
A Niche for Cowpea in Sub-Tropical Australia?

 , , and
, , and
Abstract
:1. Introduction
2. History of Cowpea Research in Australia
3. Genetic Resources & Breeding Technologies
4. Predicting the Agronomic Fit of Cowpea
4.1. Simulation Modelling Approach & Assumptions
- Photosynthetic response to temperature was adjusted to reflect the higher temperature tolerance [21] and higher water use-efficiency of cowpea under water stress [20]. This was implemented by increasing by 4 °C the maximum thresholds at which the radiation use efficiency (RUE) is reduced, increasing the maximum temperature for root advance from 32 to 38 °C, and increasing the maximum temperatures that induce leaf senescence from 34 to 35 °C.
- Cowpea have been shown to exhibit much lower leaf senescence and abscission and, hence, to maintain leaf area for longer under moisture-stressed rainfed conditions than mungbean [22]. To capture this difference, the leaf senescence rate during water stress for cowpea was also reduced relative to mungbean (from 0.05 to 0.02), to represent the capacity of cowpea to maintain leaf area through drought periods.
- Root shoot ratio was increased by 30% during all growth stages, and the root growth rate was increased to reflect a greater allocation to roots in cowpea than mungbean [20].
- The phenological development was adjusted to allow for a longer period of grain filling from 280 to 360 degree-days for cv. Banjo and from 362 to 450 degree-days for Red Caloona. This was based on previous field observation of their relative phenology (Andrew James, unpublished data).
- Initial leaf dry matter in the cowpea model was increased from 0.03 to 0.1, and initial leaf area index (LAI) was reduced from 800 to 200 mm2/plant in order to be consistent with mungbean.
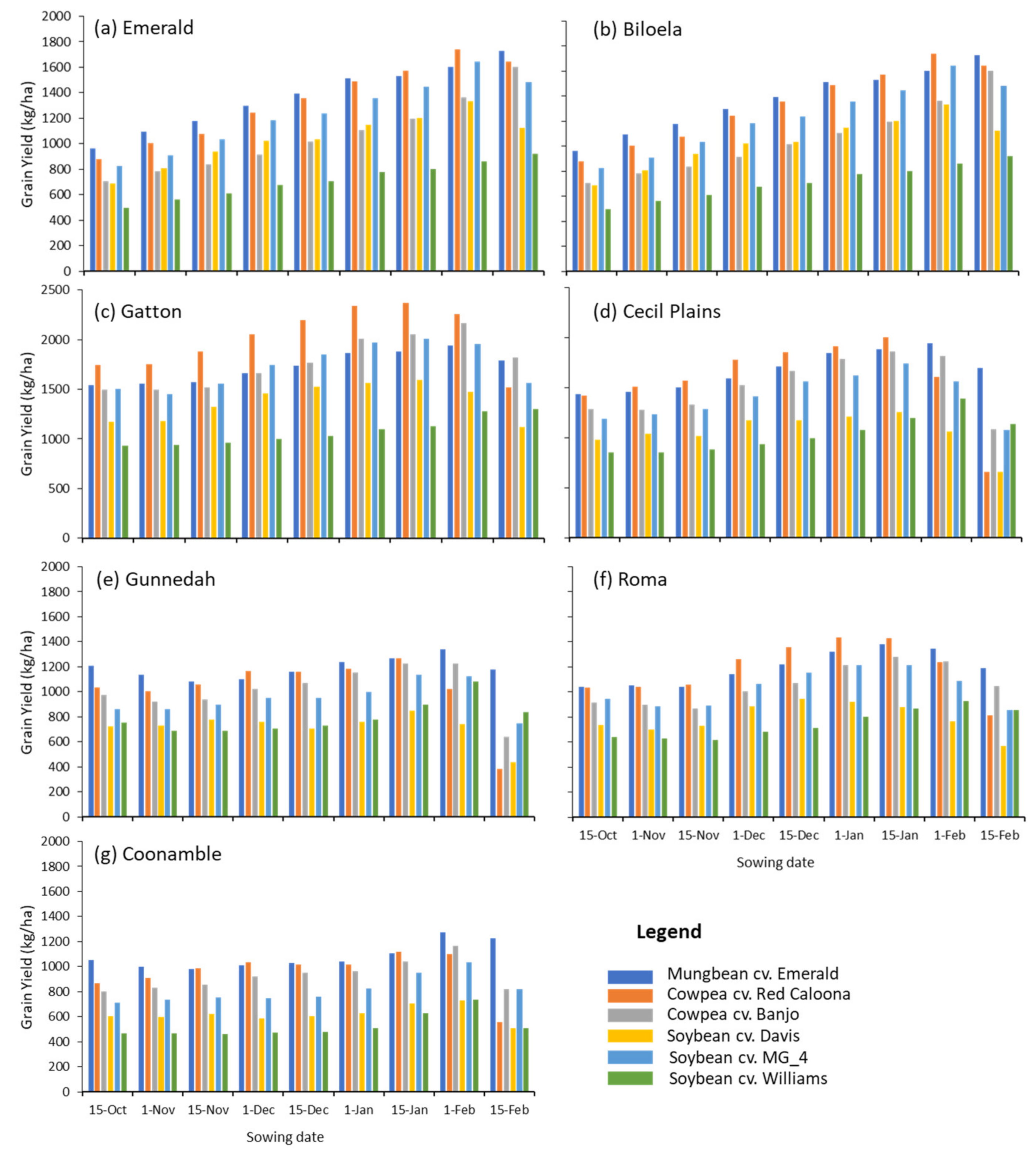
4.2. Predicted Relative Productivity and Fit of Cowpea
4.3. Further Considerations and Analysis Needs
5. Cowpea Potential for the Food Industry
6. Cowpea in Aquafeeds
7. Conclusions and Commercial Opportunities
