
Impacts of Litter Composition on the Structure and Functional Pathways of Soil Microbial Community during Phyllostachys Edulis Expansion



Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil and Litter Sampling
2.2. Experimental Design
2.3. Analyses of Soil Properties and Soil Enzyme Activities
2.4. DNA Extraction and Amplicon Sequencing
2.5. Statistical Analyses
3. Results
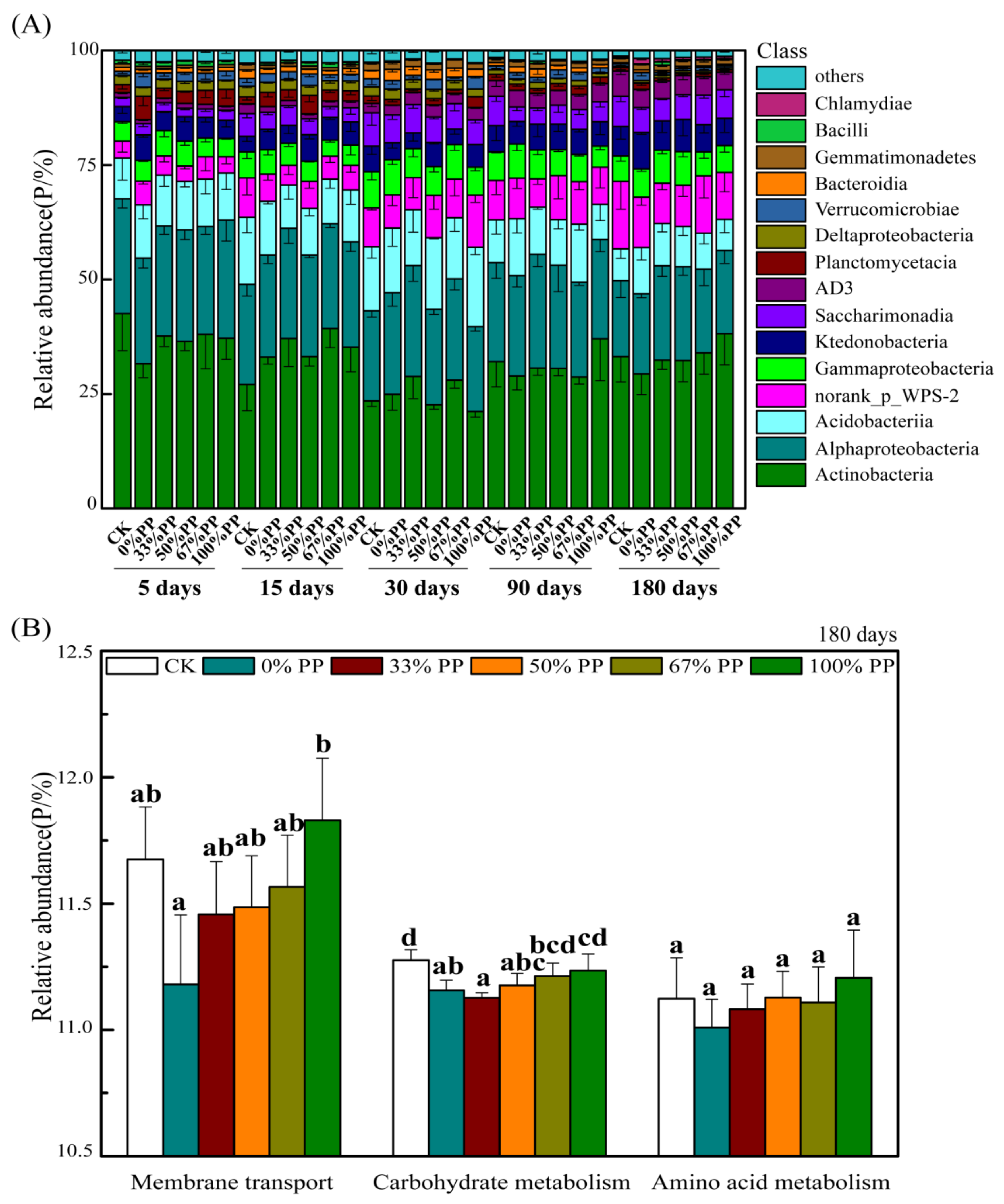
3.1. Bacterial Community Composition and Functional Pathways
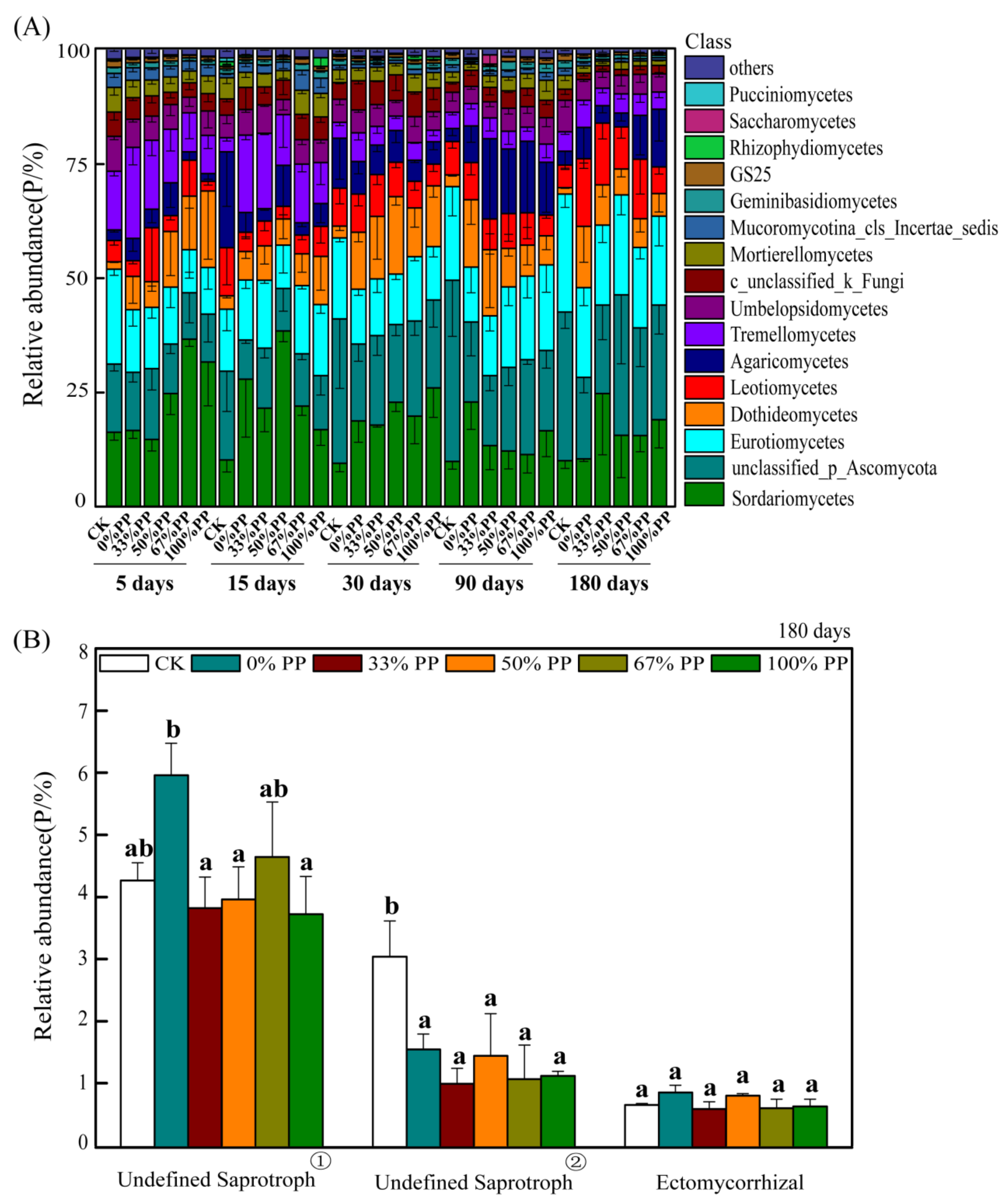
3.2. Fungal Community Composition and Functional Pathways
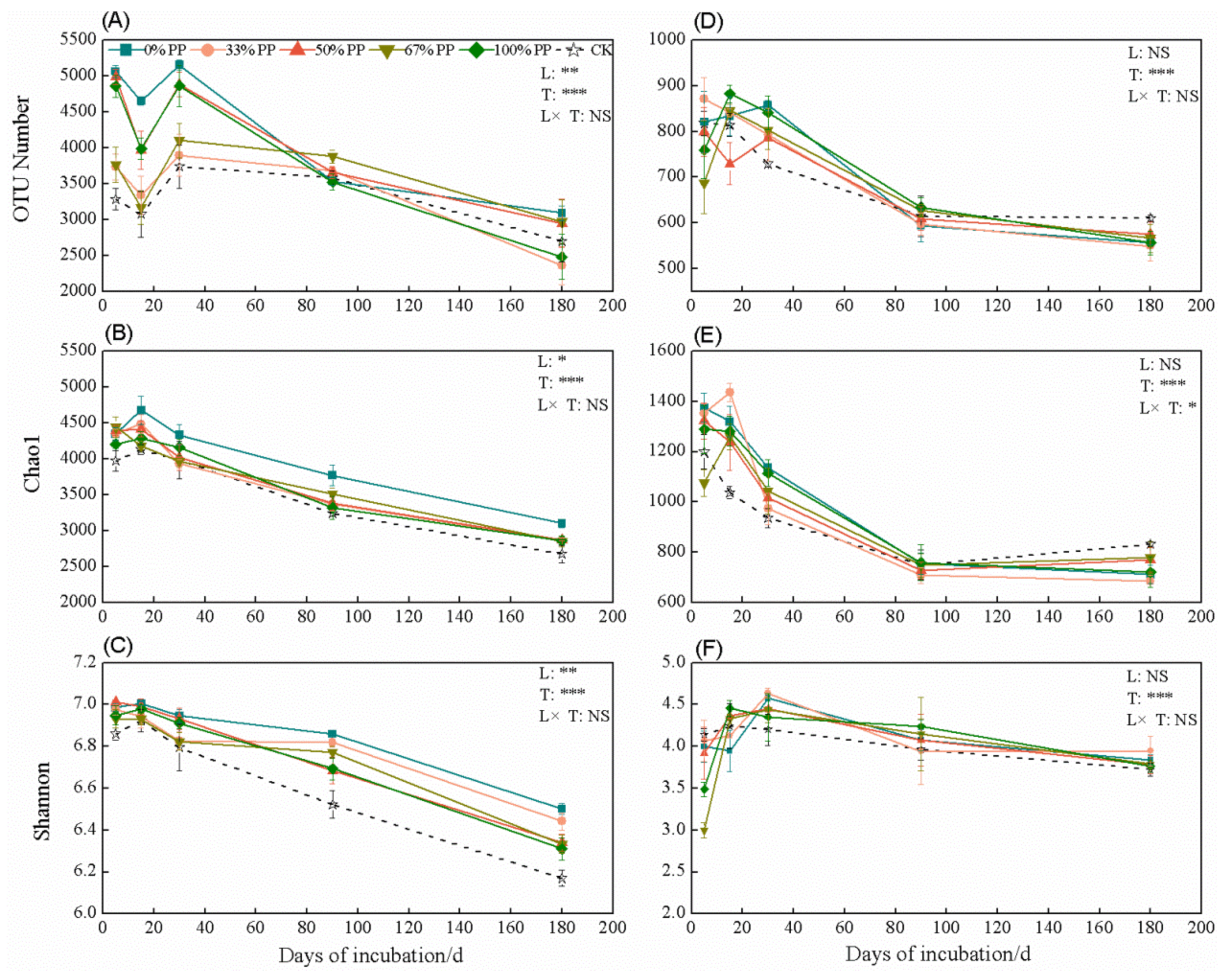
3.3. Bacterial and Fungal α-Diversity Vary with Litter Type and Incubation Time
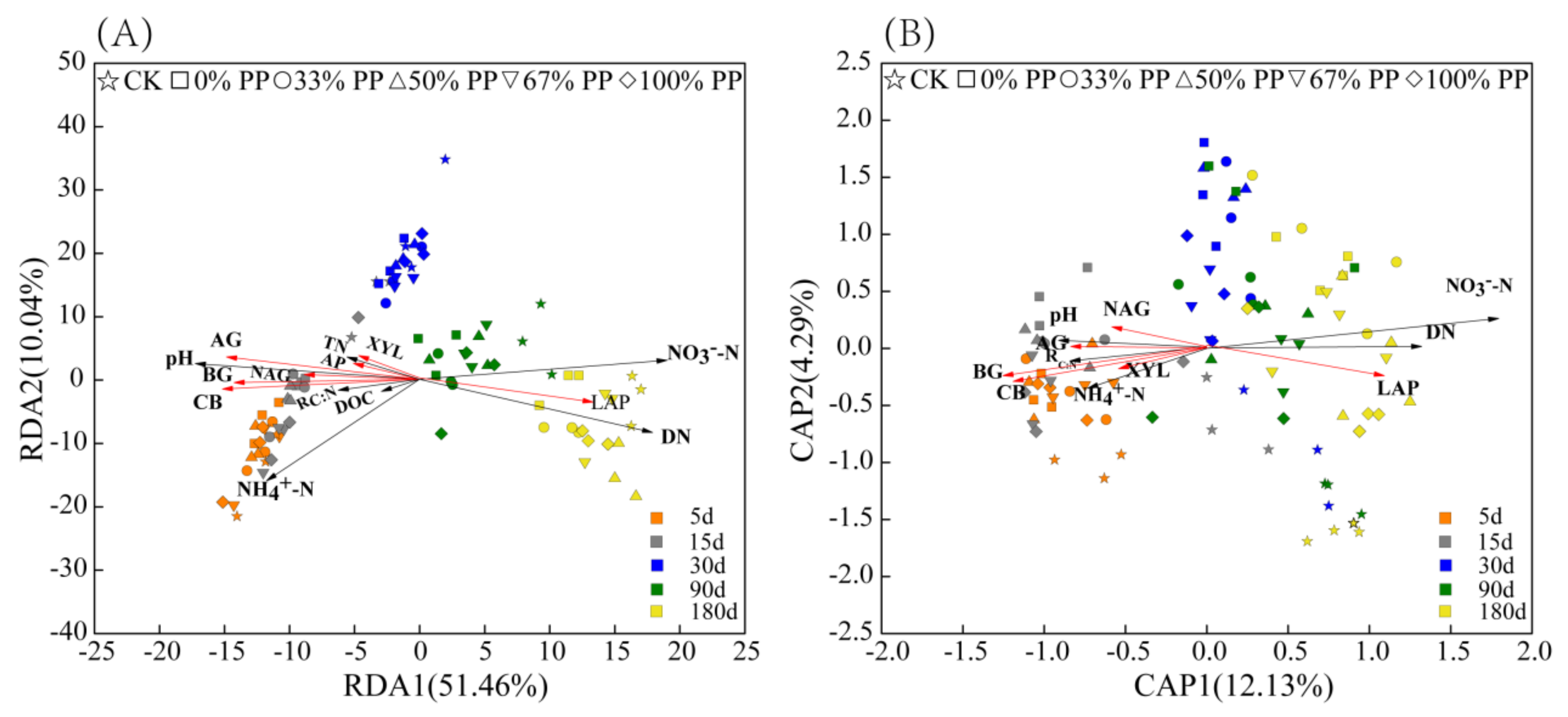
3.4. Contributions of Environmental Factors to Microbial Community Structure
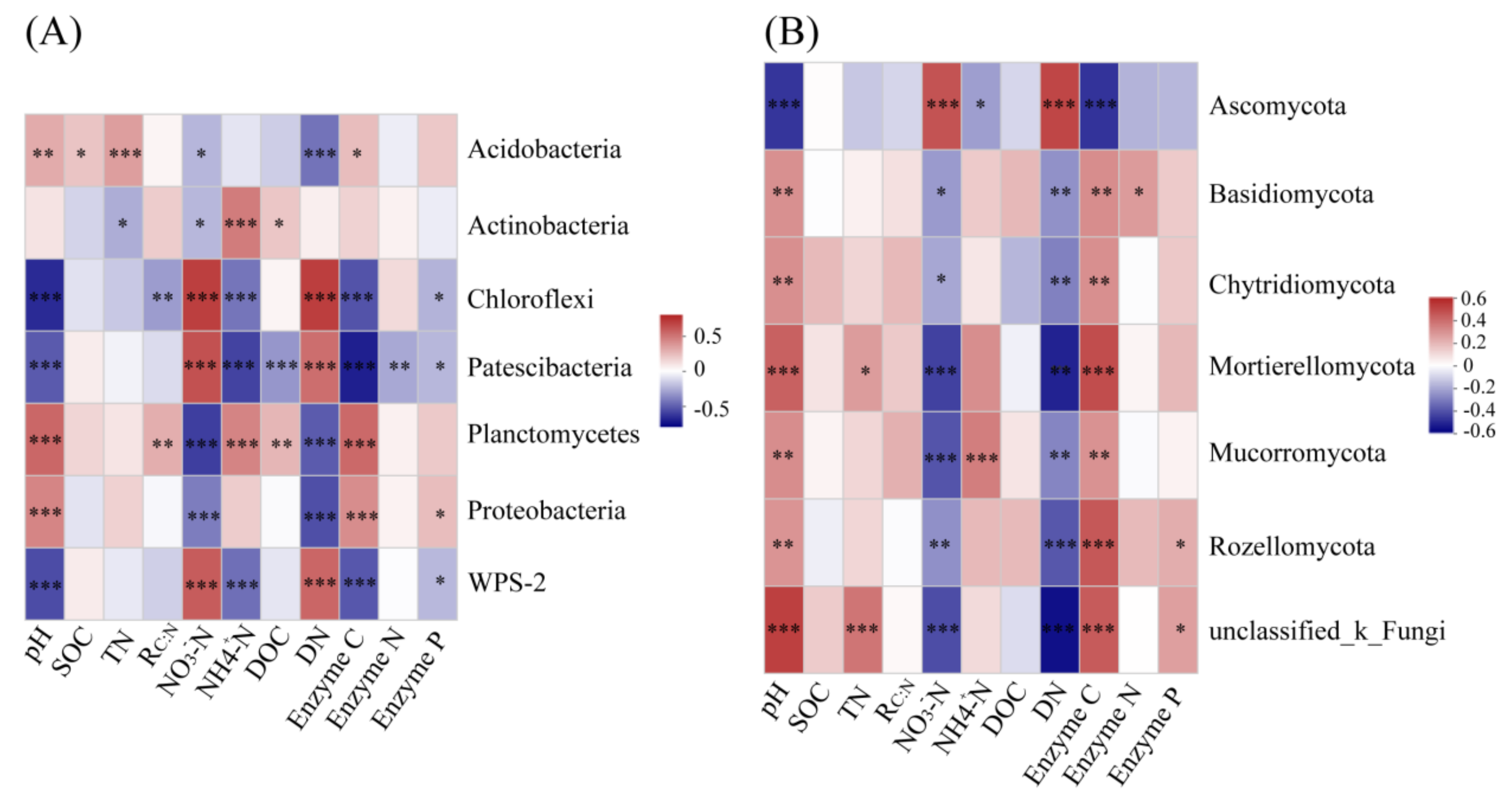
3.5. Impact of Environmental Factors on Bacterial and Fungal Taxa
4. Discussion
4.1. Effects of Litter on Soil Bacterial and Fungal Community Structure
4.2. Effect of Bamboo Litter on Bacterial and Fungal Community Composition and Function Pathways
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, R.; Zhang, Y.; Hu, X.F.; Wan, S.Z.; Wang, H.M.; Liang, C.; Chen, F.S. Litter manipulation effects on microbial communities and enzymatic activities vary with soil depth in a subtropical Chinese fir plantation. For. Ecol. Manag. 2021, 480, 118641. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.A.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Wang, W.B.; Chen, D.S.; Sun, X.M.; Zhang, Q.; Koide, R.T.; Insam, H.; Zhang, S.G. Impacts of mixed litter on the structure and functional pathway of microbial community in litter decomposition. Appl. Soil Ecol. 2019, 144, 72–82. [Google Scholar] [CrossRef]
