


Lack of Tradeoff between Leaf Hydraulic Efficiency and Safety across Six Contrasting Water-Stress Tolerant Fruit Tree Species


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description
2.2. Biological Material
2.3. Experimental Setup
2.4. Measurements and Estimates
2.4.1. Leaf Hydraulic Conductance
2.4.2. Pressure-Volume Curves (PVC) Traits
2.4.3. Predawn (Ψpd), Xylem Midday (Ψxyl) Water Potential, and Stomatal Conductance (gs)
2.4.4. Leaf Mass per Area (LMA) and Petiole Density (Dp)
2.4.5. Leaf 13C Isotopic Composition (δ13C)
2.5. Statistical Analysis
Leaf Hydraulic Conductance Vulnerability Curves
3. Results
3.1. Water Status of the Species
3.2. Morpho-Physiological Traits Related to Water Relations
3.3. Pressure–Volume Curve Traits Curve
3.4. Leaf Hydraulic Conductance Vulnerability
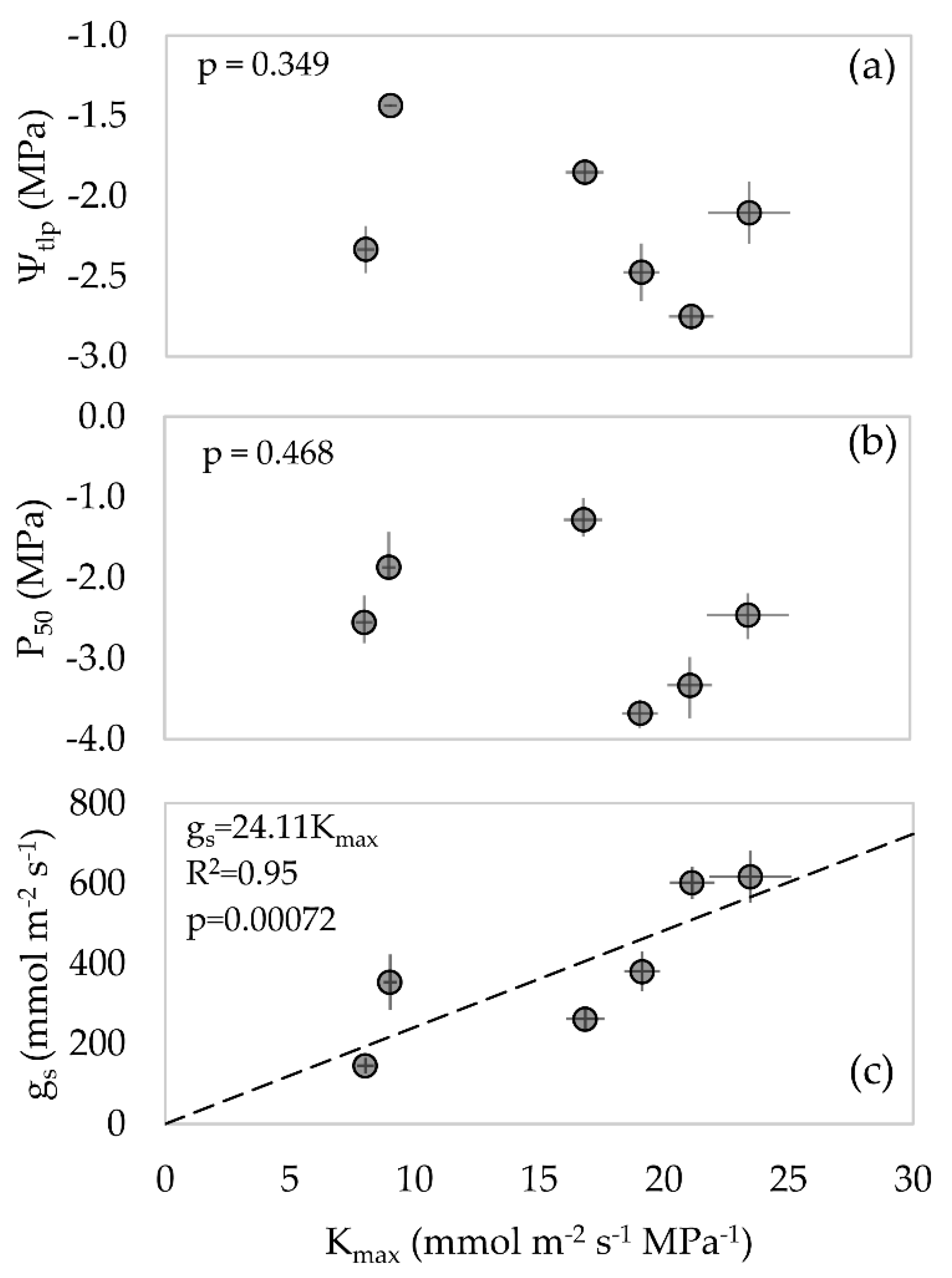
3.5. Relationship between Hydraulic, Water Stress Tolerance, and Morphological Traits
3.6. Correlation Analysis
4. Discussion
4.1. No Tradeoff between Efficiency and Hydraulic Safety across Six Fruit Tree Species
4.2. Ranking of Tolerance to Water Stress and Association between Tolerance Traits and Foliar Morphology
4.3. Leaf Hydraulic Function Ranges
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | P12 (MPa) | Boot-2.5% | Boot–97.5% | P88 (MPa) | Boot–2.5% | Boot–97.5% |
|---|---|---|---|---|---|---|
| Avocado | 0.68 | 0.5 | 1.08 | 1.94 | 1.32 | 2.34 |
| Fig_tree | 1.4 | 0.94 | 1.86 | 3.57 | 3 | 4.71 |
| Mandarin | 1.65 | 1.26 | 2.59 | 3.39 | 2.7 | 3.85 |
| Olive | 1.77 | 1.32 | 2.33 | 5.06 | 3.41 | NA |
| Pomegranate | 1.99 | 1.71 | 2.26 | 5.51 | 5.03 | NA |
| Vine | 1.51 | 1.12 | 1.83 | 2.16 | 1.93 | NA |
Appendix B
| Species | ε (MPa−1) | TLC (mol m−2 MPa−1) | TLC* (mol m−2 MPa−1) |
|---|---|---|---|
| Avocado | 19.77 ± 3.38 a | 0.67 ± 0.13 b | 1.07 ± 0.14 b |
| Fig tree | 10.19 ± 0.84 b | 1.41 ± 0.20 a | 2.43 ± 0.60 a |
| Mandarin | 12.67 ± 2.21 b | 1.17 ± 0.25 a | 2.73 ± 0.52 a |
| Olive | 11.80 ± 2.46 b | 1.22 ± 0.16 a | 2.90 ± 0.74 a |
| Pomegranate | 10.12 ± 1.74 b | 0.98 ± 0.15 a | 1.85 ± 0.53 a |
| Vine | 15.75 ± 1.51 a | 0.49 ± 0.04 b | 0.70 ± 0.08 b |
References
- Verbist, K.; Santibáñez, F.; Gabriels, B.; Soto, G. Atlas de Zonas Áridas de América Latina y el Caribe, 25th ed.; UNESCO-PHI, CALAZAC: Montevideo, Uruguay, 2010; 48p. [Google Scholar]
- Apgaua, D.M.G.; Ishida, F.Y.; Tng, D.Y.P.; Laidlaw, M.J.; Santos, R.M.; Rumman, R.; Eamus, D.; Holtum, J.A.M.; Laurance, S.G.W. Functional traits and water transport strategies in lowland tropical rainforest trees. PLoS ONE 2015, 10, e0130799. [Google Scholar] [CrossRef] [PubMed]
- Tyree, M.T.; Ewers, F.W. The hydraulic architecture of trees and other woody plants. New Phytol. 1991, 119, 345–360. [Google Scholar] [CrossRef]
- Cruiziat, P.; Cochard, H.; Ameglio, T. Hydraulic architecture of trees: Main concepts and results. Ann. For. Sci. 2002, 59, 723–752. [Google Scholar] [CrossRef]