- There is abundant germplasm that could be used in a revived cowpea breeding program. This, together with the new genetic resources that are available for cowpea, will speed-up breeding efforts in this crop.
- Our modelling work indicates that cowpea will be a competitive crop in the targeted areas. Across all locations, cowpea genotypes were simulated to achieve similar average grain yields to those in mungbean, and, in some locations and situations, average grain yields of cowpea were predicted to be larger.
- Regarding uses of cowpea as a food ingredient, we have found that cowpea protein has the potential to be part of the growing market for global protein ingredients and that it can have different functionalities to other food legumes for making differentiated concentrates and isolates.
- We have identified potential application for cowpea and protein enriched products in aquafeed formulations, due to the distinctive composition of the grain. The development of formulation strategies including cowpeas and their protein concentrates could add extra value to this crop.
- Cowpea can offer genetically modified approaches, for example for insect-resistance, which could differentiate cowpea from other similar beans.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rachaputi, R.C.; Sands, D.; McKenzie, K.; Agius, P.; Lehane, J.; Seyoum, S. Eco-physiological drivers influencing mungbean [Vigna radiata (L.) Wilczek] productivity in subtropical Australia. Field Crop. Res. 2019, 238, 74–81. [Google Scholar] [CrossRef]
- Vanambathina, P.; Rachaputi, R.C.N.; Henry, R.J.; Norton, S.L. Phenotypic variation in Australian wild Cajanus and their interspecific hybrids. Genet. Resour. Crop. Evol. 2019, 66, 1699–1712. [Google Scholar] [CrossRef]
- Earth Systems and Climiate Change Hub (2015–2021). Climate Change in a Land of Extremes. Available online: https://nespclimate.com.au/wp-content/uploads/2021/06/NESP-Climate-Change-Hub-Climate-Change-in-a-Land-of-Extremes-Report-Web_med-res-1.pdf (accessed on 1 August 2021).
