
On the Use of Sap Flow Measurements to Assess the Water Requirements of Three Australian Native Tree Species




Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Tree Species and Sites
2.2. Measurement of Tree Characteristics
2.3. Determination of Wood Properties
2.4. Thermal Diffusivity Estimate
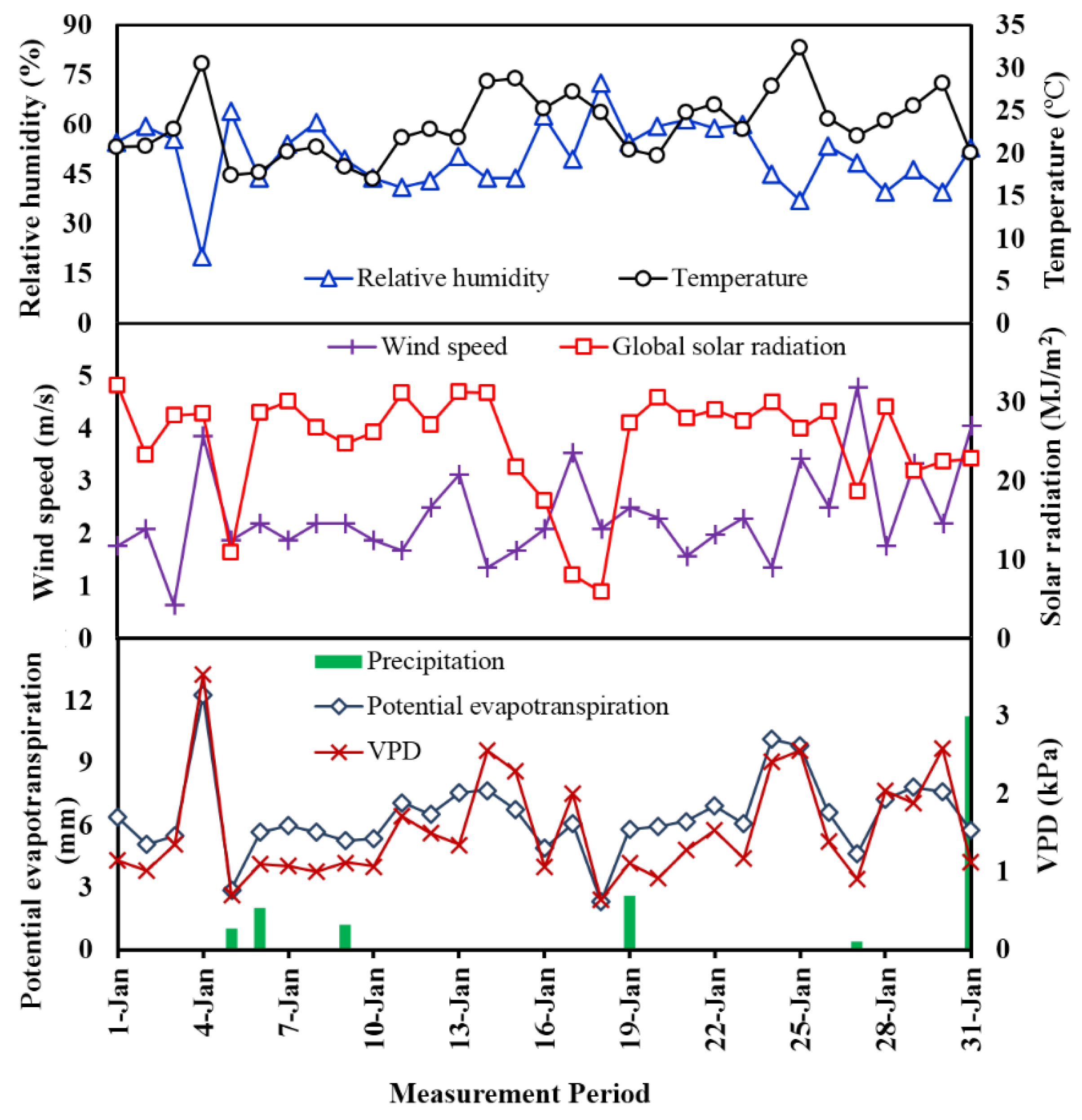
2.5. Meteorological and Environmental Measurement
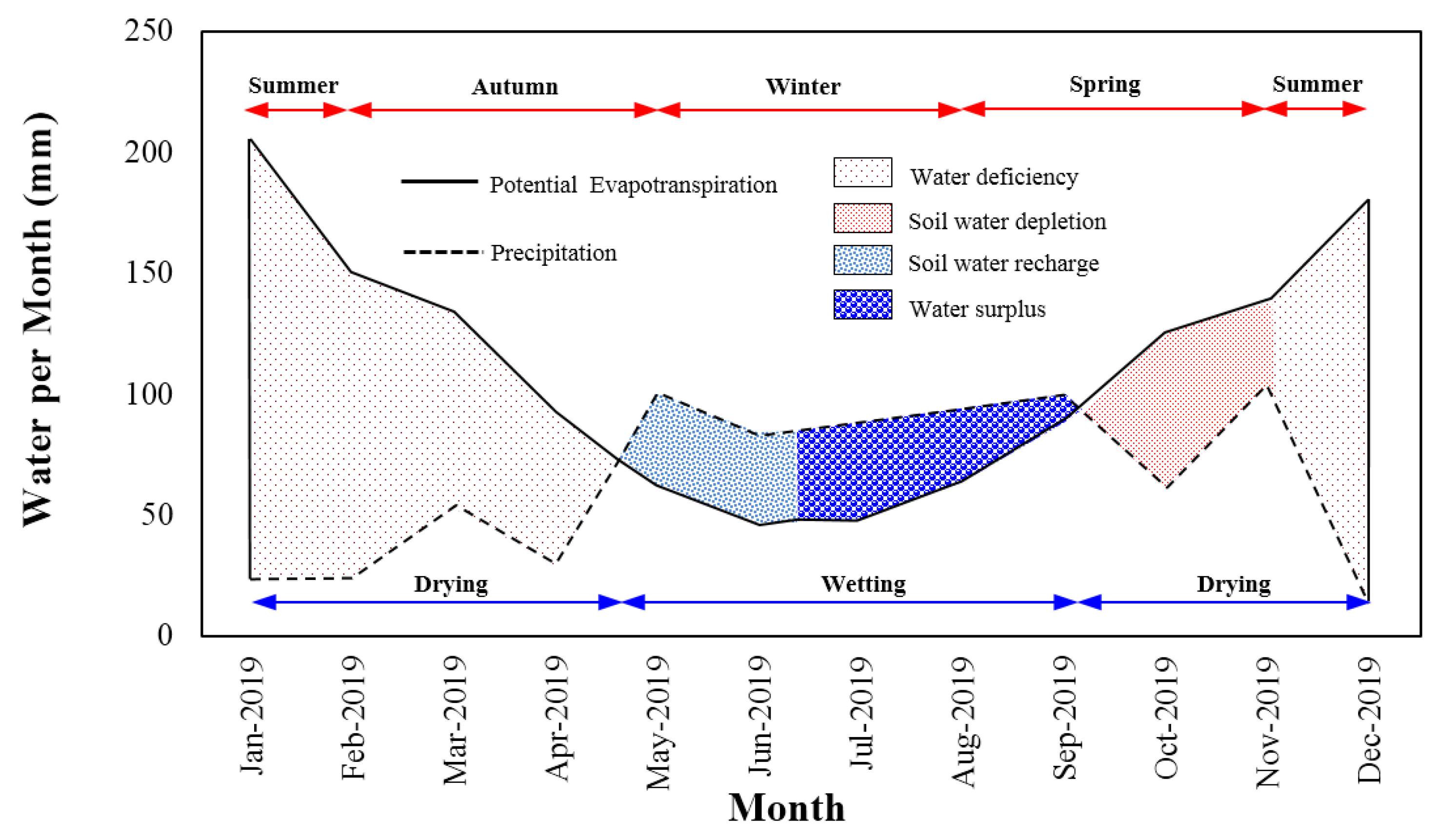
Soil Water Budget
2.6. Meteorological and Environmental Measurement
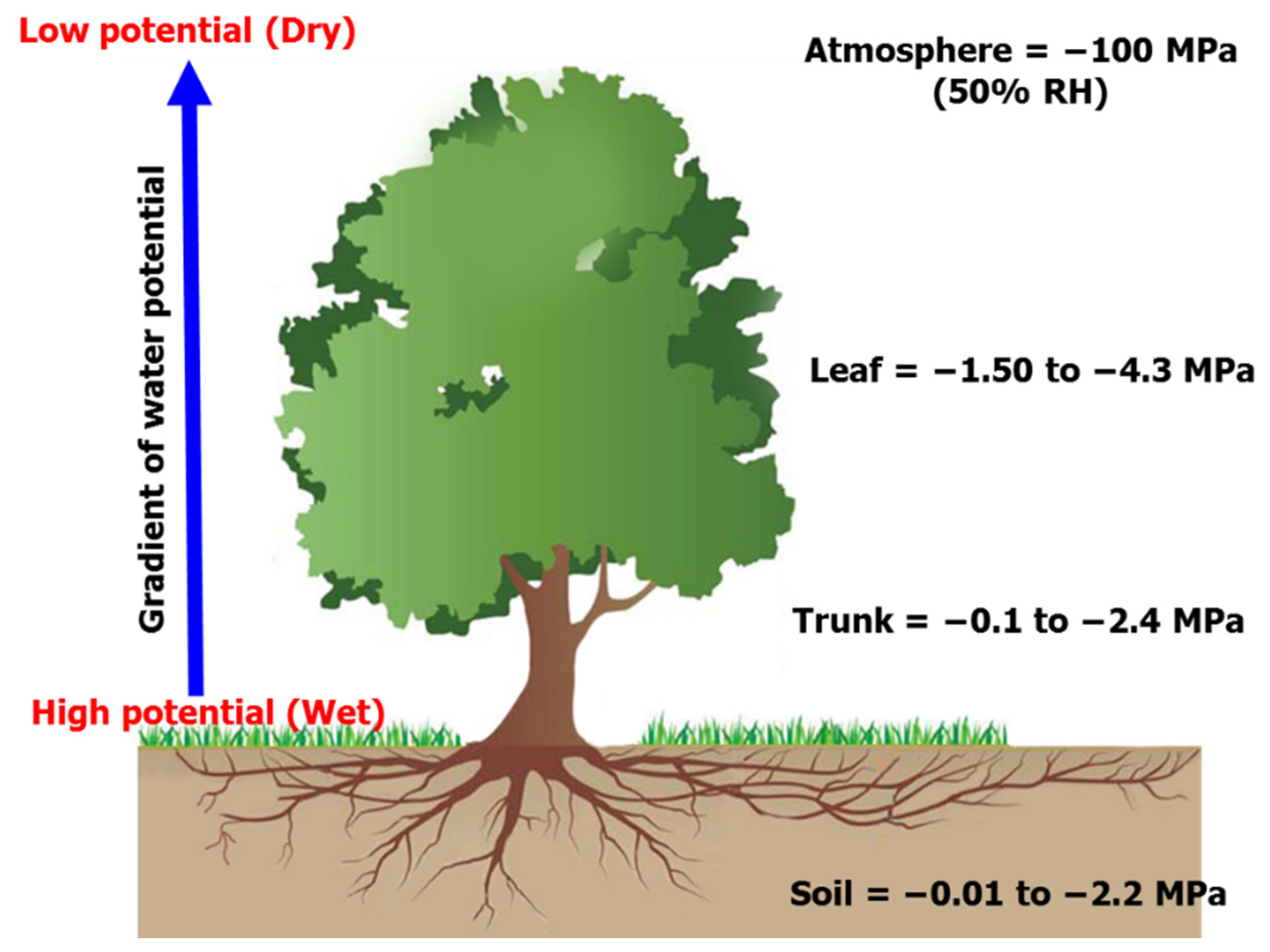
2.6.1. Soil Water Potential
2.6.2. Leaf Water Potential
2.6.3. Stem Water Potential
2.7. Sap Flow Instrument
2.8. Growth Rate and Temperature
3. Results and Discussions
3.1. Tree Information
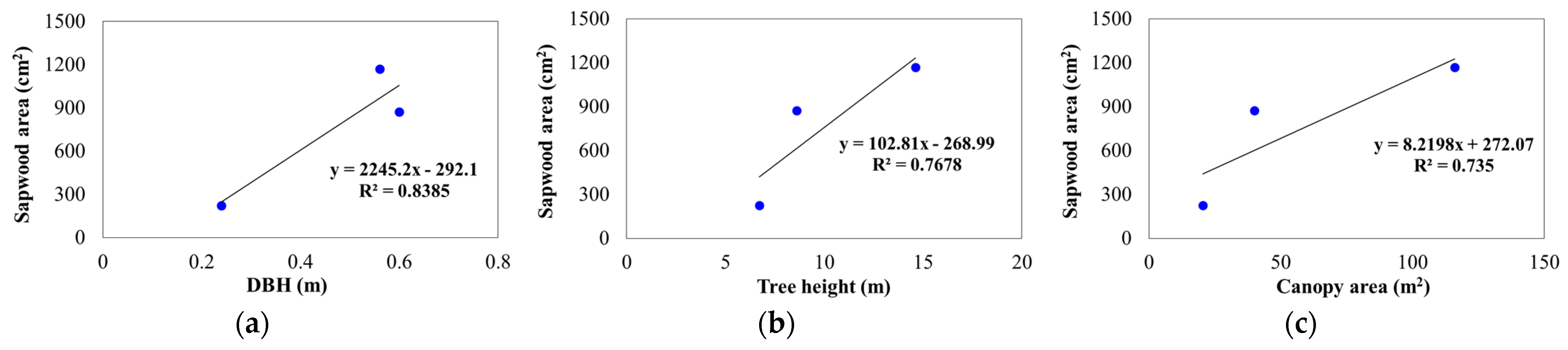
3.2. Wood Properties
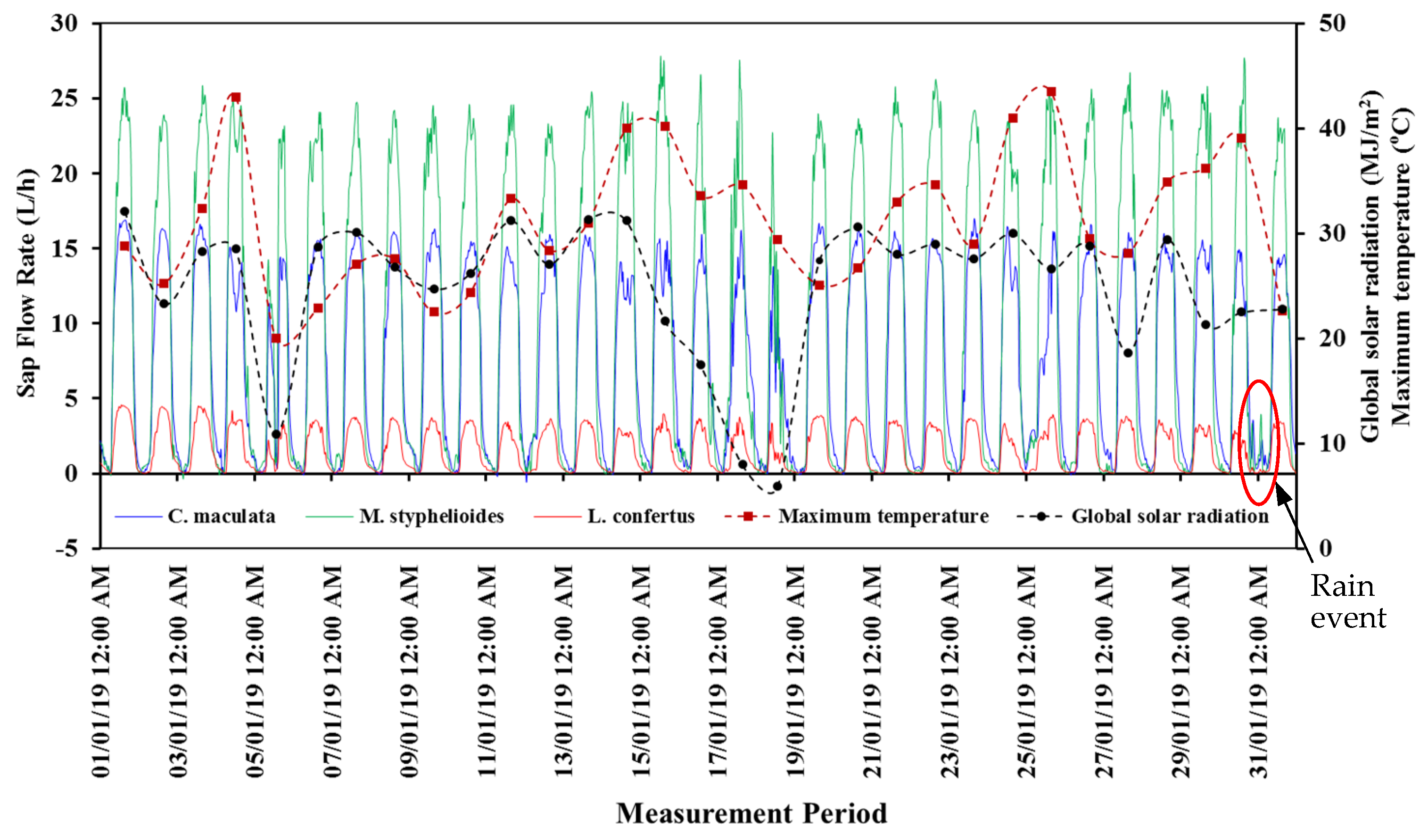