- Cosme, L.H.M.; Schietti, J.; Costa, F.R.C.; Oliveira, R. The importance of hydraulic architecture to the distribution patterns of trees in a central Amazonian forest. New Phytol. 2017, 215, 113–125. [Google Scholar] [CrossRef]
- Brodribb, T.J. Xylem hydraulic physiology: The functional backbone of terrestrial plant productivity. Plant Sci. 2009, 177, 245–251. [Google Scholar] [CrossRef]
- Levitt, J. Responses of Plants to Environmental Stresses: Water, Radiation, Salt, and Other Stresses, 2nd ed.; Academic Press: New York, NY, USA, 1980; 607p. [Google Scholar]
- Ennajeh, M.; Vadel, A.; Cochard, H.; Khemira, H. Comparative impacts of water stress on the leaf anatomy of a drought-resistant and a drought-sensitive olive cultivar. J. Hortic. Sci. Biotechnol. 2010, 85, 289–294. [Google Scholar] [CrossRef]
- Fan, D.-Y.; Jie, S.-L.; Liu, C.-C.; Zhang, X.-Y.; Xu, X.-W.; Zhang, S.-R.; Xie, Z.-Q. The trade-off between safety and efficiency in hydraulic architecture in 31 woody species in a karst area. Tree Physiol. 2011, 31, 865–877. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; Feild, T.S.; Jordan, G. Leaf maximum photosynthetic rate and venation are linked by hydraulics. Plant Physiol. 2007, 144, 1890–1898. [Google Scholar] [CrossRef]
- Sack, L.; Holbrook, N.M. Leaf hydraulics. Annu. Rev. Plant Biol. 2006, 57, 361–381. [Google Scholar] [CrossRef]
- Maire, V.; Wright, I.; Prentice, I.C.; Batjes, N.H.; Bhaskar, R.; van Bodegom, P.; Cornwell, W.; Ellsworth, D.; Niinemets, Ü.; Ordonez, A.; et al. Global effects of soil and climate on leaf photosynthetic traits and rates. Glob. Ecol. Biogeogr. 2015, 24, 706–717. [Google Scholar] [CrossRef]
- McCulloh, K.A.; Domec, J.; Johnson, D.M.; Smith, D.D.; Meinzer, F.C. A dynamic yet vulnerable pipeline: Integration and coordination of hydraulic traits across whole plants. Plant Cell Environ. 2019, 42, 2789–2807. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.M.; McCulloh, K.A.; Meinzer, F.; Woodruff, D.R.; Eissenstat, D.M. Hydraulic patterns and safety margins, from stem to stomata, in three eastern US tree species. Tree Physiol. 2011, 31, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.M.; Berry, Z.C.; Baker, K.V.; Smith, D.D.; McCulloh, K.A.; Domec, J. Leaf hydraulic parameters are more plastic in species that experience a wider range of leaf water potentials. Funct. Ecol. 2018, 32, 894–903. [Google Scholar] [CrossRef]
- Sack, L.; Buckley, T.N.; Scoffoni, C. Why are leaves hydraulically vulnerable? J. Exp. Bot. 2016, 67, 4917–4919. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Vilalta, J.; Poyatos, R.; Aguadé, D.; Retana, J.; Mencuccini, M. A new look at water transport regulation in plants. New Phytol. 2014, 204, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; Holbrook, N.M. Diurnal depression of leaf hydraulic conductance in a tropical tree species. Plant Cell Environ. 2004, 27, 820–827. [Google Scholar] [CrossRef]