- Ehlers, J.D.; Hall, A.E. Cowpea (Vigna unguiculata L. Walp.). Field Crop. Res. 1997, 53, 187–204. [Google Scholar] [CrossRef]
- Holzworth, D.P.; Huth, N.I.; deVoil, P.G.; Zurcher, E.J.; Herrmann, N.I.; McLean, G.; Keating, B.A. APSIM—Evolution towards a new generation of agricultural systems simulation. Environ. Model. Softw. 2014, 62, 327–350. [Google Scholar] [CrossRef]
- Imrie, B.C. Register of Australian grain legume cultivars: Vigna radiata L. (cowpea) cv. Big Buff. Aust. J. Exp. Agric. 1995, 35, 678. [Google Scholar] [CrossRef]
- Imrie, B.C.; Gammie, R.L. Register of Australian grain legume cultivars: Vigna unguiculata L. (cowpea) cv. Holstein: Reg. no. ARGL 94-4. Aust. J. Exp. Agric. 1995, 35, 679. [Google Scholar] [CrossRef]
- Lawn, R.J. The Australian Vigna species: A case study in the collection and conservation of crop wild relatives. In Crop Wild Relatives and Climate Change; Redden, R.J., Yadav, S.S., Maxted, N., Dulloo, M.E., Guarino, L., Smith, P., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2015; pp. 318–335. [Google Scholar]
- Singh, B.B.; Ehlers, J.D.; Sharma, B.; Freire Filho, F.R. Recent progress in cowpea breeding. In Challenges and Opportunities for Enhancing Sustainable Cowpea Production; Fatokun, C.A., Tarawali, S.A., Singh, B.B., Kormawa, P.M., Tamo, M., Eds.; International Institute of Tropical Agriculture: Ibadan, Nigeria, 2002; pp. 22–40. [Google Scholar]
- Lonardi, S.; Muñoz-Amatriaín, M.; Liang, Q.; Shu, S.; Wanamaker, S.I.; Lo, S.; Close, T.J. The genome of cowpea (Vigna unguiculata [L.] Walp.). Plant J. 2019, 98, 767–782. [Google Scholar] [CrossRef] [Green Version]
- Yao, S.; Jiang, C.; Huang, Z.; Torres-Jerez, I.; Chang, J.; Zhang, H.; Verdier, J. The Vigna unguiculata Gene Expression Atlas (VuGEA) from de novo assembly and quantification of RNA-seq data provides insights into seed maturation mechanisms. Plant J. 2016, 88, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Huynh, B.L.; Ehlers, J.D.; Huang, B.E.; Muñoz-Amatriaín, M.; Lonardi, S.; Santos, J.R.; Roberts, P.A. A Multi-Parent Advanced Generation Inter-Cross (MAGIC) population for genetic analysis and improvement of cowpea (Vigna unguiculata L. Walp.). Plant J. 2018, 93, 1129–1142. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.R.P.; Ndeve, A.D.; Huynh, B.-L.; Matthews, W.C.; Roberts, P.A. QTL mapping and transcriptome analysis of cowpea reveals candidate genes for root-knot nematode resistance. PLoS ONE 2018, 13, e0189185. [Google Scholar]
- Lo, S.; Muñoz-Amatriaín, M.; Hokin, S.A.; Cisse, N.; Roberts, P.A.; Farmer, A.D.; Xu, S.; Close, T.J. A genome-wide association and meta-analysis reveal regions associated with seed size in cowpea [Vigna unguiculata (L.) Walp]. Appl. Genet. 2019, 132, 3079–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, D.P.; Sharma, S.P.; Lal, M.; Ranwah, B.R.; Sharma, V. Induction of Genetic variability for polygentraits through physical and chemical mutagens in cowpea (Vigna unguiculata). Legume Res. 2013, 36, 10–14. [Google Scholar]
- Raina, A.; Laskar, R.A.; Tantray, Y.R.; Khursheed, S.; Wani, M.R.; Khan, S. Characterization of Induced High Yielding Cowpea Mutant Lines Using Physiological, Biochemical and Molecular Markers. Sci. Rep. 2020, 10, 3687. [Google Scholar] [CrossRef] [Green Version]
- Bett, B.; Gollasch, S.; Moore, A.; Harding, R.; Higgins, T.J.V. An Improved Transformation System for Cowpea (Vigna unguiculata L. Walp) via Sonication and a Kanamycin-Geneticin Selection Regime. Front. Plant Sci. 2019, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Barrero, J.M.; Higgins, T.J.V. Nigeria Has Given a New GM Cowpea Variety the Go Ahead. Why It Matters. Conversation. 2020. Available online: https://theconversation.com/nigeria-has-given-a-new-gm-cowpea-variety-the-go-ahead-why-it-matters-130304 (accessed on 1 August 2021).