- Yang, W.; Yan, Y.E.; Jiang, F.; Leng, X.; Cheng, X.L.; An, S.Q. Response of the soil microbial community composition and biomass to a short-term Spartina alterniflora invasion in a coastal wetland of eastern China. Plant Soil 2016, 408, 443–456. [Google Scholar] [CrossRef]
- Wang, Y.J.; Liu, L.; Yang, J.F.; Duan, Y.M.; Luo, Y.; Taherzadeh, M.J.; Li, Y.F.; Li, H.K.; Awasthi, M.K.; Zhao, Z.Y. The diversity of microbial community and function varied in response to different agricultural residues composting. Sci. Total Environ. 2020, 715, 136983. [Google Scholar] [CrossRef]
- Agnieszka, K.B.; Wojciech, B.; Edyta, S.; Agnieszka, B.; Lynn, B.; Gabriela, W. The role of plants and soil properties in the enzyme activities of substrates on hard coal mine spoil heaps. Sci. Rep. 2021, 11, 5155. [Google Scholar]
- Bai, X.J.; Dippold, M.A.; An, S.S.; Wang, B.R.; Zhang, H.X.; Loeppmann, S. Extracellular enzyme activity and stoichiometry: The effect of soil microbial element limitation during leaf litter decomposition. Ecol. Indic. 2021, 121, 107200. [Google Scholar] [CrossRef]
- Luan, J.W.; Li, S.Y.; Dong, W.; Liu, Y.C.; Wang, Y.; Liu, S.R. Litter decomposition affected by bamboo expansion is modulated by litter-mixing and microbial composition. Funct. Ecol. 2021, 35, 2562–2574. [Google Scholar] [CrossRef]
- Chapman, S.K.; Newman, G.S. Biodiversity at the plant–soil interface: Microbial abundance and community structure respond to litter mixing. Oecologia 2010, 162, 763–769. [Google Scholar] [CrossRef]