- Domec, J.-C.; Gartner, B.L. Cavitation and water storage capacity in bole xylem segments of mature and young Douglas-fir trees. Trees Struct. Funct. 2001, 15, 204–214. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Klein, T.; Jansen, S.; Choat, B.; Sack, L. The correlations and sequence of plant stomatal, hydraulic, and wilting responses to drought. Proc. Natl. Acad. Sci. USA 2016, 113, 13098–13103. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Wheeler, J.K.; Castro, L. Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiol. 2006, 26, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Meinzer, F.C.; McCulloh, K.A.; Lachenbruch, B.; Woodruff, D.R.; Johnson, D.M. The blind men and the elephant: The impact of context and scale in evaluating conflicts between plant hydraulic safety and efficiency. Oecologia 2010, 164, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Maherali, H.; Pockman, W.; Jackson, R.B. Adaptive variation in the vulnerability of woody plants to xylem cavitation. Ecology 2004, 85, 2184–2199. [Google Scholar] [CrossRef]
- Tyree, M.T.; Davis, S.D.; Cochard, H. Biophysical perspectives of xylem evolution: Is there a trade-off of hydraulic efficiency for vulnerability to dysfunction? IAWA J. 1994, 15, 335–360. [Google Scholar] [CrossRef]
- Garrido, M.; Bown, H.; Ayamante, J.; Orell, M.; Sánchez, A.; Acevedo, E. The adjustment of Prosopis tamarugo hydraulic architecture traits has a homeostatic effect over its performance under descent of phreatic level in the Atacama Desert. Trees Struct. Funct. 2020, 34, 89–99. [Google Scholar] [CrossRef]
- Gleason, S.M.; Westoby, M.; Jansen, S.; Choat, B.; Hacke, U.G.; Pratt, R.B.; Bhaskar, R.; Brodribb, T.J.; Bucci, S.J.; Cao, K.F.; et al. Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species. New Phytol. 2016, 209, 123–136. [Google Scholar] [CrossRef]
- Ocheltree, T.W.; Nippert, J.B.; Prasad, P.V.V. A safety vs efficiency trade-off identified in the hydraulic pathway of grass leaves is decoupled from photosynthesis, stomatal conductance and precipitation. New Phytol. 2016, 210, 97–107. [Google Scholar] [CrossRef]
- De Guzman, M.E.; Santiago, L.S.; Schnitzer, S.A.; Álvarez-Cansino, L. Trade-offs between water transport capacity and drought resistance in neotropical canopy liana and tree species. Tree Physiol. 2016, 37, 1404–1414. [Google Scholar] [CrossRef]
- Dayer, S.; Herrera, J.; Dai, Z.; Burlett, R.; Lamarque, L.J.; Delzon, S.; Bortolami, G.; Cochard, H.; A Gambetta, G. The sequence and thresholds of leaf hydraulic traits underlying grapevine varietal differences in drought tolerance. J. Exp. Bot. 2020, 71, 4333–4344. [Google Scholar] [CrossRef]
