


Use of Phosphonate Salts to Control Chestnut ‘Brown Rot’ by Gnomoniopsis castaneae in Fruit Orchards of Castanea sativa

 ,
,  , ,
, , 

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Evaluated Products
2.2. In Vitro Tests
2.2.1. Inhibition of Mycelial Growth
2.2.2. Inhibition of Conidia Germination
2.3. Treatment Trials
2.4. Samples Processing
2.5. DNA Extractions and qPCR Reactions
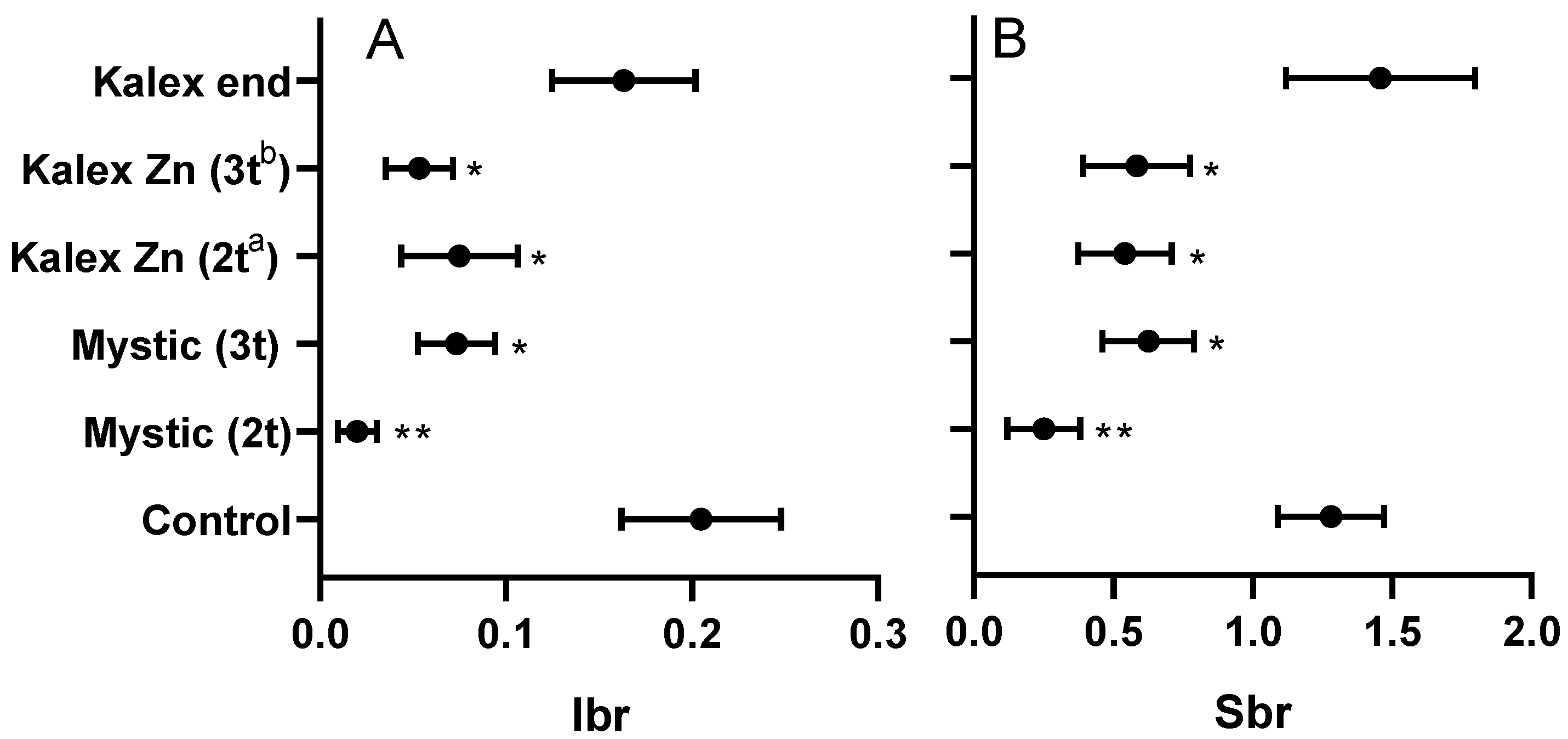
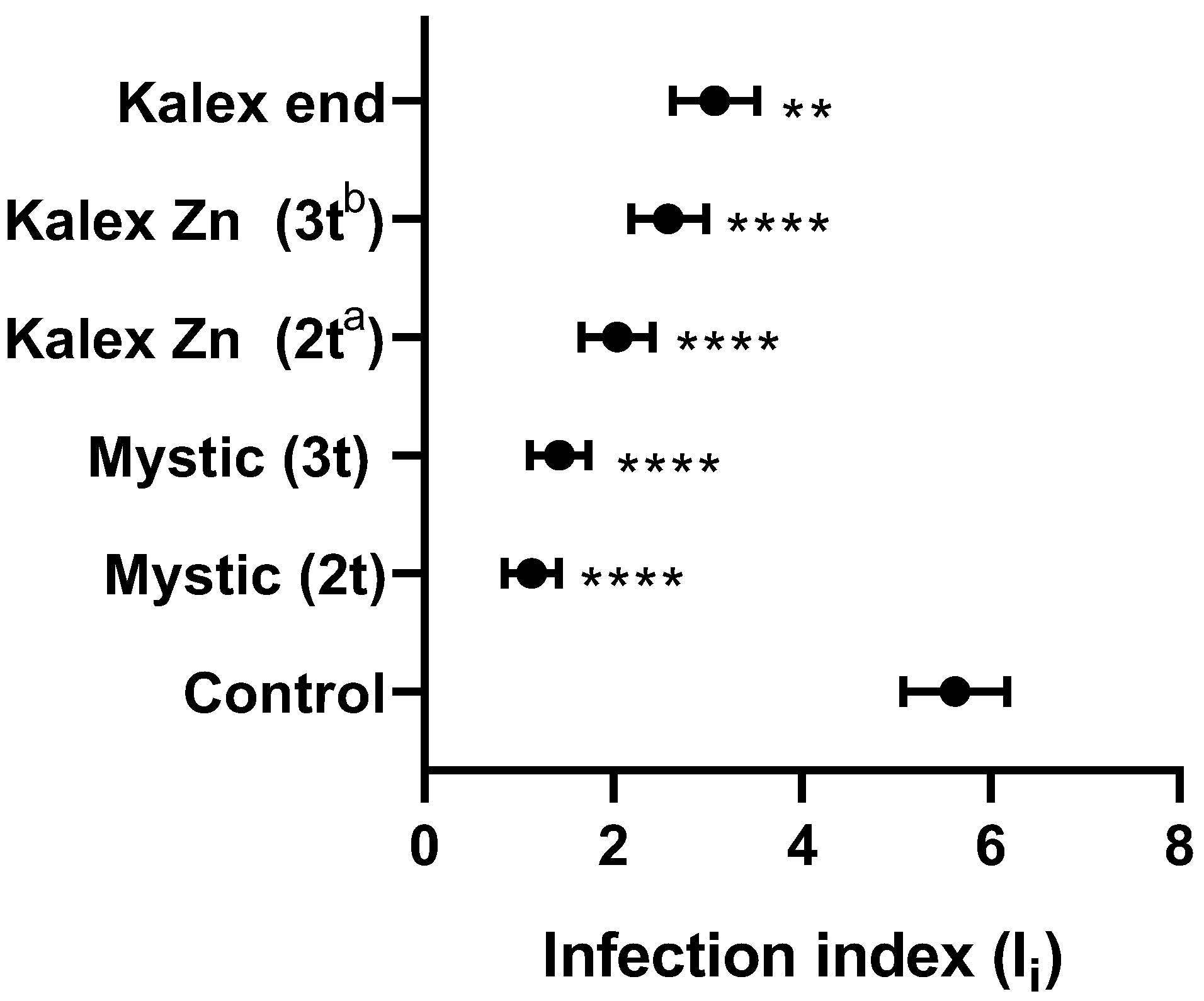
2.6. Assessment of Disease and Infection Indexes
2.6.1. Disease Indexes Based on Symptoms Expression
2.6.2. Infection Index Based on qPCR Results
2.7. Residues in Fruits
2.8. Statistical Analyses
3. Results
3.1. In Vitro Test
3.2. Treatment Trials
3.3. Residues in Fruits
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Crous, P.; Summerell, B.; Shivas, R.; Burgess, T.; Decock, C.; Dreyer, L.; Granke, L.; Guest, D.; Hardy, G.S.; Hausbeck, M. Fungal planet description sheets: 107–127. Pers.-Mol. Phylogeny Evol. Fungi 2012, 28, 138–182. [Google Scholar] [CrossRef]
- Visentin, I.; Gentile, S.; Valentino, D.; Gonthier, P.; Tamietti, G.; Cardinale, F. Gnomoniopsis castanea sp. nov. (Gnomoniaceae, Diaporthales) as the causal agent of nut rot in sweet chestnut. J. Plant Pathol. 2012, 94, 411–419. [Google Scholar] [CrossRef]
- Tamietti, G. On the fungal species Gnomoniopsis castaneae (“castanea”) and its synonym G. smithogilvyi. J. Plant Pathol. 2016, 1, 189–190. [Google Scholar] [CrossRef]
- Maresi, G.; Longa, C.O.; Turchetti, T. Brown rot on nuts of Castanea sativa Mill: An emerging disease and its causal agent. iForest-Biogeosci. For. 2013, 6, 294–301. [Google Scholar] [CrossRef] [Green Version]
- Shuttleworth, L.A.; Liew, E.C.Y.; Guest, D.I. Survey of the incidence of chestnut rot in south-eastern Australia. Australas. Plant Pathol. 2012, 42, 63–72. [Google Scholar] [CrossRef]
- Morales-Rodríguez, C.; Bastianelli, G.; Caccia, R.; Bedini, G.; Massantini, R.; Moscetti, R.; Thomidis, T.; Vannini, A. Impact of ‘brown rot’caused by Gnomoniopsis castanea on chestnut fruits during the post-harvest process: Critical phases and proposed solutions. J. Sci. Food Agric. 2022, 102, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Shuttleworth, L.A.; Guest, D.I. The infection process of chestnut rot, an important disease caused by Gnomoniopsis smithogilvyi (Gnomoniaceae, Diaporthales) in Oceania and Europe. Australas. Plant Pathol. 2017, 46, 397–405. [Google Scholar] [CrossRef]