- Liu, C.C.; Liu, Y.G.; Guo, K.; Zhao, H.W.; Qiao, X.G.; Wang, S.J.; Zhang, L.; Cai, X.L. Mixing litter from deciduous and evergreen trees enhances decomposition in a subtropical karst forest in southwestern China. Soil Biol. Biochem. 2016, 101, 44–54. [Google Scholar] [CrossRef]
- Li, H.Y.; Wei, Z.S.; Huangfu, C.H.; Chen, X.W.; Yang, D.L. Litter mixture dominated by leaf litter of the invasive species, Flaveria bidentis, accelerates decomposition and favors nitrogen release. J. Plant Res. 2017, 130, 167–180. [Google Scholar] [CrossRef]
- Sun, H.; Wang, Q.X.; Zhang, C.G.; Li, L.; Liu, Z.B.; Liu, N.; Shao, C.; Zhang, Y.Y.; Yan, J.; Li, Y.X. Effects of different leaf litters on the physicochemical properties and soil microbial communities in Panax ginseng-growing soil. Acta Ecol. Sinica 2018, 38, 3603–3615. [Google Scholar]
- Shiau, Y.J.; Chiu, C.Y. Changes in Soil Biochemical Properties in a Cedar Plantation Invaded by Moso Bamboo. Forests 2017, 8, 222. [Google Scholar] [CrossRef]
- You, Y.M.; Wang, J.; Huang, X.M.; Tang, Z.X.; Liu, S.R.; Sun, O.J. Relating microbial community structure to functioning in forest soil organic carbon transformation and turnover. Ecol. Evol. 2014, 4, 633–647. [Google Scholar] [CrossRef]
- Liu, Y.X.; Wang, S.L.; Wang, Q.K.; Zhang, J. Effects of mixed-species leaf litter on litter decomposition and soil microbial communities in experimental subtropical plantation forest. J. Food Agric. Environ. 2010, 8, 1102–1107. [Google Scholar]
- Pei, Z.Q.; Leppert, K.N.; Eichenberg, D.; Bruelheide, H.; Niklaus, P.A.; Buscot, F.; Gutknecht, J.L.M. Leaf litter diversity alters microbial activity, microbial abundances, and nutrient cycling in a subtropical forest ecosystem. Biogeochemistry 2017, 134, 163–181. [Google Scholar] [CrossRef]