- Torres-Ruiz, J.M.; Diaz-Espejo, A.; Perez-Martin, A.; Hernandez-Santana, V. Role of hydraulic and chemical signals in leaves, stems and roots in the stomatal behaviour of olive trees under water stress and recovery conditions. Tree Physiol. 2015, 35, 415–424. [Google Scholar] [CrossRef]
- Atauri, I.D.G.C. Adaptation du Modèle Stics à la Vigne (Vitis vinifera L.): Utilisation Dans le Cadre D’Une éTude D’Impact du Changement Climatique à L’éChelle de la France. Ph.D. Thesis, Montpellier SupAgro, Montpellier, France, 2006. [Google Scholar]
- Goharrizi, K.J.; Hamblin, M.R.; Karami, S.; Nazari, M. Physiological, biochemical, and metabolic responses of abiotic plant stress: Salinity and drought. Turk. J. Bot. 2021, 45 (Suppl. S1), 623–642. [Google Scholar] [CrossRef]
- Giordano, M.; Petropoulos, S.A.; Rouphael, Y. Response and defence mechanisms of vegetable crops against drought, heat and salinity stress. Agriculture 2021, 11, 463. [Google Scholar] [CrossRef]
- Hassani, A.; Azapagic, A.; Shokri, N. Predicting long-term dynamics of soil salinity and sodicity on a global scale. Proc. Natl. Acad. Sci. USA 2020, 117, 33017–33027. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant Salinity Stress: Many Unanswered Questions Remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef]
- Luebert, F.; Pliscoff, P. Sinopsis Bioclimática y Vegetacional de Chile; Editorial Universitaria: Santiago, Chile, 2006. [Google Scholar]
- Brodribb, T.J.; Holbrook, N.M. Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiol. 2003, 132, 2166–2173. [Google Scholar] [CrossRef]
- Graciano, C.; Fernández, M.E. Curvas Presión-Volumen Para Estimación de Parámetros Hídricos. In Técnicas de Medición en Ecofisiología Vegetal: Conceptos y Procedimientos; Fernández, M., Gyenge, J., Eds.; INTA: Buenos Aires, Argentina, 2010; Chapter 6; pp. 79–86. 140p. [Google Scholar]
- Scholander, P.F.; Bradstreet, E.D.; Hemmingsen, E.A.; Hammel, H.T. Sap pressure in vascular plants: Negative hydrostatic pressure can be measured in plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Bchir, A.; Escalona, J.M.; Gallé, A.; Hernández-Montes, E.; Tortosa, I.; Braham, M.; Medrano, H. Carbon isotope discrimination (δ13C) as an indicator of vine water status and water use efficiency (WUE): Looking for the most representative sample and sampling time. Agric. Water Manag. 2016, 167 (Suppl. C), 11–20. [Google Scholar] [CrossRef]
- Ogle, K.; Barber, J.J.; Willson, C.; Thompson, B. Hierarchical statistical modeling of xylem vulnerability to cavitation. New Phytol. 2009, 182, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Duursma, R.; Choat, B. Fitplc—An R package to fit hydraulic vulnerability curves. J. Plant Hydraul. 2017, 4, 2. [Google Scholar] [CrossRef]
- Liu, H.; Ye, Q.; Gleason, S.M.; He, P.; Yin, D. Weak trade-off between xylem hydraulic efficiency and safety: Climatic seasonality matters. New Phytol. 2020, 229, 1440–1452. [Google Scholar] [CrossRef]