- Popelka, J.C.; Gollasch, S.; Moore, A.; Molvig, L.; Higgins, T.J. Genetic transformation of cowpea (Vigna unguiculata L.) and stable transmission of the transgenes to progeny. Plant. Cell Rep. 2006, 25, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Lawn, R.J. Response of four grain legumes to water stress in south-eastern Queensland. III. Dry matter production, yield and water-use-efficiency. Aust. J. Agric. Res. 1982, 33, 511–521. [Google Scholar] [CrossRef]
- Lawn, R.J. Response of four grain legumes to water stress in south-eastern Queensland. I. Physiological response mechanisms. Aust. J. Agric. Res. 1982, 33, 481–496. [Google Scholar] [CrossRef]
- Lawn, R.J. Response of four grain legumes to water stress in south-eastern Queensland. II. Plant growth and soil water extraction patterns. Aust. J. Agric. Res. 1982, 33, 497–509. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Muchow, R.C.; Ludlow, M.M.; Leach, G.J.; Lawn, R.J.; Foale, M.A. Field and model analysis of the effect of water deficits on carbon and nitrogen accumulation by soybean, cowpea and black gram. Field Crop. Res. 1987, 17, 121–140. [Google Scholar] [CrossRef]
- Hall, C.; Hillen, C.; Robinson, J.G. Composition, Nutritional Value, and Health Benefits of Pulses. Cereal. Chem. 2017, 94, 11–31. [Google Scholar] [CrossRef]
- Jayathilake, C.; Visvanathan, R.; Deen, A.; Bangamuwage, R.; Jayawardana, B.C.; Nammi, S.; Liyanage, R. Cowpea: An overview on its nutritional facts and health benefits. J. Sci. Food Agric. 2018, 98, 4793–4806. [Google Scholar] [CrossRef]
- Farinu, G.O.; Ingrao, G. Gross composition, amino acid, phytic acid and trace element contents of thirteen cowpea cultivars and their nutritional significance. J. Sci. Food Agric. 1991, 55, 401–410. [Google Scholar] [CrossRef]
- Ogunmodede, B.K.; Oyenuga, V.A. Vitamin B content of cowpeas (Vigna unguiculata Walp). II. Pyridoxine, pantothenic acid, biotin and folic acid. J. Sci. Food Agric. 1970, 21, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Hoppner, K.; Lampi, B. Folate retention in dried legumes after different methods of meal preparation. Food Res. Int. 1993, 26, 45–48. [Google Scholar] [CrossRef]
- Gonçalves, A.; Goufo, P.; Barros, A.; Domínguez-Perles, R.; Trindade, H.; Rosa, E.A.; Ferreira, L.; Rodrigues, M. Cowpea (Vigna unguiculata L. Walp), a renewed multipurpose crop for a more sustainable agri-food system: Nutritional advantages and constraints. J. Sci. Food Agric. 2016, 96, 2941–2951. [Google Scholar] [CrossRef]
- Plahar, W.A.; Annan, N.T.; Nti, C.A. Cultivar and processing effects on the pasting characteristics, tannin content and protein quality and digestibility of cowpea (Vigna unguiculata). Plant. Foods Hum. Nutr. 1997, 51, 343–356. [Google Scholar] [CrossRef]
- Chang, M.C.; Bailey, J.W.; Collins, J.L. Dietary tannins from cowpeas and tea transiently alter apparent calcium absorption but not absorption and utilization of protein in rats. J. Nutr. 1994, 124, 283–288. [Google Scholar] [CrossRef]
- OECD Consensus Document on Compositional Considerations for New Varieties of Cowpea (Vigna Un-Guiculata). Series on the Safety of Novel Foods and Feeds No. 30. Environment Directorate. Paris. 2018. Available online: http://www.oecd.org/officialdocuments/publicdisplaydocumentpdf/?cote=env%2Fjm%2Fmono(2018)36&doclanguage=en (accessed on 10 February 2021).