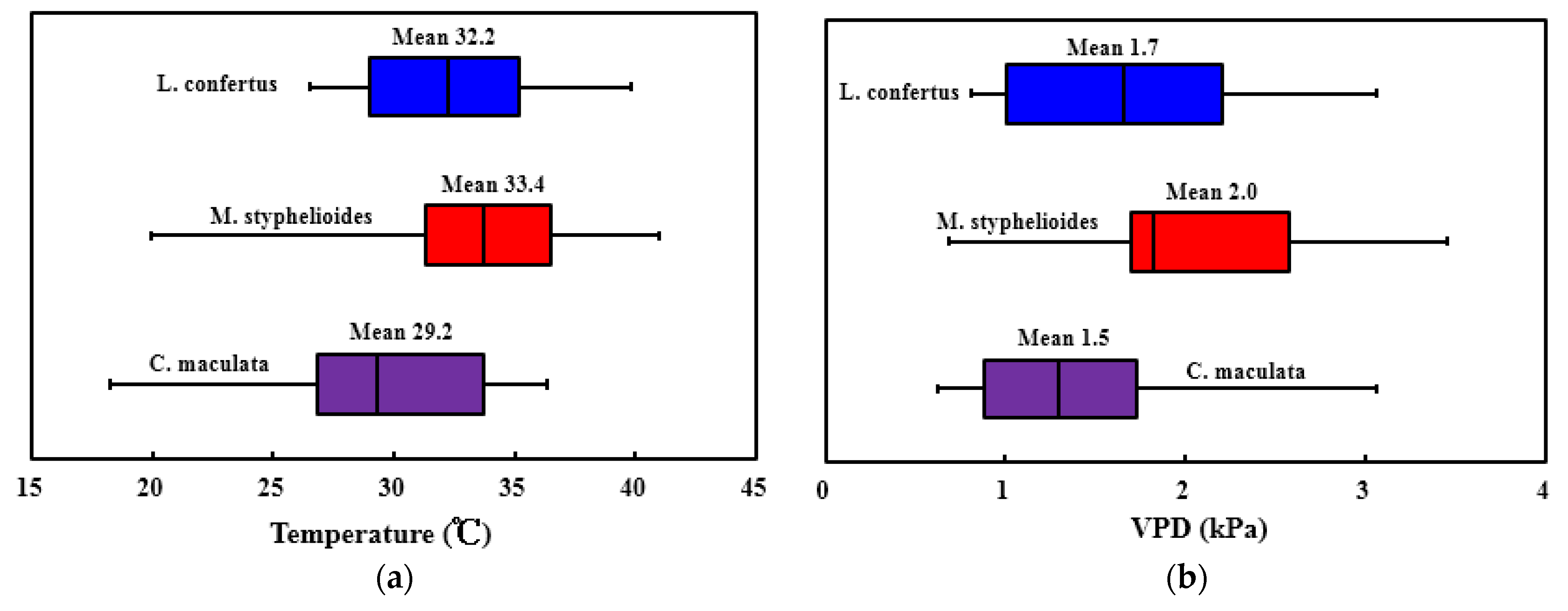
3.3. Meteorological Measurement
3.4. Soil-Plant-Atmosphere Continuum
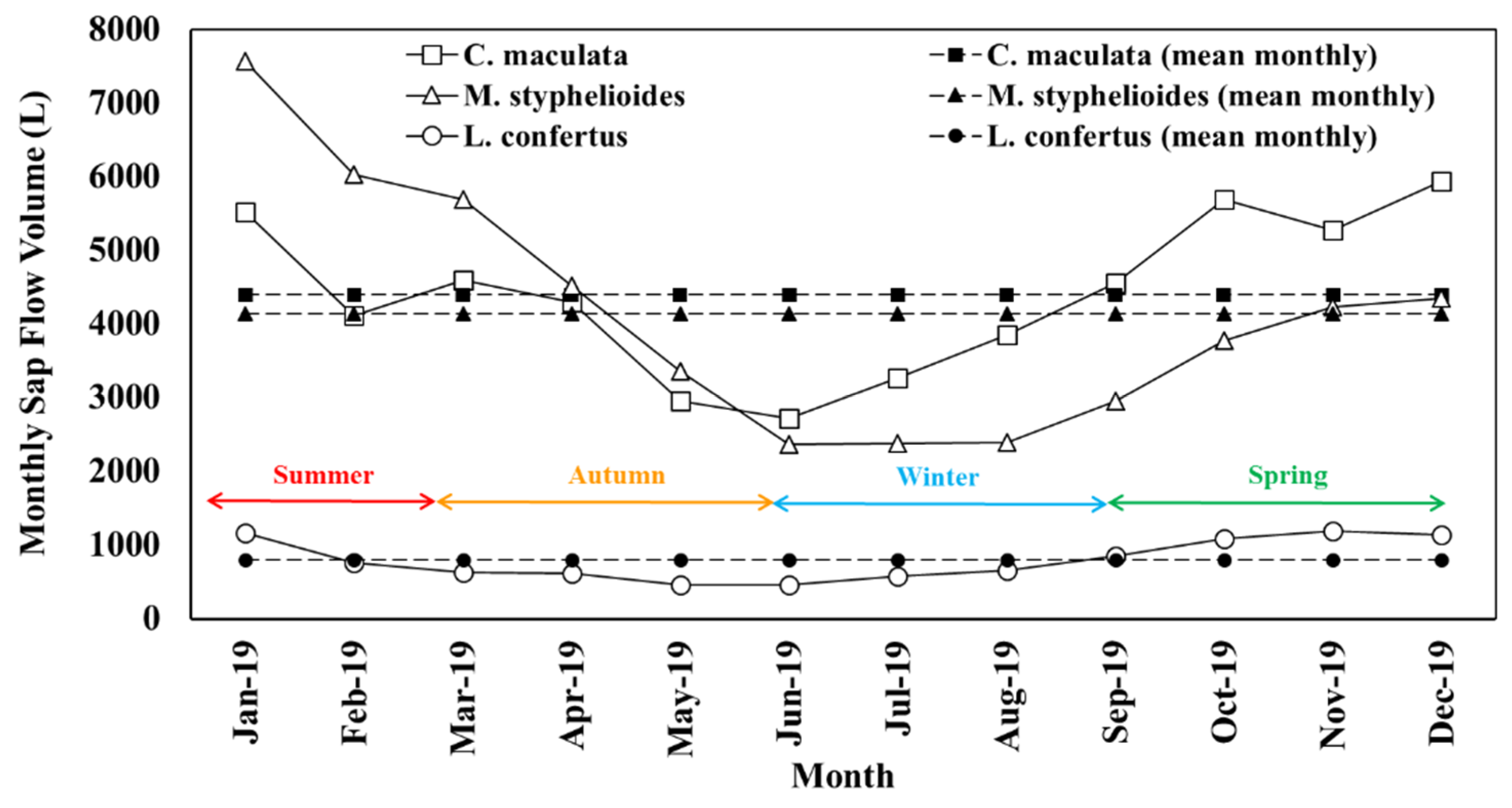
3.5. Sap Flow Measurement
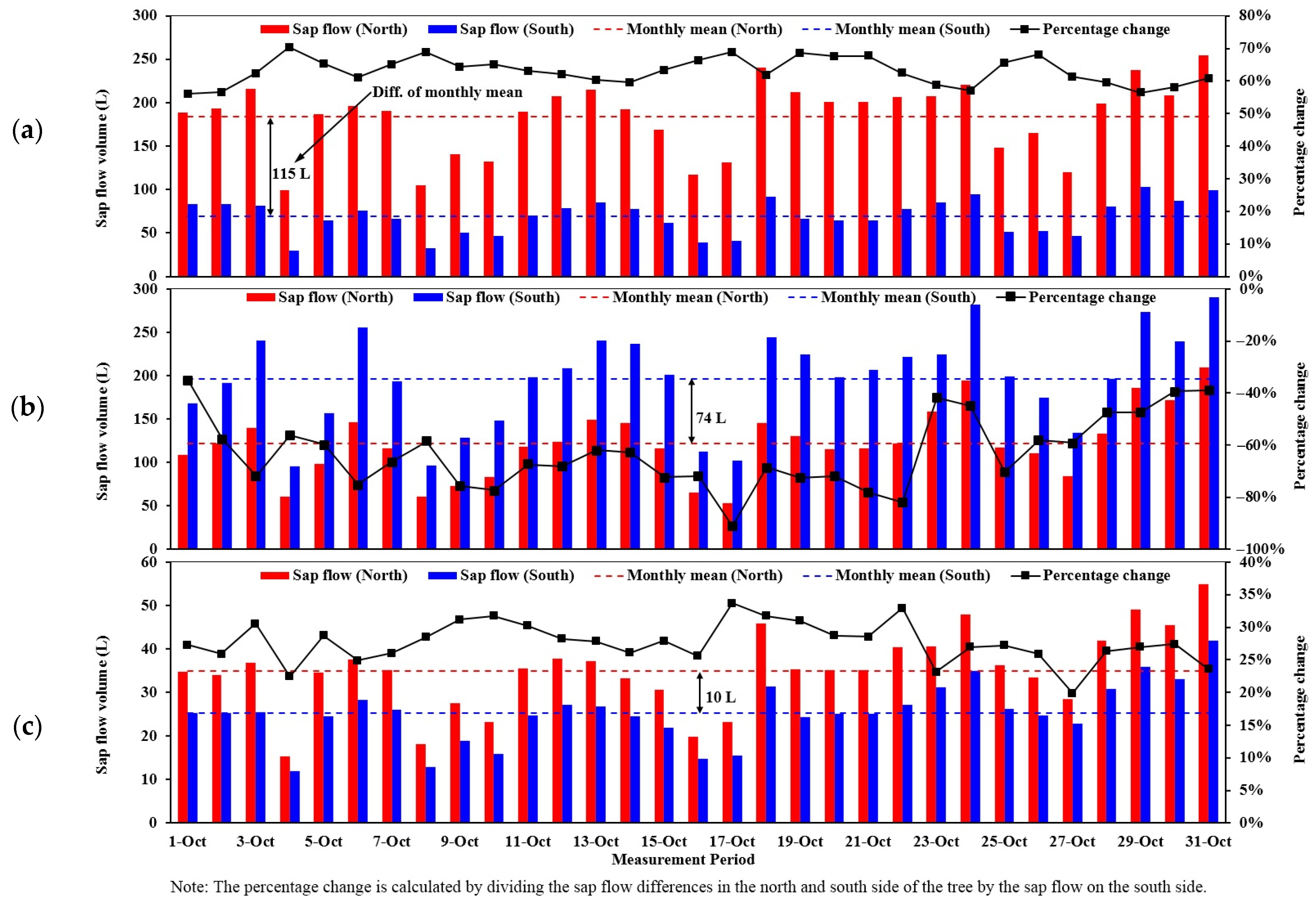
Sap flow in the North and South Sides of the Tree Trunk
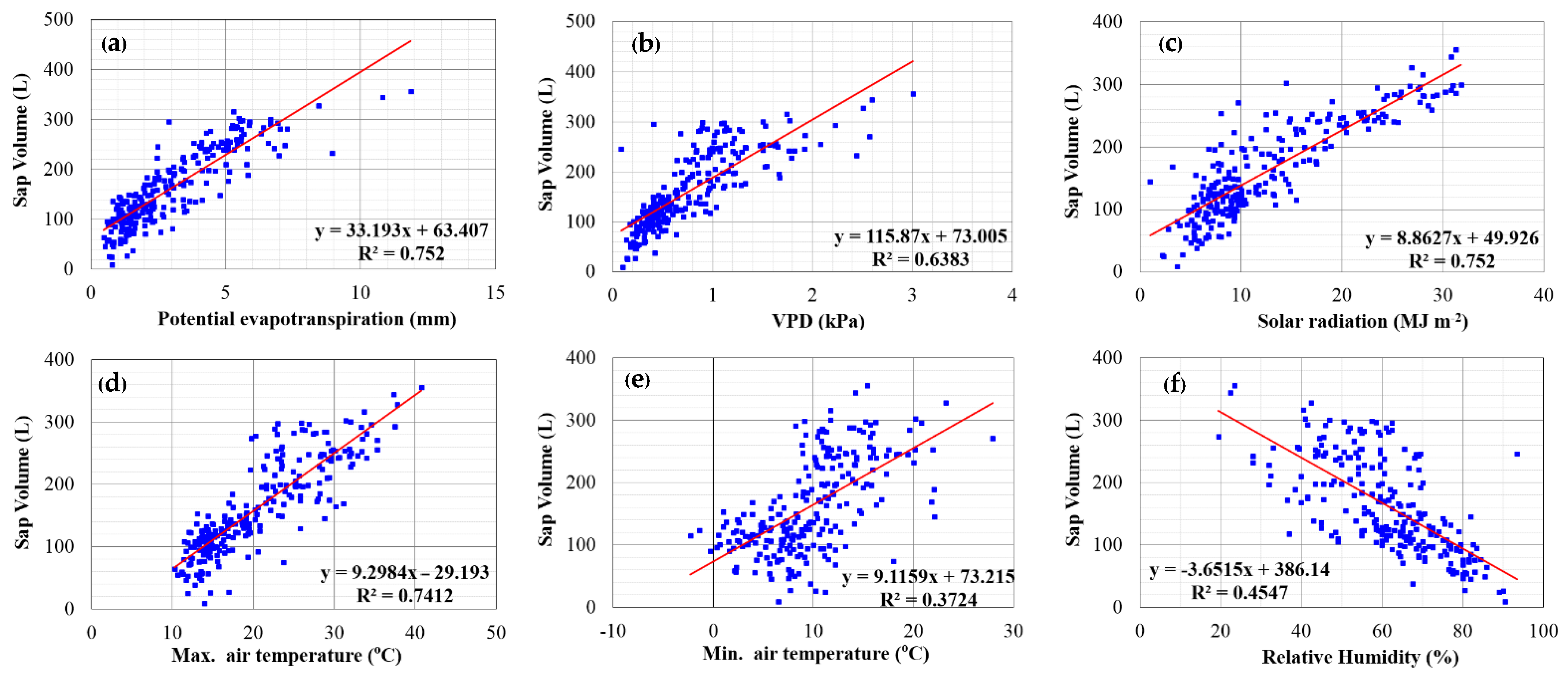
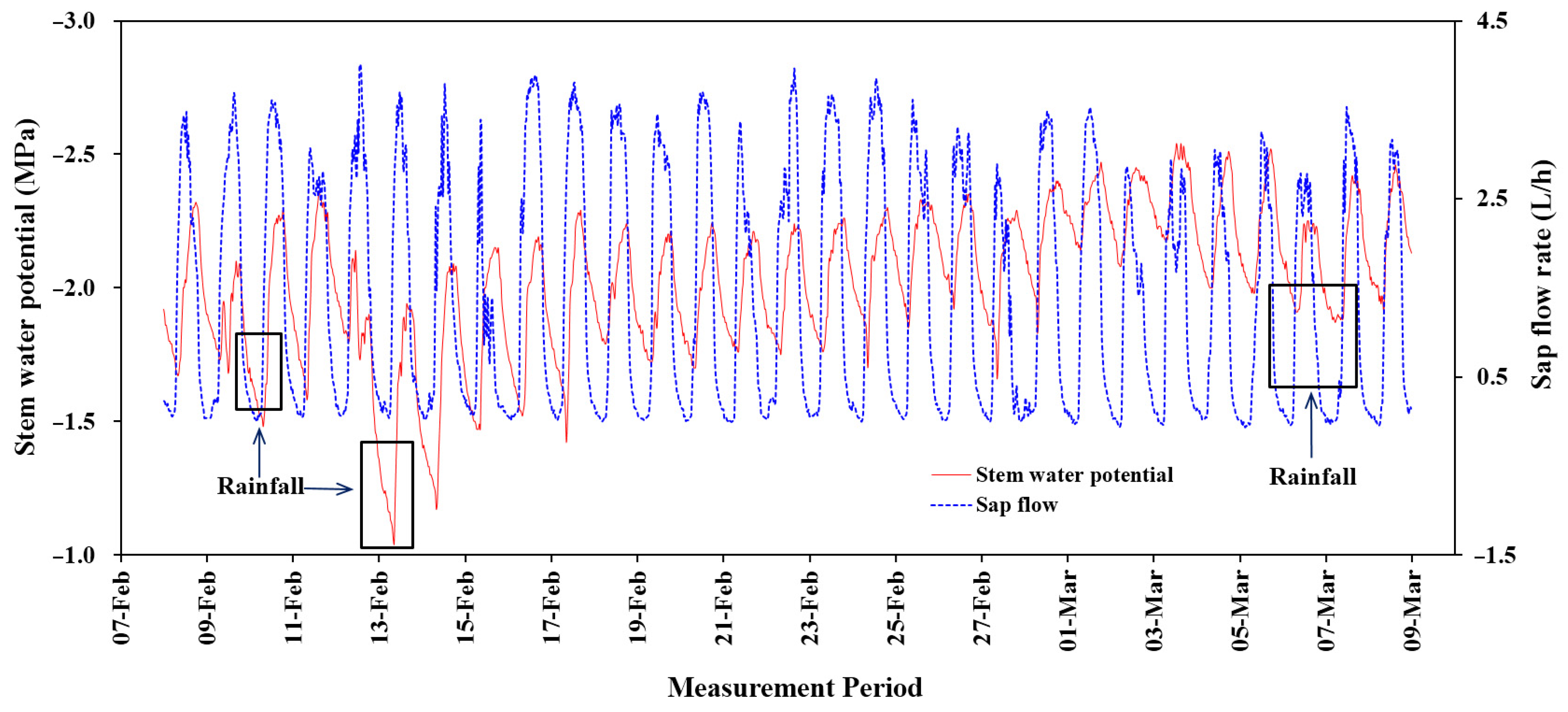
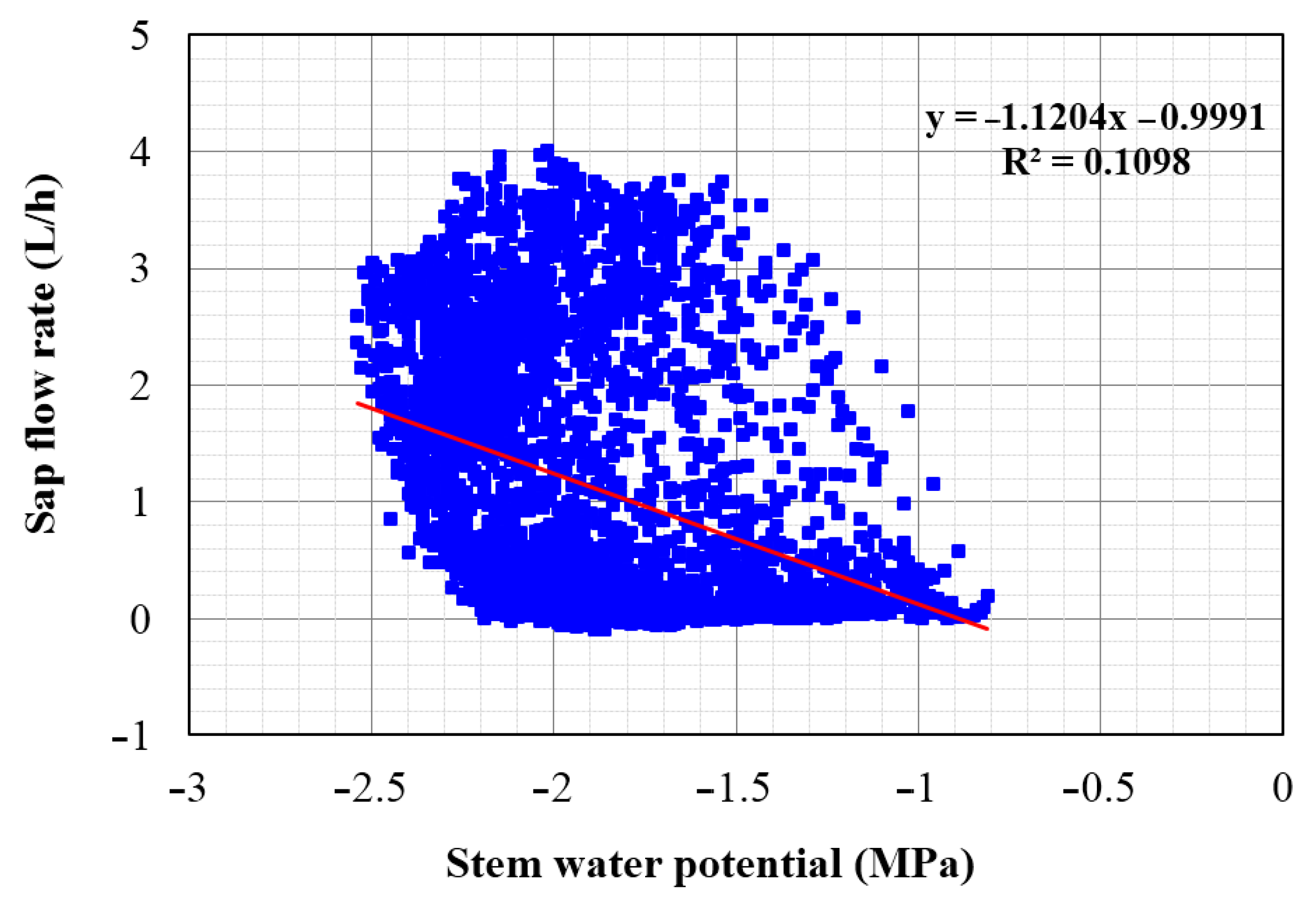
3.6. Relationship between Tree Sap Flow, Meteorological Parameters and Stem Water Potential
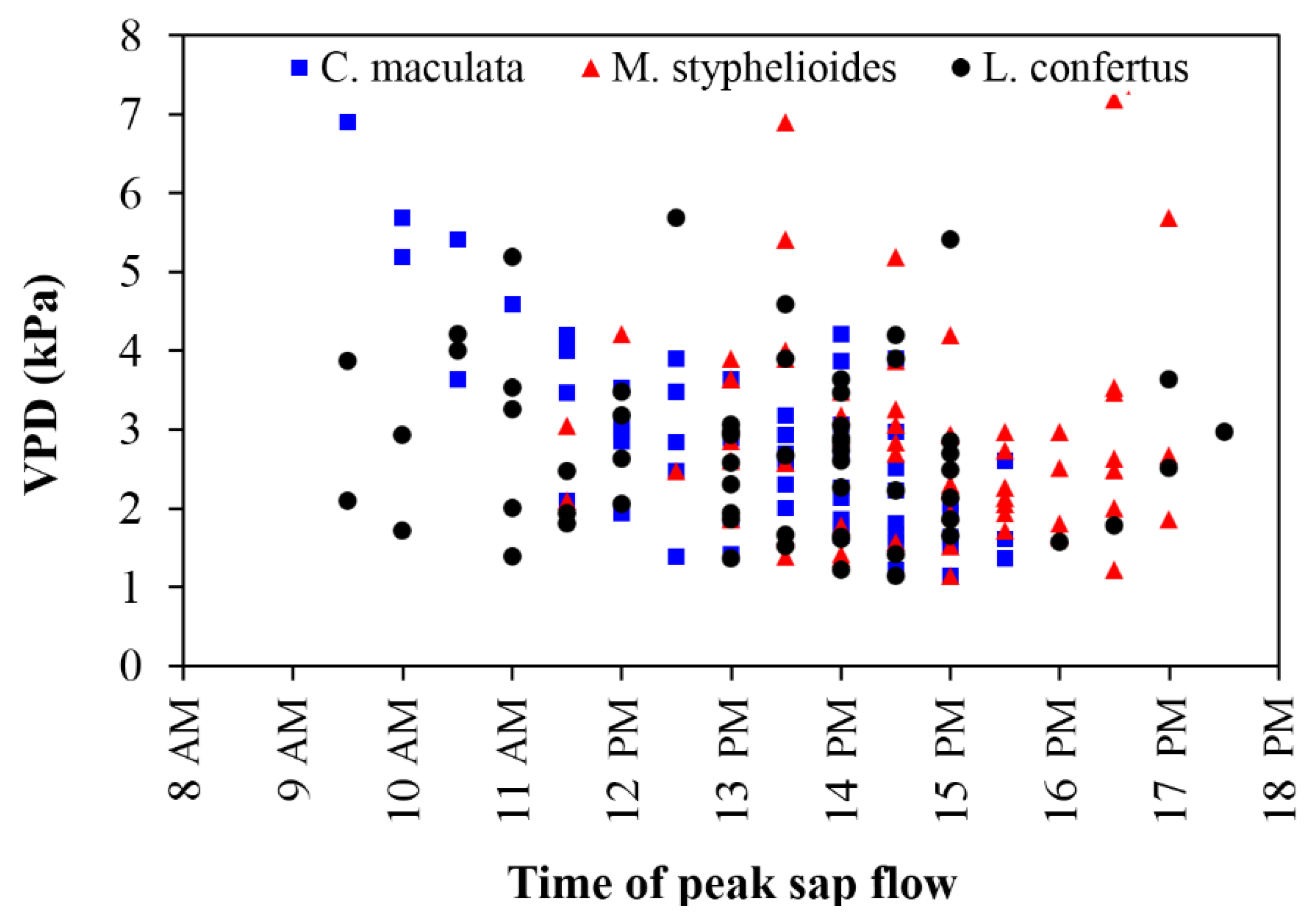
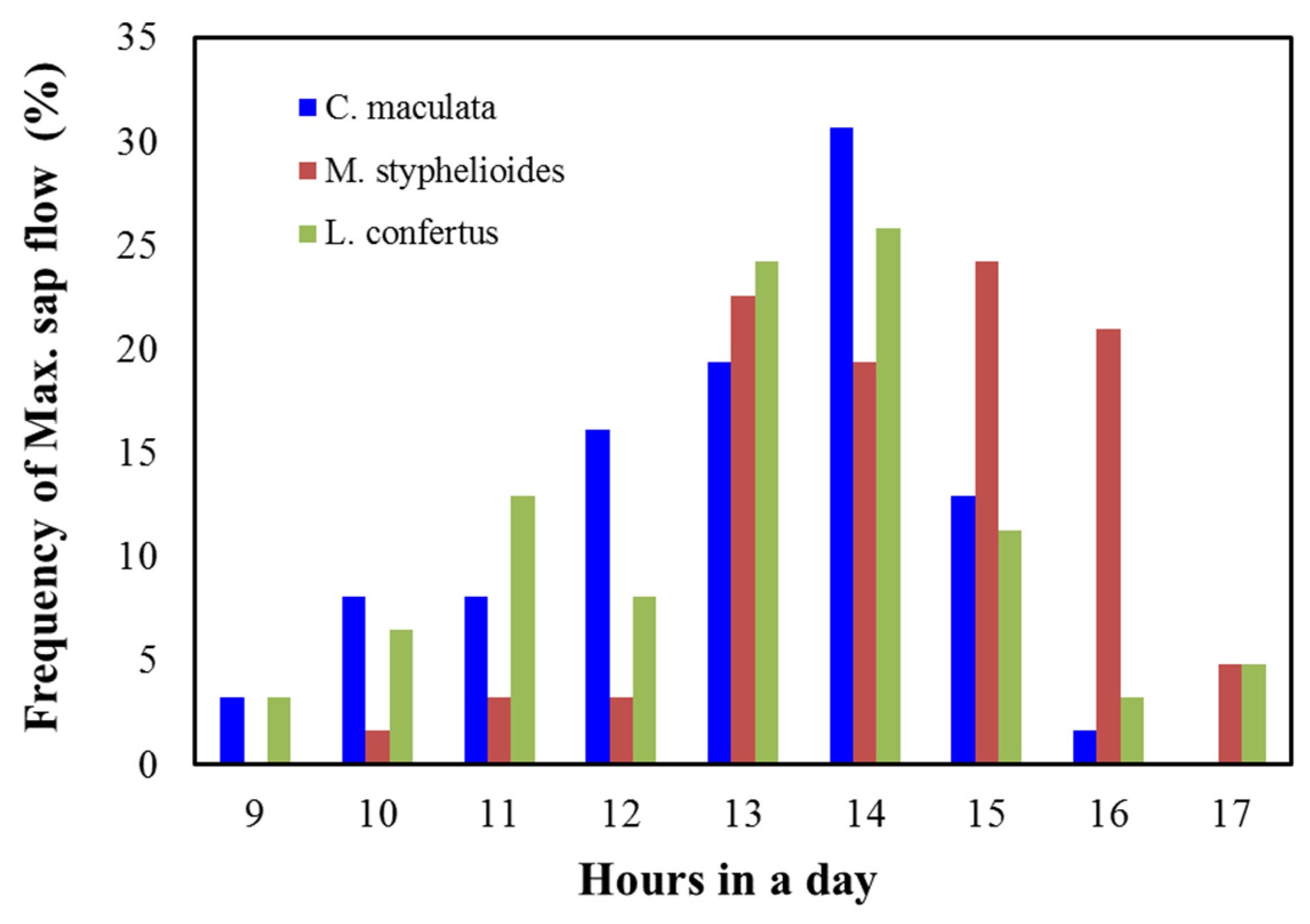
3.7. Tree Performance
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jasechko, S.; Sharp, Z.D.; Gibson, J.J.; Birks, S.J.; Yi, Y.; Fawcett, P.J. Terrestrial water fluxes dominated by transpiration. Nature 2013, 496, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Swanson, R.H. Significant