- Whitehead, D.; Edwards WR, N.; Jarvis, P.G. Conducting sapwood area, foliage area, and permeability in mature trees of Picea sitchensis and Pinus contorta. Can. J. For. Res. 1984, 14, 940–947. [Google Scholar] [CrossRef]
- Westoby, M.; Wright, I.J. Land-plant ecology on the basis of functional traits. Trends Ecol. Evol. 2006, 21, 261–268. [Google Scholar] [CrossRef]
- Degu, A.; Hochberg, U.; Wong, D.C.J.; Alberti, G.; Lazarovitch, N.; Peterlunger, E.; Castellarin, S.D.; Herrera, J.C.; Fait, A. Swift metabolite changes and leaf shedding are milestones in the acclimation process of grapevine under prolonged water stress. BMC Plant Biol. 2019, 19, 69. [Google Scholar] [CrossRef] [PubMed]
- Scoffoni, C.; Albuquerque, C.; Cochard, H.; Buckley, T.N.; Fletcher, L.R.; Caringella, M.A.; Bartlett, M.; Brodersen, C.R.; Jansen, S.; McElrone, A.J.; et al. The causes of leaf hydraulic vulnerability and its influence on gas exchange in Arabidopsis thaliana. Plant Physiol. 2018, 178, 1584–1601. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, S.; Vico, G.; Katul, G.; Palmroth, S.; Jackson, R.B.; Porporato, A. Hydraulic limits on maximum plant transpiration and the emergence of the safety-efficiency trade-off. New Phytol. 2013, 198, 169–178. [Google Scholar] [CrossRef]
- Rodriguez-Iturbe, I.; Porporato, A. Ecohydrology of Water-Controlled Ecosystems; Cambridge University Press: Cambridge, UK, 2004. [Google Scholar]
- Urli, M.; Porté, A.J.; Cochard, H.; Guengant, Y.; Burlett, R.; Delzon, S. Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees. Tree Physiol. 2013, 33, 672–683. [Google Scholar] [CrossRef]
- Sack, L.; Cowan, P.D.; Jaikumar, N.; Holbrook, N.M. The ‘hydrology’ of leaves: Co-ordination of structure and function in temperate woody species. Plant Cell Environ. 2003, 26, 1343–1356. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Woodruff, D.R.; Marias, D.E.; Smith, D.D.; McCulloh, K.A.; Howard, A.R.; Magedman, A.L. Mapping ‘hydroscapes’ along the iso-to anisohydric continuum of stomatal regulation of plant water status. Ecol. Lett. 2016, 19, 1343–1352. [Google Scholar] [CrossRef]
- Huang, L.; Li, Z.; Liu, Q.; Pu, G.; Zhang, Y.; Li, J. Research on the adaptive mechanism of photosynthetic apparatus under salt stress: New directions to increase crop yield in saline soils. Ann. Appl. Biol. 2019, 175, 1–17. [Google Scholar] [CrossRef]
- Perri, S.; Katul, G.G.; Molini, A. Xylem–phloem hydraulic coupling explains multiple osmoregulatory responses to salt stress. New Phytol. 2019, 224, 644–662. [Google Scholar] [CrossRef]
- Pérez-Pérez, J.; Robles, J.; Tovar, J.; Botía, P. Response to drought and salt stress of lemon ‘Fino 49’ under field conditions: Water relations, osmotic adjustment and gas exchange. Sci. Hortic. 2009, 122, 83–90. [Google Scholar] [CrossRef]
