Green Version]
- Martins, S.C.V.; Detmann, K.C.; Dos Reis, J.V.; Pereira, L.F.; Sanglard, L.M.V.P.; Rogalski, M.; DaMatta, F.M. Photosynthetic induction and activity of enzymes related to carbon metabolism: Insights into the varying net photosynthesis rates of coffee sun and shade leaves. Theor. Exp. Plant Physiol. 2013, 25, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Diaz-Espejo, A.; Berry, J.A.; Cifre, J.; Galmés, J.; Kaldenhoff, R.; Medrano, H.; Ribas-Carbo, M. Analysis of leakage in IRGA’s leaf chambers of open gas exchange systems: Quantification and its effects in photosynthesis parameterization. J. Exp. Bot. 2007, 58, 1533–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta (BBA) Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Brooks, M.D.; Niyogi, K.K. Use of a Pulse-Amplitude Modulated Chlorophyll Fluorometer to Study the Efficiency of Photosynthesis in Arabidopsis Plants. In Chloroplast Research in Arabidopsis; Jarvis, R.P., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2011; Volume 775, pp. 299–310. ISBN 978-1-61779-236-6. [Google Scholar]
- Silva Neto, L.; de França da Silva, I.; Inda, A.V.; do Nascimento, P.C.; Bortolon, L. Atributos físicos e químicos de agregados pedogênicos e de coprólitos de minhocas em diferentes classes de solos da paraíba. Cienc Agrotecnol. 2010, 34, 1365–1371. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Pinheiro, H.; DaMatta, F.M.; Chaves, A.R.; Fontes, E.; Loureiro, M.E. Drought tolerance in relation to protection against oxidative stress in clones of Coffea canephora subjected to long-term drought. Plant Sci. 2004, 167, 1307–1314. [Google Scholar] [CrossRef]
- Lima, A.L.S.; DaMatta, F.M.; Pinheiro, H.; Tótola, M.; Loureiro, M.E. Photochemical responses and oxidative stress in two clones of Coffea canephora under water deficit conditions. Environ. Exp. Bot. 2002, 47, 239–247. [Google Scholar] [CrossRef]
- De Lobo, F.A.; de Barros, M.P.; Dalmagro, H.J.; Dalmolin, A.; Pereira, W.E.; de Souza, É.C.; Vourlitis, G.L.; Ortíz, C.E.R. Fitting net photosynthetic light-response curves with Microsoft Excel—A critical look at the models. Photosynthetica 2013, 51, 445–456. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Duursma, R.A. Plantecophys—An R Package for Analysing and Modelling Leaf Gas Exchange Data. PLoS ONE 2015, 10, e0143346. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. Linear and nonlinear mixed effects models. R Package Version 2019, 3, 111. [Google Scholar]
- R Development Core Team. R: The R Project for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2019; ISBN 3-900051-07-0. Available online: https://www.r-project.org/ (accessed on 25 June 2019).
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Di, R.C.W. InfoStat; Grupo InfoStat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2019. [Google Scholar]
- Ayegboyin, K.O.; Akinrinde, E.A. Effect of Water Deficit Imposed during the Early Developmental Phase on Photosynthesis of Cocoa (Theobroma cacao L.). Agric. Sci. 2016, 7, 11–19. [Google Scholar]
- Lahive, F.; Hadley, P.; Daymond, A.J. The impact of elevated CO2 and water deficit stress on growth and photosynthesis of juvenile cacao (Theobroma cacao L.). Photosynthetica 2017, 56, 911–920. [Google Scholar] [CrossRef] [Green Version]
- Baligar, V.C.; Elson, M.K.; Almeida, A.-A.F.; de Araujo, Q.R.; Ahnert, D.; He, Z. Carbon Dioxide Concentrations and Light Levels on Growth and Mineral Nutrition of Juvenile Cacao Genotypes. Am. J. Plant Sci. 2021, 12, 818–839. [Google Scholar] [CrossRef]
- Baligar, V.C.; Bunce, J.A.; Bailey, B.A.; Machado, R.C.; Pomella, A.W. Carbon Dioxide and Photosynthetic Photon Flux Density Effects on Growth and Mineral Uptake of Cacao. Food Agr. Environ. 2015, 3, 142–147. [Google Scholar]
- Saavedra, F.; Peña, E.J.; Schneider, M.; Naoki, K. Effects of environmental variables and foliar traits on the transpiration rate of cocoa (Theobroma cacao L.) under different cultivation systems. Agrofor. Syst. 2020, 94, 2021–2031. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez, J.C.; Casanoves, F.; Bieng, M.A.N.; Melgarejo, L.M.; Di Rienzo, J.A.; Armas, C. Prediction model for sap flow in cacao trees under different radiation intensities in the western Colombian Amazon. Sci. Rep. 2021, 11, 10512. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, D.; Moreno, F.; Díez, M.C. Photosynthesis, growth, and survival in seedlings of four tropical fruit-tree species under intense radiation. Acta Amaz. 2021, 51, 1–9. [Google Scholar] [CrossRef]
- De Almeida, J.; Herrera, A.; Tezara, W. Phenotypic plasticity to photon flux density of physiological, anatomical and growth traits in a modern Criollo cocoa clone. Physiol. Plant. 2018, 166, 821–832. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suárez, J.C.; Gelpud, C.; Noriega, J.E.; Ortiz-Morea, F.A. How Do Different Cocoa Genotypes Deal with Increased Radiation? An Analysis of Water Relation, Diffusive and Biochemical Components at the Leaf Level. Agronomy 2021, 11, 1422. https://doi.org/10.3390/agronomy11071422
Suárez JC, Gelpud C, Noriega JE, Ortiz-Morea FA. How Do Different Cocoa Genotypes Deal with Increased Radiation? An Analysis of Water Relation, Diffusive and Biochemical Components at the Leaf Level. Agronomy. 2021; 11(7):1422. https://doi.org/10.3390/agronomy11071422
Chicago/Turabian StyleSuárez, Juan Carlos, Cristian Gelpud, Jhon Eduar Noriega, and Fausto Andrés Ortiz-Morea. 2021. "How Do Different Cocoa Genotypes Deal with Increased Radiation? An Analysis of Water Relation, Diffusive and Biochemical Components at the Leaf Level" Agronomy 11, no. 7: 1422. https://doi.org/10.3390/agronomy11071422
APA StyleSuárez, J. C., Gelpud, C., Noriega, J. E., & Ortiz-Morea, F. A. (2021). How Do Different Cocoa Genotypes Deal with Increased Radiation? An Analysis of Water Relation, Diffusive and Biochemical Components at the Leaf Level. Agronomy, 11(7), 1422. https://doi.org/10.3390/agronomy11071422