- Sipeniece, E.; Mišina, I.; Qian, Y.; Grygier, A.; Sobieszczańska, N.; Sahu, P.K.; Rudzińska, M.; Patel, K.S.; Górnaś, P. Fatty Acid Profile and Squalene, Tocopherol, Carotenoid, Sterol Content of Seven Selected Consumed Legumes. Plant. Foods Hum. Nutr. 2021, 76, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Aguilera, J.M. A review of the textural defects in cooked reconstituted llegumes—The influence of structure and composition. J. Food Biochem. 1985, 9, 277–323. [Google Scholar] [CrossRef]
- Petterson, D.S.; Mackintosh, J.B. The Chemical Composition and Nutritive Value of Australian Legume Grains; Grains Research and Development Corporation: Canberra, Australia, 1994. [Google Scholar]
- Phillips, R.; McWatters, K.H.; Chinnan, M.S.; Hung, Y.-C.; Beuchat, L.R.; Sefa-Dedeh, S.; Sakyi-Dawson, E.; Ngoddy, P.; Nnanyelugo, D.; Enwere, J.; et al. Utilization of cowpeas for human food. Field Crop. Res. 2003, 82, 193–213. [Google Scholar] [CrossRef]
- Prinyawiwatkul, W.; McWatters, K.H.; Beuchat, L.R.; Phillips, R.D.; Uebersak, M.A. Cowpea flour: A potential ingredient in food products. Crit. Rev. Food Sci. Nutr. 1996, 36, 413–436. [Google Scholar] [CrossRef]
- Shevkani, K.; Singh, N.; Chen, Y.; Kaur, A.; Yu, L. Pulse proteins: Secondary structure, functionality and applications. J. Food Sci. Technol. 2019, 56, 2787–2798. [Google Scholar] [CrossRef] [PubMed]
- Peyrano, F.; Speroni, F.; Avanza, M.V. Physicochemical and functional properties of cowpea protein isolates treated with temper-ature or high hydrostatic pressure. Innov. Food Sci. Emerg. Technol. 2016, 33, 38–46. [Google Scholar] [CrossRef]
- Technavio 2017. Global Meat Substitutes Markets 2017–2022. Available online: https://www.technavio.com/report/meat-substitutes-market-industry-analysis (accessed on 15 May 2021).
- Rangel, A.; Saraiva, K.; Schwengber, P.; Narciso, M.S.; Domont, G.B.; Ferreira, S.T.; Pedrosa, C. Biological evaluation of a protein isolate from cowpea (Vigna unguiculata) seeds. Food Chem. 2004, 87, 491–499. [Google Scholar] [CrossRef]
- Frota, K.; dos Santos, R.D.; Ribeiro, V.Q.; Arêas JA, G. Cowpea protein reduces LDL-cholesterol and apolipoprotein B concentrations but does not improve biomarkers of inflammation or endothelial dysfunction in adults with moderate hy-percholesterolemia. Nutr. Hosp. 2015, 31, 1611–1619. [Google Scholar]
- Mune, M.M.; Saxena, D.; Minka, S. Antioxidant activity of cowpea protein isolate hydrolyzed by pepsin. Acta Aliment. 2014, 43, 614–622. [Google Scholar] [CrossRef] [Green Version]
- Thumbrain, D.; Dwarka, D.; Gerrano, A.S.; Mellem, J.J. Antioxidant and apoptotic potential of protein isolates derived from Vigna unguiculata (L.) Walp. Int. J. Food Sci. Technol. 2020, 55, 2813–2823. [Google Scholar] [CrossRef]
- Agriculture and Nutrition Opportunity Engine Series—Opportunities in the Global Protein Ingredients Market, Forecast to 2022. 2018. Available online: https://www.researchandmarkets.com/reports/4495384/agriculture-and-nutrition-opportunity-engine (accessed on 15 May 2021).