- Poorter, L.; Wright, S.J.; Paz, H.; Ackerly, D.; Condit, R.; Ibarra-Manríquez, G.; Harms, K.E.; Licona, J.C.; Martínez-Ramos, M.; Mazer, S.J.; et al. Are functional traits good predictors of demographic rates? Evidence from five neotropical forests. Ecology 2008, 89, 1908–1920. [Google Scholar] [CrossRef]
- Opazo, I.; Toro, G.; Salvatierra, A.; Pastenes, C.; Pimentel, P. Rootstocks modulate the physiology and growth responses to water deficit and long-term recovery in grafted stone fruit trees. Agric. Water Manag. 2020, 228, 105897. [Google Scholar] [CrossRef]
- Fu, X.; Meinzer, F.C. Metrics and proxies for stringency of regulation of plant water status (iso/anisohydry): A global data set reveals coordination and trade-offs among water transport traits. Tree Physiol. 2019, 39, 122–134. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Herrera, J.C.; Dayer, S.; Feng, Q.; Hochberg, U.; Castellarin, S.D. The physiology of drought stress in grapevine: Towards an integrative definition of drought tolerance. J. Exp. Bot. 2020, 71, 4658–4676. [Google Scholar] [CrossRef] [PubMed]
- Friedlingstein, P.; Joel, G.; Field, C.B.; Fung, I.Y. Toward an allocation scheme for global terrestrial carbon models. Glob. Chang. Biol. 1999, 5, 755–770. [Google Scholar] [CrossRef]
- Silva, S.; Miranda, M.T.; Costa, V.E.; Machado, E.C.; Ribeiro, R.V. Sink strength of citrus rootstocks under water deficit. Tree Physiol. 2021, 41, 1372–1383. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Moutinho-Pereira, J.M.; Gonçalves, B.C.; Ferreira, H.F.; Correia, C.M. Changes in growth, gas exchange, xylem hydraulic properties and water use efficiency of three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2007, 60, 183–192. [Google Scholar] [CrossRef]
- Hochberg, U.; Bonel, A.G.; David-Schwartz, R.; Degu, A.; Fait, A.; Cochard, H.; Peterlunger, E.; Herrera, J.C. Grapevine acclimation to water deficit: The adjustment of stomatal and hydraulic conductance differs from petiole embolism vulnerability. Planta 2017, 245, 1091–1104. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Holbrook, N.M.; Edwards, E.J.; Gutiérrez, M.V. Relations between stomatal closure, leaf turgor and xylem vulnerability in eight tropical dry forest trees. Plant Cell Environ. 2003, 26, 443–450. [Google Scholar] [CrossRef]
- Flexas, J.; Carriquí, M.; Nadal, M. Gas exchange and hydraulics during drought in crops: Who drives whom? J. Exp. Bot. 2018, 69, 3791–3795. [Google Scholar] [CrossRef] [Green Version]




| Scientific Name | Common Name | Cultivar |
|---|---|---|
| Citrus reticulata Blanco | Mandarin | Orogrande (grafted over ‘Carrizo’ (Citrus sinensis L. Osb.× Poncirus trifoliata L. Raf.)) |
| Ficus carica L. | Fig tree | Black Mission |
| Olea europaea L. | Olive | Sevillana |
| Persea americana Mill. | Avocado | Hass (grafted over ‘Mexícola’ (Persea americana Mill.)) |
| Punica granatum L. | Pomegranate | Wonderfull |