- Santonja, M.; Rancon, A.; Fromin, N.; Baldy, V.; Hättenschwiler, S.; Fernandez, C.; Montès, N.; Mirleau, P. Plant litter diversity increases microbial abundance, fungal diversity, and carbon and nitrogen cycling in a Mediterranean shrubland. Soil Biol. Biochem. 2017, 111, 124–134. [Google Scholar] [CrossRef]
- Li, Y.B.; Bezemer, T.M.; Yang, J.J.; Lü, X.T.; Li, X.Y.; Liang, W.J.; Han, X.G.; Li, Q. Changes in litter quality induced by N deposition alter soil microbial communities. Soil Biol. Biochem. 2019, 130, 33–42. [Google Scholar] [CrossRef]
- Schneider, T.; Keiblinger, K.M.; Schmid, E.; Sterflinger-Gleixner, K.; Ellersdorfer, G.; Roschitzki, B.; Richter, A.; Eberl, L.; Zechmeister-Boltenstern, S.; Riedel, K. Who is who in litter decomposition? Metaproteomics reveals major microbial players and their biogeochemical functions. ISME J. 2012, 6, 1749–1762. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Shi, W. Short-term effects of plant litter on the dynamics, amount, and stoichiometry of soil enzyme activity in agroecosystems. Eur. J. Soil Biol. 2014, 65, 23–29. [Google Scholar] [CrossRef]
- Shi, L.; Fan, S.H.; Jiang, Z.H.; Qi, L.H.; Liu, G.L. Mixed leaf litter decomposition and N, P release with a focus on Phyllostachys edulis (Carrière) J. Houz. forest in subtropical southeastern China. Acta Soc. Bot. Pol. 2015, 84, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.Z.; Liang, C.F.; Shao, S.; Chen, J.H.; Qin, H.; Xu, Q.F. Linkages of litter and soil C:N:P stoichiometry with soil microbial resource limitation and community structure in a subtropical broadleaf forest invaded by Moso bamboo. Plant Soil 2021, 465, 473–490. [Google Scholar] [CrossRef]