- Rombenso, A.N.; Esmaeili, M.; Araujo, B.; Emerenciano, M.; Truong, H.; Viana, M.T.; Li, E.; Simon, C. Macronutrient Research in Aquaculture Nutrition. Global Advocate Aquaculture Magazine. 2021. Available online: https://www.aquaculturealliance.org/advocate/macronutrient-research-in-aquaculture-nutrition/ (accessed on 15 May 2021).
- NRC (National Research Council). Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Garg, S.K.; Kalla, A.; Bhatnagar, A. Evaluation of raw and hydrothermically processed leguminous seeds as supplementary feed for the growth of two Indian major carp species. Aquac. Res. 2002, 33, 151–163. [Google Scholar] [CrossRef]
- Olivera-Castillo, L.; Pino-Aguilar, M.; Lara-Flores, M.; Granados-Puerto, S.; Montero-Muñoz, J.; Olvera-Novoa, M.; Grant, G. Substitution of fish meal with raw or treated cowpea (Vigna unguiculata L. Walp, IT86-D719) meal in diets for Nile tilapia (Oreochromis niloticus L.) fry. Aquac. Nutr. 2011, 17, e101–e111. [Google Scholar] [CrossRef]
- Eusebio, P.S. Effect of dehulling on the nutritive value of some leguminous seeds as protein sources for tiger prawn, Penaeus monodon, juveniles. Aquaculture 1991, 99, 297–308. [Google Scholar] [CrossRef]
- Millamena, O.M.; Trino, A.T. Low-cost feed for Penaeus monodon reared in tanks and under semi-intensive and intensive conditions in brackishwater ponds. Aquaculture 1997, 154, 69–78. [Google Scholar] [CrossRef]
- Rivas-Vega, M.E.; Goytortúa-Bores, E.; Ezquerra-Brauer, J.; Salazar-García, M.; Cruz-Suarez, L.E.; Nolasco, H.; Civera-Cerecedo, R. Nutritional value of cowpea (Vigna unguiculata L. Walp) meals as ingredients in diets for Pacific white shrimp (Litopenaeus vannamei Boone). Food Chem. 2006, 97, 41–49. [Google Scholar] [CrossRef]
- Aya, F.A.; Cuvin-Aralar, M.L.; Coloso, R.M. Potential of cowpea (Vigna unguiculata L.) meal as an alternative protein source in diets for giant freshwater prawn (Macrobrachium rosenbergii, de Man 1879). In Proceedings of the International Workshop on Resource Enhancement and Sustainable Aquaculture Practices in Southeast Asia 2014 (RESA), Iloilo City, Philippines, 5–7 March 2014; pp. 231–237. [Google Scholar]
- Eusebio, P.S.; Coloso, R.M.; Mamauag, R.E.P. Apparent digestibility of selected ingredients in diets for juvenile grouper, Ephinephelus coioides (Hamilton). Aquac. Res. 2004, 35, 1261–1269. [Google Scholar] [CrossRef]
- Keembiyehetty, C.N.; de Silva, S.S. Performance of juvenile Oreochromis niloticus (L.) reared on diets containing cowpea, Vigna catiang, and black gram, Phaseolus mungo, seeds. Aquaculture 1993, 112, 207–215. [Google Scholar] [CrossRef]
- Olvera-Novoa, M.A.; Pereira-Pacheco, F.; Olivera-Castillo, L.; Pérez-Flores, V.; Navarro, L.; Sámano, J.C. Cowpea (Vigna unguiculata) protein concentrate as replacement for fish meal in diets for tilapia (Oreochromis niloticus) fry. Aquaculture 1997, 158, 107–116. [Google Scholar] [CrossRef]
- Gatlin, D.M., III; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, T.G.; Hardy, R.W.; Wurtele, E. Expanding the utilization of sustainable plant products in aquafeeds: A review. Aquac. Res. 2007, 38, 551–579. [Google Scholar] [CrossRef]
- Turchini, G.M.; Trushenski, J.T.; Glencross, B.D. Thoughts for the future of Aquaculture nutrition: Realigning perspectives to reflect contemporary issues related to judicious use of marine resources in aquafeeds. N. Am. J. Aquac. 2019, 81, 13–39. [Google Scholar] [CrossRef]
- Tacon, A.G.J.; Metian, M. Feed matters: Satisfying the feed demand of aquaculture. Rev. Fish. Sci. Aquac. 2015, 23, 1–10. [Google Scholar] [CrossRef]
- Alltech. Global Feed Survey. 2019. Available online: Alltechfeedsurvey.com (accessed on 15 May 2021).