- Lione, G.; Giordano, L.; Sillo, F.; Gonthier, P. Testing and modelling the effects of climate on the incidence of the emergent nut rot agent of chestnut Gnomoniopsis castanea. Plant Pathol. 2014, 64, 852–863. [Google Scholar] [CrossRef]
- Fernández Fernández, M.M.; Bezos García, D.; Díez Casero, J.J. Fungi associated with necrotic galls of Dryocosmus kuriphilus (hymenoptera: Cynipidae) in northern spain. Silva Fenn. 2018, 52, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Vannini, A.; Vettraino, A.; Martignoni, D.; Morales-Rodríguez, C.; Contarini, M.; Caccia, R.; Paparatti, B.; Speranza, S. Does Gnomoniopsis castanea contribute to the natural biological control of chestnut gall wasp? Fungal Biol. 2017, 121, 44–52. [Google Scholar] [CrossRef]
- Vannini, A.; Morales-Rodríguez, C. Integrated disease management in tree nut cultivation. In Achieving Sustainable Cultivation of Tree Nuts; Serdar, Ü., Fulbright, D., Eds.; Burleigh Dodds Science Pub: Cambridge, UK, 2019; p. 530. [Google Scholar]
- Dennert, F.G.; Broggini, G.A.L.; Gessler, C.; Storari, M. Gnomoniopsis castanea is the main agent of chestnut nut rot in Switzerland. Phytopathol. Mediterr. 2015, 54, 199–211. [Google Scholar] [CrossRef]
- Sakalidis, M.L.; Medina-Mora, C.; Kolp, M.; Fulbright, D.W. First Report of Gnomoniopsis smithogilvyi Causing Chestnut Brown Rot on Chestnut Fruit in Michigan. Plant Dis. 2019, 103, 2134. [Google Scholar] [CrossRef]
- Cisterna-Oyarce, V.; Carrasco-Fernández, J.; Castro, J.F.; Santelices, C.; Muñoz-Reyes, V.; Millas, P.; Buddie, A.G.; France, A. Gnomoniopsis smithogilvyi: Identification, characterization and incidence of the main pathogen causing brown rot in postharvest sweet chestnut fruits (Castanea sativa) in Chile. Australas. Plant Dis. Notes 2022, 17, 1–3. [Google Scholar] [CrossRef]
- Silva-Campos, M.; Islam, T.; Cahill, D.M. Fungicide control of Gnomoniopsis smithogilvyi, causal agent of chestnut rot in Australia. Australas. Plant Pathol. 2022, 51, 483–494. [Google Scholar] [CrossRef]