- Liu, J.L.; Wang, G.H.; Xu, D.M. Soil organic carbon mineralization response to litter addition in desert steppe grassland sites with differences in number of years grazing by fencing. Acta Prataculturae Sin. 2019, 28, 47–57. [Google Scholar]
- Lu, R.K. Methods of Soil and Agro-chemical Analysis. China Agric. Science Technol. Press 2000. [Google Scholar]
- Justine, M.F.; Yang, W.Q.; Wu, F.Z.; Tan, B.; Naeem Khan, M.; Li, Z.J. Dissolved organic matter in soils varies across a cheonsequence of Pinus massoniana plantations. Ecosphere 2017, 8, e01764. [Google Scholar] [CrossRef]
- Lyu, M.K.; Xie, J.S.; Vadeboncoeur, M.A.; Wang, M.H.; Qiu, X.; Ren, Y.B.; Jiang, M.H.; Yang, Y.S.; Kuzyakov, Y. Simulated leaf litter addition causes opposite priming effects on natural forest and plantation soils. Biol. Fertil. Soils 2018, 54, 925–934. [Google Scholar] [CrossRef]
- Shahzad, T.; Anwar, F.; Hussain, S.; Mahmood, F.; Arif, M.S.; Sahar, A.; Nawaz, M.F.; Perveen, N.; Sanaullah, M.; Rehman, K.; et al. Carbon dynamics in surface and deep soil in response to increasing litter addition rates in an agro-ecosystem. Geoderma 2019, 333, 1–9. [Google Scholar] [CrossRef]
- Chen, J.H.; Chen, D.; Xu, Q.F.; Fuhrmann, J.J.; Li, L.Q.; Pan, G.X.; Li, Y.F.; Qin, H.; Liang, C.F.; Sun, X. Organic carbon quality, composition of main microbial groups, enzyme activities, and temperature sensitivity of soil respiration of an acid paddy soil treated with biochar. Biol. Fertil. Soils 2019, 55, 185–197. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Koljalg, U.; Larsson, K.H.; Abarenkov, K.; Nilsson, R.