- FAO (Food and Agriculture Organization of the United Nations). The State of World Fisheries and Aquaculture; SOFIA: Rome, Italy, 2020. [Google Scholar]
- Escobar, N.; Tizado, E.J.; zu Ermgassen, E.K.; Löfgren, P.; Börner, J.; Godar, J. Spatially-explicit footprints of agricultural commodities: Mapping carbon emissions embodied in Brazil’s soy export. Glob. Environ. Change 2020, 62, 102067. [Google Scholar] [CrossRef]
- Stone, D.A.J.; Allan, G.L.; Parkinson, S.; Frances, J. Replacement of fishmeal in diets for Australian silver perch, Bidyanus bidyanus: Digestibility of alternative ingredients. Aquaculture 2000, 186, 293–310. [Google Scholar] [CrossRef]
- Elharadallou, S.B.; Khalid, I.I.; Gobouri, A.A.; Abdel-Hafez, S.H. Amino Acid Composition of Cowpea (Vigna ungiculata L. Walp) Flour and Its Protein Isolates. Food Nutr. Sci. 2015, 6, 790–797. [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations). Commodity Price. Available online: http://www.fao.org/prices/en/ (accessed on 15 May 2021).
- Index Mundi Commodity Prices. Available online: https://www.indexmundi.com/commodities/ (accessed on 15 May 2021).




{kind=link}
{kind=link}
{kind=link}
| Location | Climate File No. | Lat | Long | Mean Annual Rain (mm) | CV of Annual Rainfall | Aridity of Summer (Nov–Apr) | Mean Daily Temperatures during Summer (Nov–Apr) (°C) | APSoil No. | PAWC (mm) | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Max. | Min. | Mungbean | Cowpea | Sorghum | Soybean | ||||||||
| Biloela | 39006 | −24.379 | 150.516 | 662 | 0.26 | 0.39 | 31.8 | 17.7 | 1186 | 175 | 215 | 230 | 189 |
| Emerald | 35264 | −23.569 | 148.176 | 605 | 0.33 | 0.34 | 33.0 | 19.8 | 1175 | 272 | 272 | 272 | 272 |
| Gatton | 40082 | −27.544 | 152.338 | 783 | 0.28 | 0.54 | 29.7 | 17.5 | 148 | 174 | 249 | 249 | 195 |
| Cecil Plains | 41019 | −27.723 | 151.287 | 643 | 0.26 | 0.34 | 30.0 | 15.9 | 198 | 255 | 255 | 225 | |
| Roma | 43091 | −26.548 | 148.771 | 599 | 0.33 | 0.28 | 32.3 | 17.8 | 125 | 130 | 166 | 153 | 149 |
| Gunnedah | 55202 | −30.954 | 150.249 | 606 | 0.26 | 0.29 | 30.2 | 15.1 | 213 | 165 | 219 | 273 | 219 |
| Coonamble | 51161 | −30.978 | 148.380 | 494 | 0.33 | 0.21 | 31.5 | 16.5 | 168 | 134 | 181 | 181 | 181 |
| Pulse | Protein | Fat | Total Carbohydrate | Ash |
|---|---|---|---|---|
| Cowpea (Vigna unguiculata) a | 22.8 | 1.5 | 61.7 | 3.5 |
| Kidney bean (Phaseolus vulgaris) a | 22.5 | 1.5 | 61.9 | 3.7 |
| Lentil (Lens esculenta) a | 24.7 | 1.1 | 60.1 | 3.0 |
| Pigeon pea (Cajanus cajan) a | 20.4 | 1.4 | 63.7 | 3.7 |
| Groundnut (Arachis hypogaea) a | 26.3 | 48.4 | 17.6 | 2.3 |
| Soybean (Glycine max) a | 34.1 | 17.7 | 33.5 | 4.7 |