historical developments in thermal methods for measuring sap flow in trees. Agric. For. Meteorol. 1994, 72, 113–132. [Google Scholar] [CrossRef]
- Granier, A.; Biron, P.; Breda, N.; Pontailler, J.Y.; Saugier, B. Transpiration of trees and forest stands: Short and long-term monitoring using sapflow methods. Glob. Chang. Biol. 1996, 2, 265–274. [Google Scholar] [CrossRef]
- Green, S.R.; Clothier, B.E.; McLeod, D.J. The response of sap flow in apple roots to localised irrigation. Agric. Water Manag. 1997, 33, 63–78. [Google Scholar] [CrossRef]
- Li, J.; Zhou, Y.; Guo, L.; Tokhi, H. The establishment of a field site for reactive soil and tree monitoring in Melbourne. J. Aust. Geomech. 2014, 49, 63–72. [Google Scholar]
- Li, J.; Guo, L. Field investigation and numerical analysis of residential building damaged by expansive soil movement caused by tree root drying. ASCE J. Perform. Constr. Facil. 2017, 31, 1–10. [Google Scholar] [CrossRef]
- Li, J. Influence of trees on expansive soils in Melbourne. J. Aust. Geomech. 2018, 53, 61–76. [Google Scholar]
- Burgess, S.S.O.; Adams, M.A.; Turner, N.C.; Beverly, C.R.; Ong, C.K.; Khan, A.A.H.; Bleby, T.M. An improved heat pulse method to measure low and reverse rates of sap flow in woody plants. Tree Physiol. 2001, 21, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Nadezhdina, N.; Steppe, K.; De Pauw, D.J.W.; Bequet, R.; Cermak., J.; Ceulemans, R. Stem-mediated hydraulic redistribution in large roots on opposing sides of a Douglas-fir tree following localised irrigation. New Phytol. 2009, 184, 932–943. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Kang, E.S.; Zhou, M.X. Evaluation of the sap flow using heat pulse method to determine transpiration of the Populus euphratica canopy. Front. For. China 2007, 2, 323–328. [Google Scholar] [CrossRef]
- Fernández, J.E.; Palomo, M.J.; Díaz-Espejo, A.; Clothier, B.E.; Green, S.R.; Girón, I.F.; Moreno, F. Heat-pulse measurements of sap flow in olives for automating irrigation: Tests, root flow and diagnostics of water stress. Agric. Water Manag. 2001, 51, 99–123. [Google Scholar] [CrossRef] [Green Version]
- Juice, S.M.; Templer, P.H.; Phillips, N.G.; Ellison, A.M.; Pelini, S.L. Ecosystem warming increases sap flow rates of northern red oak trees. Ecosphere 2016, 7, 1–17. [Google Scholar] [CrossRef]
- Hatton, T.J.; Wu, H.I. Scaling theory to extrapolate individual tree water use to stand water use. Hydrol. Process. 1995, 9, 527–540. [Google Scholar] [CrossRef]
- Vertessy, R.A.; Hatton, T.J.; Reece, P.; O’Sullivan, S.K.; Benyon, R.G. Estimating stand water use of large mountain ash trees and validation of the sap flow measurement technique. Tree Physiol. 1997, 17, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.S.; Ma, L.Y.; Wang, X.P.; Zhai, M.P. Temporal and spatial variation of sap flow of Chinese pine (Pinus tabulaeformis). J. Beijing For. Univ. 2000, 22, 1–6. [Google Scholar]
- Juhász, Á.; Sepsi, P.; Nagy, Z.; Tőkei, L.; Hrotkó, K. Water consumption of sweet cherry trees estimated by sap flow measurement. Sci. Hortic. 2013, 164, 41–49. [Google Scholar] [CrossRef]
- CSIRO; BOM. Climate Change in Australia; Technical Report; CSIRO Publishing: Canberra, Australia, 2007; p. 148.
- Feng, S.; Fu, Q. Expansion of global drylands under a warming climate. Atmos. Chem. Phys. 2013, 13, 10081–10094. [Google Scholar] [CrossRef] [Green Version]
- Čermák, J.; Nadezhdina, N. Instrumental approaches for studying tree-water relations along gradients of tree size and forest age. In Size- and Age-Related Changes in Tree Structure and Function; Meinzer, F.C., Lachenbruch, B., Dawson, T.E., Eds.; Springer: New York, NY, USA, 2011; pp. 385–426. [Google Scholar]
- Li, J.; Sun, X. Evaluation of changes of Thornthwaite Moisture Index in Victoria. J. Aust. Geomech. 2015, 50, 39–49. [Google Scholar]
- Sun, X.; Li, J.; Zhou, A.N. Assessment of the impact of climate change on expansive soil movements and site classification. J. Aust. Geomech. 2017, 52, 39–50. [Google Scholar]
- Bureau of Meteorology (BOM). Climate Data Online. 2020. Available online: http://www.bom.gov.au/climate/data/ (accessed on 19 October 2020).