- Morales-Rodríguez, C.; Vettraino, A.M.; Vannini, A. Efficacy of Biofumigation with Brassica carinata Commercial Pellets (BioFence) to Control Vegetative and Reproductive Structures of Phytophthora cinnamomi. Plant Dis. 2016, 100, 324–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vannini, A.; Morales-Rodriguez, C.; Aleandri, M.; Bruni, N.; Valle, M.D.; Mazzetto, T.; Martignoni, D.; Vettraino, A. Emerging new crown symptoms on Castanea sativa (Mill.): Attempting to model interactions among pests and fungal pathogens. Fungal Biol. 2018, 122, 911–917. [Google Scholar] [CrossRef]
- Turco, S.; Bastianelli, G.; Morales-Rodríguez, C.; Vannini, A.; Mazzaglia, A. Development of a taqman qpcr assay for the detection and quantification of Gnomoniopsis castaneae in chestnut tissues. For. Pathol. 2021, 51, 1–10. [Google Scholar] [CrossRef]
- Vannini, A.; Natili, G.; Anselmi, N.; Montaghi, A.; Vettraino, A.M. Distribution and gradient analysis of Ink disease in chestnut forests. For. Pathol. 2010, 40, 73–86. [Google Scholar] [CrossRef]
- Dann, E.; McLeod, A. Phosphonic acid: A long-standing and versatile crop protectant. Pest Manag. Sci. 2020, 77, 2197–2208. [Google Scholar] [CrossRef]
- Guest, D.I.; Bompeix, G. The Complex Mode of Action of Phosphonates. Australas. Plant Pathol. 1990, 19, 113–115. [Google Scholar] [CrossRef]
- Brunings, A.M.; Datnoff, L.E.; Simonne, E.H. Phosphorous Acid and Phosphoric Acid: When all P Sources are not Equal. EDIS 2005, 4, 1–8. [Google Scholar] [CrossRef]
- Sutradhar, A.K.; Arnall, D.B.; Dunn, B.L.; Raun, W.R. Does phosphite, a reduced form of phosphate contribute to phosphorus nutrition in corn (Zea mays L.)? J. Plant Nutr. 2019, 42, 982–989. [Google Scholar] [CrossRef]
- Manghi, M.C.; Masiol, M.; Calzavara, R.; Graziano, P.L.; Peruzzi, E.; Pavoni, B. The use of phosphonates in agriculture. Chemical, biological properties and legislative issues. Chemosphere 2021, 283, 131187. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, P.; Kannan, H.; Das, S.; Young, M.; Santra, S. Comparative analysis of copper and zinc based agrichemical biocide products: Materials characteristics, phytotoxicity and in vitro antimicrobial efficacy. AIMS Environ. Sci. 2016, 3, 439–455. [Google Scholar] [CrossRef]
- He, L.; Liu, Y.; Mustapha, A.; Lin, M. Antifungal activity of zinc oxide nanoparticles against Botrytis cinerea and Penicillium expansum. Microbiol. Res. 2011, 166, 207–215. [Google Scholar] [CrossRef]
- Król, A.; Pomastowski, P.; Rafińska, K.; Railean-Plugaru, V.; Buszewski, B. Zinc oxide nanoparticles: Synthesis, antiseptic activity and toxicity mechanism. Adv. Colloid Interface Sci. 2017, 249, 37–52. [Google Scholar] [PubMed]