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Hoiland, K.; Kjoller, R.; Larsson, E.; et al. UNITE: A database providing web-based methods for the molecular identification of ectomycorrhizal fungi. New Phytol. 2005, 166, 1063–1068. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Zhang, W.W.; Lu, Z.T.; Yang, K.; Zhu, J.J. Impacts of conversion from secondary forests to larch plantations on the structure and function of microbial communities. Appl. Soil Ecol. 2017, 111, 73–83. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, X.Y.; Li, Y.C.; Chen, Z.H.; Yao, Z.X.; Fang, T.; Zhang, B.G. Effects of litter addition of Moso Bamboo (Phyllostachys edulis) and broadeaf forest on soil bacteria community. J. Agric. Biotechnol. 2020, 28, 951–962. [Google Scholar]
- Xing, W.; Lu, X.M.; Xu, F.W.; Ying, J.Y.; Chen, D.M.; Bai, Y.F. Linking microbial community structure to carbon substrate chemistry in soils following aboveground and belowground litter additions. Appl. Soil Ecol. 2019, 141, 18–25. [Google Scholar] [CrossRef]
- Zhang, B.B.; Wan, X.H.; Yang, J.Q.; Wang, T.; Huang, Z.Q. Effects of litters different in quality on soil microbial community structure in Cunninghamia lanceolata plantation. Acta Pedol. Sinica 2021, 58, 1040–1049. [Google Scholar] [CrossRef]
- Dong, H.Y.; Ge, J.F.; Sun, K.; Wang, B.Z.; Xue, J.M.; Wakelin, S.A.; Wu, J.S.; Sheng, W.X.; Liang, C.F.; Xu, Q.F.; et al. Change in root-associated fungal communities affects soil enzymatic activities during Pinus massoniana forest development in subtropical China. For. Ecol. Manag. 2021, 482, 118817. [Google Scholar] [CrossRef]