| Faba bean (Vicia faba) b | 24.2 | 1.2 | - | 2.7 |
| Mung bean (Vigna radiata) b | 25.9 | 0.8 | - | 3.6 |
| Pulse | Protein | Carbohydrate | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Crude Protein (%) | Albumins (% of Protein) | Globulins (% of Protein) | Glutelin (% of Protein) | Prolamin (% of Protein) | Total (%) | Sugars (%) | Starch (%) | Dietary Fibre (%) | |
| Cowpea | 24–28 | 4–12 | 58–80 | 10–15 | 1–3 | 42–63 | 3–10 | 35–52 | 11-34 |
| Kidney bean | 17–27 | 15 | 60 | N/a | N/a | 63–74 | 8–11 | 31–43 | 18–30 |
| Chickpea | 19–27 | 8–12 | 53–60 | 18–24 | 3–7 | 52–71 | 3–5 | 30–56 | 6–15 |
| Lentil | 23–31 | 17 | 51 | 11 | 4 | 42–72 | 5–6 | 37–59 | 7–23 |
| Pea | 14–31 | 15–25 | 49–70 | 11 | 5 | 55–72 | 5–12 | 30–49 | 3–20 |
| Lupin | 32–44 | 9–22 | 44–60 | 6–23 | 47 | 10–16 | 1–9 | 14–55 | |
| Protein (%) | Lysine (%) | Methionine (%) | Price of Commodity (USD/MT-2020–2021) e,f | |
|---|---|---|---|---|
| Fishmeal (herring) a | 72 | 7.30 | 2.20 | 1500 |
| Soybean meal a | 44 | 2.83 | 0.61 | ~440 |
| Soy protein concentrate a | 64 | 3.93 | 0.81 | ~565 |
| Soy protein isolate a | 81 | 3.02 | 1.15 | ~1000 |
| Canola meal solvent extracted a | 38 | 2.02 | 0.77 | ~400 |
| Canola protein concentrate a | 69 | 3.10 | 1.26 | - |
| Cottonseed meal solvent extracted a | 42 | 1.60 | 0.58 | 178 |
| Sunflower meal solvent extracted a | 32 | 1.20 | 0.82 | ~300–500 |
| Lupin meal a | 30 | 1.54 | 0.27 | ~80–210 |
| Wheat flour a | 12 | 0.58 | 0.19 | ~240 |
| Barley whole grain a | 11 | 0.53 | 0.18 | ~80–120 |
| Dehulled cowpea meal b | 21.3–25.6 | 7.0–7.5 | 1.4–2.2 | ~360 |
| Cowpea protein isolate c,d | 75 c | 6.8 d | 1.4 d | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bell, L.W.; James, A.T.; Augustin, M.A.; Rombenso, A.; Blyth, D.; Simon, C.; Higgins, T.J.V.; Barrero, J.M. A Niche for Cowpea in Sub-Tropical Australia? Agronomy 2021, 11, 1654. https://doi.org/10.3390/agronomy11081654
Bell LW, James AT, Augustin MA, Rombenso A, Blyth D, Simon C, Higgins TJV, Barrero JM. A Niche for Cowpea in Sub-Tropical Australia? Agronomy. 2021; 11(8):1654. https://doi.org/10.3390/agronomy11081654
Chicago/Turabian StyleBell, Lindsay W., Andrew T. James, Mary Ann Augustin, Artur Rombenso, David Blyth, Cedric Simon, Thomas J. V. Higgins, and Jose M. Barrero. 2021. "A Niche for Cowpea in Sub-Tropical Australia?" Agronomy 11, no. 8: 1654. https://doi.org/10.3390/agronomy11081654
APA StyleBell, L. W., James, A. T., Augustin, M. A., Rombenso, A., Blyth, D., Simon, C., Higgins, T. J. V., & Barrero, J. M. (2021). A Niche for Cowpea in Sub-Tropical Australia? Agronomy, 11(8), 1654. https://doi.org/10.3390/agronomy11081654