- Ashajyothi, C.; Prabhurajeshwar, C.; Handral, H.K. Investigation of antifungal and anti-mycelium activities using biogenic nanoparticles: An eco-friendly approach. Environ. Nanotechnol. Monit. Manag. 2016, 5, 81–87. [Google Scholar]
- Whiley, A.; Hargreaves, P.; Pegg, K.; Doogan, V.; Ruddle, L.; Saranah, J.; Langdon, P. Changing sink strengths influence translocation of phosphonate in avocado (Persea americana Mill.) trees. Aust. J. Agric. Res. 1995, 46, 1079–1090. [Google Scholar] [CrossRef]
- Tkaczyk, M.; Pacia, A.; Siebyła, M.; Oszako, T. Phosphite fertilisers as inhibitors of Hymenoscyphus fraxineus (anamorph Chalara fraxinea) growth in tests in vitro. Folia For. Pol. 2017, 59, 79–81. [Google Scholar] [CrossRef]
- Felipini, R.B.; Boneti, J.I.; Katsurayama, Y.; Neto, A.C.R.; Veleirinho, B.; Maraschin, M.; Di Piero, R.M. Apple scab control and activation of plant defence responses using potassium phosphite and chitosan. Eur. J. Plant Pathol. 2016, 145, 929–939. [Google Scholar] [CrossRef]
- Kromann, P.; Pérez, W.G.; Taipe, A.; Schulte-Geldermann, E.; Sharma, B.P.; Andrade-Piedra, J.; Forbes, G.A. Use of Phosphonate to Manage Foliar Potato Late Blight in Developing Countries. Plant Dis. 2012, 96, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Gessler, C.; Pertot, I.; Perazzolli, M. Plasmopara viticola: A review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediterr. 2011, 50, 3–44. [Google Scholar]
- Lione, G.; Giordano, L.; Sillo, F.; Brescia, F.; Gonthier, P. Temporal and spatial propagule deposition patterns of the emerging fungal pathogen of chestnut Gnomoniopsis castaneae in orchards of north-western Italy. Plant Pathol. 2021, 70, 2016–2033. [Google Scholar] [CrossRef]
- Einhardt, A.M.; Souza, G.M.F.; Silveira, P.R.; Rodrigues, F. Potassium, calcium, and zinc phosphites on white mold control in soybean. Bragantia 2020, 79, 417–424. [Google Scholar] [CrossRef]
- Gómez-Merino, F.C.; Trejo-Téllez, L.I. Conventional and novel uses of phosphite in horticulture: Potentialities and challenges. Italus Hortus 2016, 23, 1–13. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Product | Application | Rate | Date | Phenological Stage |
|---|---|---|---|---|---|
| 2019 | Kalex® | endotherapy | 0.8 mL/tree | 13 June | After bud burst |
| 2019 | Kalex Zn® | crown spray | 3 L/ha | 25 June and 6 July | Blooming and burr formation |