- Aragao, L.E.O.C.; Malhi, Y.; Metcalfe, D.B.; Silva-Espejo, J.E.; Jimenez, E.; Navarrete, D.; Almeida, S.; Costa, A.C.L.; Salinas, N.; Phillips, O.L.; et al. Above-and below-ground net primary productivity across ten Amazonian forests on contrasting soils. Biogeosciences 2009, 6, 2759–2778. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.F.; Wang, L.; Hu, Y.; Tsang, Y.F.; Wu, J.H.; Fu, X.H.; Sun, Y. Plant litter composition selects different soil microbial structures and in turn drives different litter decomposition pattern and soil carbon sequestration capability. Geoderma 2018, 319, 194–203. [Google Scholar] [CrossRef]
- Zhang, H.C.; Yuan, W.P.; Dong, W.J.; Liu, S.G. Seasonal patterns of litterfall in forest ecosystem worldwide. Ecol. Complex. 2014, 20, 240–247. [Google Scholar] [CrossRef]
- Wu, Z.X.; Hao, Z.P.; Sun, Y.Q.; Guo, L.P.; Huang, L.Q.; Zeng, Y.; Wang, Y.; Yang, L.; Chen, B.D. Comparison on the structure and function of the rhizosphere microbial community between healthy and root-rot Panax notoginseng. Appl. Soil Ecol. 2016, 107, 99–107. [Google Scholar] [CrossRef]
- Bromke, M.A. Amino Acid biosynthesis pathways in diatoms. Metabolites 2013, 3, 294–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.S.; Quadir, Q.F.; Rahman, A.; Asha, M.N.; Chowdhury, A.K. Screening and characterization of phosphorus solubilizing bacteria and their effect on rice seedlings. Res. Agric. Livest. Fish. 2014, 1, 27–35. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Litter Composition | Quantity of Litter/g 100 g−1 soil | Total C/mg g−1 | Total N/mg g−1 | C/N | Lignin/% | Cellulose/% |
|---|---|---|---|---|---|---|---|
| 0% PP | Broadleaved | 1 | 442.58 ± 10.78 d | 12.57 ± 1.38 a | 36.23 ± 4.27 a | 49.15 ± 1.15 b | 11.83 ± 0.53 a |