| Vitis vinifera L. | Vine | Emperor |
| Species | gs (mmol m2 s−1) | Kmax (mmol MPa−1 m2 s−1) | Ψpd (MPa) | Ψxyl (MPa) |
|---|---|---|---|---|
| Avocado | 262.0 ± 48.7 b | 16.9 ± 0.77 b | −0.48 ± 0.02 a | −1.08 ± 0.10 a |
| Fig tree | 616.7 ± 61.6 a | 23.5 ± 1.65 a | −0.54 ± 0.09 a | −1.28 ± 0.12 a |
| Mandarin | 145.3 ± 51.1 c | 8.0 ± 0.34 c | −0.59 ± 0.06 a | −1.52 ± 0.09 b |
| Olive | 601.9 ± 45.9 a | 21.1 ± 0.90 a | −0.61 ± 0.05 a | −1.61 ± 0.15 b |
| Pomegranat | 380.6 ± 52.1 b | 19.1 ± 0.72 b | −0.57 ± 0.04 a | −1.31 ± 0.10 a |
| Vine | 353.6 ± 52.1 b | 9.0 ± 0.26 c | −0.53 ± 0.07 a | −1.04 ± 0.10 a |
| Species | LMA (g m−2) | Dp (mg mm−3) | δ13C (‰) |
|---|---|---|---|
| Avocado | 108.5 ± 4.1 b | 0.32 ± 0.01 b | −27.5 ± 0.6 b |
| Fig tree | 86.7 ± 5.6 c | 0.21 ± 0.02 a | −27.8 ± 0.6 b |
| Mandarin | 124.3 ± 10.5 b | 0.44 ± 0.02 c | −25.1 ± 0.6 a |
| Olive | 181.0 ± 14.4 a | 0.53 ± 0.05 c | −25.6 ± 0.3 a |
| Pomegranate | 95.8 ± 3.0 c | 0.35 ± 0.06 b | −25.3 ± 0.3 a |
| Vine | 52.9 ± 4.1 d | 0.14 ± 0.01 a | −26.5 ± 0.8 b |
| Species | Ψo (MPa) | Ψtlp (MPa) |
|---|---|---|
| Avocado | −1.53 ± 0.09 b | −1.85 ± 0.07 b |
| Fig tree | −1.64 ± 0.19 b | −2.10 ± 0.19 b |
| Mandarin | −1.66 ± 0.12 b | −2.33 ± 0.15 c |
| Olive | −1.86 ± 0.23 b | −2.75 ± 0.08 c |
| Pomegranate | −1.81 ± 0.22 b | −2.48 ± 0.18 c |
| Vine | −1.13 ± 0.03 a | −1.44 ± 0.01 a |
| Species | P50 | Boot–2.5% | Boot–97.5% | Sx | Boot–2.5% | Boot–97.5% |
|---|---|---|---|---|---|---|
| (MPa) | (MPa−1) | |||||
| Avocado | −1.28 a | 1.07 | 1.55 | 72.44 a | 54.13 | 210.34 |
| Fig tree | −2.46 b | 2.16 | 2.73 | 42.26 a | 25.33 | 74.63 |
| Mandarin | −2.55 b | 2.29 | 2.88 | 53.18 a | 38.01 | 844.06 |
| Olive | −3.33 c | 2.92 | 3.68 | 27.84 a | 18.48 | 84.29 |
| Pomegranate | −3.68 c | 3.50 | 3.85 | 26.00 a | 21.39 | 31.66 |
| Vine | −1.87 b | 1.74 | 2.31 | 145.75 a | 43.37 | 947.41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido, M.I.; Vergara, S. Lack of Tradeoff between Leaf Hydraulic Efficiency and Safety across Six Contrasting Water-Stress Tolerant Fruit Tree Species. Agronomy 2022, 12, 2351. https://doi.org/10.3390/agronomy12102351
Garrido MI, Vergara S. Lack of Tradeoff between Leaf Hydraulic Efficiency and Safety across Six Contrasting Water-Stress Tolerant Fruit Tree Species. Agronomy. 2022; 12(10):2351. https://doi.org/10.3390/agronomy12102351
Chicago/Turabian StyleGarrido, Marco Isaac, and Sebastián Vergara. 2022. "Lack of Tradeoff between Leaf Hydraulic Efficiency and Safety across Six Contrasting Water-Stress Tolerant Fruit Tree Species" Agronomy 12, no. 10: 2351. https://doi.org/10.3390/agronomy12102351
APA StyleGarrido, M. I., & Vergara, S. (2022). Lack of Tradeoff between Leaf Hydraulic Efficiency and Safety across Six Contrasting Water-Stress Tolerant Fruit Tree Species. Agronomy, 12(10), 2351. https://doi.org/10.3390/agronomy12102351