| 2019 | Mystic® 430 SC | crown spray | 350 mL/ha | 25 June and 6 July | Blooming and burr formation |
| 2019 | Control | - | - | - | - |
| 2020 | Kalex® | endotherapy | 0.8 mL/tree | 23 June | Blooming |
| 2020 | Kalex Zn® | crown spray | 3 L/ha | 23 June and 9 July | Blooming |
| 2020 | Kalex Zn® | crown spray | 3 L/ha | 23 June; 9 July and 27 August | Blooming; burr development and kernel development |
| 2020 | Mystic® 430 SC | crown spray | 350 mL/ha | 23 June and 9 July | Blooming |
| 2020 | Mystic® 430 SC | crown spray | 350 mL/ha | 23 June; 9 July and 27 August | Blooming; burr development and kernel development |
| 2020 | Control | - | - | - | - |
| EC50 (µL L−1) | ||||
|---|---|---|---|---|
| GN01 | GNAm | |||
| Mycelium | Conidia | Mycelium | Conidia | |
| Mystic® 430 SC | 0.06 | 0.017 | 0.06 | 0.01 |
| Kalex Zn® | 8 × 102 | 2 × 103 | 1 × 103 | 1.8 × 103 |
| Kalex® | 2.8 × 103 | 1 × 104 | 2.4 × 103 | 1 × 104 |
| Phosphonic Acid and Its Salts (mg/Kg ± SEM) | |
|---|---|
| Kalex Zn® | 7.05 ± 0.2 |
| Kalex® | 19 ± 4.2 |
| Untreated control | 0.19 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bastianelli, G.; Morales-Rodríguez, C.; Caccia, R.; Turco, S.; Rossini, L.; Mazzaglia, A.; Thomidis, T.; Vannini, A. Use of Phosphonate Salts to Control Chestnut ‘Brown Rot’ by Gnomoniopsis castaneae in Fruit Orchards of Castanea sativa. Agronomy 2022, 12, 2434. https://doi.org/10.3390/agronomy12102434
Bastianelli G, Morales-Rodríguez C, Caccia R, Turco S, Rossini L, Mazzaglia A, Thomidis T, Vannini A. Use of Phosphonate Salts to Control Chestnut ‘Brown Rot’ by Gnomoniopsis castaneae in Fruit Orchards of Castanea sativa. Agronomy. 2022; 12(10):2434. https://doi.org/10.3390/agronomy12102434
Chicago/Turabian StyleBastianelli, Giorgia, Carmen Morales-Rodríguez, Romina Caccia, Silvia Turco, Luca Rossini, Angelo Mazzaglia, Thomas Thomidis, and Andrea Vannini. 2022. "Use of Phosphonate Salts to Control Chestnut ‘Brown Rot’ by Gnomoniopsis castaneae in Fruit Orchards of Castanea sativa" Agronomy 12, no. 10: 2434. https://doi.org/10.3390/agronomy12102434
APA StyleBastianelli, G., Morales-Rodríguez, C., Caccia, R., Turco, S., Rossini, L., Mazzaglia, A., Thomidis, T., & Vannini, A. (2022). Use of Phosphonate Salts to Control Chestnut ‘Brown Rot’ by Gnomoniopsis castaneae in Fruit Orchards of Castanea sativa. Agronomy, 12(10), 2434. https://doi.org/10.3390/agronomy12102434