- AutoCAD; Autodesk, Inc.: Mill Valley, CA, USA, 2010.
- Coder, K.D. Crown Shape Factors & Volumes; Warnell School of Forestry and Natural Resources, University of Georgia: Athens, GA, USA, 2005. [Google Scholar]
- Marshall, D.C. Measurement of sap flow in conifers by heat transport. Plant Physiol. 1958, 33, 385–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, W.R.N.; Warwick, N.W.M. Transpiration from a kiwifruit vine as estimated by the heat pulse technique and the Penman- Monteith equation. N. Z. J. Agric. Res. 1984, 27, 537–543. [Google Scholar] [CrossRef]
- Hogg, E.H.; Black, T.A.; den Hartog, G.; Neumann, H.H.; Zimmermann, R.; Hurdle, P.A.; Blanken, P.D.; Nesic, Z.; Yang, P.C.; Staebler, R.M.; et al. A comparison of sap flow and eddy fluxes of water vapor from a boreal deciduous forest. J. Geophys. Res. 1997, 102, 28929–28937. [Google Scholar] [CrossRef] [Green Version]
- Swanson, R.H. Numerical and Experimental Analyses of Implanted—Probe Heat Pulse Velocity Theory. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 1983; p. 298. [Google Scholar]
- Monteith, J.L.; Unsworth, M.H. Principles of Environmental Physics; Edward Arnold Publishers: London, UK, 1990. [Google Scholar]
- Allen, R.G.; Walter, I.A.; Elliott, R.L.; Howell, T.A.; Itenfisu, D.; Jensen, M.E.; Snyder, R.L. (Eds.) The ASCE Standardised Reference Evapotranspiration Equation; American Society of Civil Engineers: Reston, VA, USA, 2005; p. 196. [Google Scholar]
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- Mather, J.R. The Climatic Water Balance in Environmental Analysis; D.C. Heath and Company: Lexington, MA, USA, 1978; p. 239. [Google Scholar]
- Mather, J.R. Use of the climatic water budget to estimate streamflow. In Use of the Climatic WATER Budget in Selected Environmental Water Problems; Mather, J.R., Elmer, N.J., Eds.; C.W. Thornthwaite Associates, Laboratory of Climatology, Publications in Climatology, University of Chicago: Chicago, IL, USA, 1979; Volume 32, pp. 1–52. [Google Scholar]
- Sun, X.; Li, J.; Zhou, A.N. Evaluation and comparison of methods for calculating Thornthwaite Moisture Index. J. Aust. Geomech. 2017, 52, 61–75. [Google Scholar]
- Ebrahimi-Birang, N.; Fredlund, D.G. Assessment of the WP4-T Device for Measuring Total Suction. Geotech. Test. J. 2016, 39, 500–506. [Google Scholar] [CrossRef]
- Sun, X.; Li, J.; Cameron, D.A.; Zhou, A.N. Field monitoring and assessment of the impact of a large eucalypt on soil desiccation. Acta Geotech. 2021, 1–14. [Google Scholar] [CrossRef]
- Dixon, M.; Grace, J.; Tyree, M.T. Concurrent measurements of stem density, leaf and stem water potential, stomatal conductance and cavitation on a sapling of Thuja occidentalis L. Plant Cell Environ. 1984, 7, 615–618. [Google Scholar] [CrossRef]
- Edwards, D.R.; Dixon, M. Mechanisms of drought response in Thuja occidentalis L. II. Post-conditioning water stress and stress relief. Tree Physiol. 1995, 15, 129–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, S.; Dixon, M.; Zheng, Y. Vascular blockage in cut roses in a suspension of Pseudomonas fluorescens. J. Hortic. Sci. Biotechnol. 2007, 82, 808–814. [Google Scholar] [CrossRef]
- Vandegehuchte, M.W.; Steppe, K. Sap-flux density measurement methods: Working principles and applicability. Funct. Plant Biol. 2013, 40, 213–223. [Google Scholar] [CrossRef]
- Holman, J.P. Heat Transfer, 9th ed.; McGraw-Hill: New York, NY, USA, 2002; ISBN 978-0-07-029639-8. [Google Scholar]
- Loustau, D.; Berbigier, P.; Roumagnac, P.; Arruda-Pacheco, C.; David, J.S.; Ferreira, M.I.; Pereira, J.S.; Tavares, R. Transpiration of a 64-year-old maritime pine stand in Portugal. 1. Seasonal course of water flux through maritime pine. Oecologia 1996, 107, 33–42. [Google Scholar] [CrossRef] [PubMed]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Tree Height (m) | Crown Height (m) | LCR | DBH (m) | Mean Crown Diameter (m) | Canopy Area (m2) | Crown Volume (m3) |
|---|---|---|---|---|---|---|---|
| C. maculata | 14.6 | 13.0 | 0.9 | 0.56 | 12.8 | 116 | 132.4 |
| M. styphelioides | 8.6 | 7.6 | 0.9 | 0.6 | 7.5 | 40 | 31.2 |
| L. confertus | 6.7 | 5.4 | 0.8 | 0.24 | 5.3 | 20.5 | 16.8 |
| Species | Bark Depth (mm) | Bark Pct. (%) | Sapwood Depth (mm) | Heartwood Depth (mm) | Xylem Radius (mm) | TBA (cm2) | Sapwood Area (cm2) | Sapwood Pct. (%) |
|---|---|---|---|---|---|---|---|---|
| C. maculata | 9.0 | 6.3 | 80.7 | 190.4 | 271.1 | 2465 | 1170 | 47.5 |
| M. styphelioides | 21.5 | 13.7 | 52.8 | 236.6 | 289.4 | 2873 | 873 | 29.4 |
| L. confertus | 5.4 | 8.7 | 36.7 | 78.9 | 115.6 | 460 | 224 | 48.8 |
| Species | wf (g) | wd (g) | V (cm3) | ρb (g/cm3) | mc (%) | Kw (W/mK) | K (W/mK) | C (J/K/kg) | k (mm2/s) |
|---|---|---|---|---|---|---|---|---|---|
| C. maculata | 1.4629 | 0.8121 | 1.28 | 0.63 | 80.14 | 0.821 | 0.708 | 2527 | 0.245 |
| M. styphelioides | 0.9332 | 0.4351 | 0.84 | 0.52 | 114.48 | 0.8291 | 0.692 | 2792 | 0.222 |
| L. confertus | 0.8812 | 0.5223 | 0.79 | 0.67 | 68.72 | 0.7950 | 0.705 | 2415 | 0.260 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Li, J.; Cameron, D.; Moore, G. On the Use of Sap Flow Measurements to Assess the Water Requirements of Three Australian Native Tree Species. Agronomy 2022, 12, 52. https://doi.org/10.3390/agronomy12010052
Sun X, Li J, Cameron D, Moore G. On the Use of Sap Flow Measurements to Assess the Water Requirements of Three Australian Native Tree Species. Agronomy. 2022; 12(1):52. https://doi.org/10.3390/agronomy12010052
Chicago/Turabian StyleSun, Xi, Jie Li, Donald Cameron, and Gregory Moore. 2022. "On the Use of Sap Flow Measurements to Assess the Water Requirements of Three Australian Native Tree Species" Agronomy 12, no. 1: 52. https://doi.org/10.3390/agronomy12010052
APA StyleSun, X., Li, J., Cameron, D., & Moore, G. (2022). On the Use of Sap Flow Measurements to Assess the Water Requirements of Three Australian Native Tree Species. Agronomy, 12(1), 52. https://doi.org/10.3390/agronomy12010052