| 33% PP | Broadleaved + Bamboo | 0.667 + 0.333 | 414.76 ± 12.13 c | 12.21 ± 0.96 a | 34.61 ± 2.80 a | 48.48 ± 1.15 ab | 11.58 ± 0.49 a |
| 50% PP | Broadleaved + Bamboo | 0.5 + 0.5 | 400.84 ± 5.12 bc | 12.04 ± 0.78 a | 33.79 ± 2.16 a | 48.15 ± 1.15 ab | 11.46 ± 0.48 a |
| 67% PP | Broadleaved + Bamboo | 0.333 + 0.667 | 386.93 ± 3.23 b | 11.86 ± 0.64 a | 32.98 ± 1.64 a | 47.82 ± 1.15 ab | 11.33 ± 0.46 a |
| 100% PP | Bamboo | 1 | 359.10 ± 0.55 a | 11.50 ± 0.59 a | 31.35 ± 1.60 a | 45.15 ± 0.29 a | 10.45 ± 0.20 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, H.; Shao, S.; Liang, C.; Xu, Q.; Chen, J.; Qin, H. Impacts of Litter Composition on the Structure and Functional Pathways of Soil Microbial Community during Phyllostachys Edulis Expansion. Agronomy 2022, 12, 220. https://doi.org/10.3390/agronomy12010220
Dong H, Shao S, Liang C, Xu Q, Chen J, Qin H. Impacts of Litter Composition on the Structure and Functional Pathways of Soil Microbial Community during Phyllostachys Edulis Expansion. Agronomy. 2022; 12(1):220. https://doi.org/10.3390/agronomy12010220
Chicago/Turabian StyleDong, Huiyun, Shuai Shao, Chenfei Liang, Qiufang Xu, Junhui Chen, and Hua Qin. 2022. "Impacts of Litter Composition on the Structure and Functional Pathways of Soil Microbial Community during Phyllostachys Edulis Expansion" Agronomy 12, no. 1: 220. https://doi.org/10.3390/agronomy12010220
APA StyleDong, H., Shao, S., Liang, C., Xu, Q., Chen, J., & Qin, H. (2022). Impacts of Litter Composition on the Structure and Functional Pathways of Soil Microbial Community during Phyllostachys Edulis Expansion. Agronomy, 12(1), 220. https://doi.org/10.3390/agronomy12010220